Détermination des effets de la tique d'hiver (Dermacentor albipictus) sur la santé des orignaux : une approche expérimentale - Mémoire Delphine De ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Détermination des effets de la tique d'hiver
(Dermacentor albipictus) sur la santé des orignaux : une
approche expérimentale
Mémoire
Delphine De Pierre
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Delphine De Pierre, 2023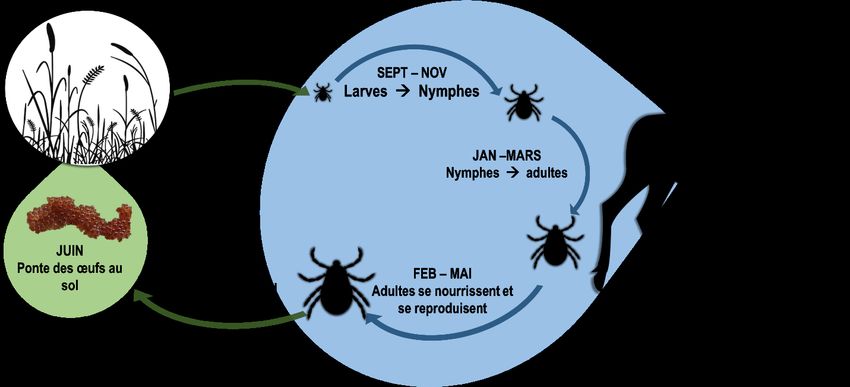
Détermination des effets de la tique d’hiver
(Dermacentor albipictus) sur la santé des orignaux : une
approche expérimentale
Mémoire
Delphine De Pierre
Sous la direction de :
Jean-Pierre Tremblay, directeur de recherche
Patrick Leighton, codirecteur de recherche
Résumé
Malgré que les effets des tiques d’hiver (Dermacentor abipictus) sur l’orignal (Alces alces) ont été étudiés en
captivité, peu d’études, à l’échelle individuelle, se sont penchées sur la relation entre la tique et l’orignal en
milieu naturel. En effet, bien que l’orignal soit un cervidé généralement bien adapté aux conditions hivernales,
ce n’est pas tout à fait le cas des veaux orignaux qui vivent leur premier hiver. Ainsi, les réponses
physiologiques des veaux orignaux faisant face à cette épreuve s’ajoutent à celles enclenchées par une
infestation par la tique d’hiver. Afin d’éclaircir les conséquences de la tique d’hiver sur la santé des veaux
orignaux, il importe de départager l’effet du parasitisme par la tique de l’effet des conditions hivernales. Pour
ce faire, nous avons testé l’efficacité d’un acaricide en application topique à base de perméthrine visant à
réduire la charge de tiques d’hiver chez les veaux orignaux traités. Nous avons également effectué un suivi
de ces mêmes veaux orignaux qui évoluaient dans deux régions du Nouveau-Brunswick le long d’un gradient
latitudinal en utilisant un plan d’échantillonnage par capture-recapture au cours d’un hiver. L’objectif étant
d’étudier les effets des tiques d’hiver et de la sévérité de l’hiver, dans deux régions distinctes, sur la masse
et les paramètres sanguins en tant qu’indices de la condition corporelle des veaux orignaux. À la suite de
cette étude, nos résultats nous indiquent que le traitement acaricide n’a pas réussi à réduire suffisamment
la charge de tiques afin d’influencer la condition corporelle des animaux traités. Malgré ce constat, les
résultats des analyses sanguines révèlent tout de même des différences entre les individus des deux régions
dont certaines pourraient s’expliquer en partie par la présence des tiques d’hiver. Cependant, ces résultats
doivent être considérés avec prudence étant donné le nombre réduit d’individus et l’absence de données en
lien avec les ressources alimentaires qui influencent aussi la condition corporelle des animaux.
iiAbstract
Even though the effects of winter ticks (Dermacentor abipictus) on moose (Alces alces) have been studied
in captivity, few studies, at the individual scale, have examined the relationship between ticks and moose in
the wild. Although moose are generally well adapted to winter conditions, this is not entirely true for moose
calves in their first winter. Thus, the physiological responses of moose calves facing their first winter are
additional to those triggered by winter tick infestation. In order to clarify the consequences of winter ticks on
the health of moose calves, it is necessary to separate the effect of tick parasitism from the effect of winter
conditions. To do this, we tested the efficacy of a permethrin based topical acaricide in reducing winter tick
burden in treated moose calves. We also followed these same moose calves as they moved along a
latitudinal gradient in two regions of New Brunswick using a capture-recapture sampling design over the
course of a winter. The objective was to investigate the effects of winter ticks and winter severity, in two
distinct regions, on mass and blood parameters as indices of body condition of moose calves. As a result of
this study, our results indicate that the acaricide treatment was not successful in reducing the tick load
sufficiently to influence the body condition of treated animals. Despite this setback, the blood test results still
show differences between individuals in the two regions, some of which may be explained in part by the
presence of winter ticks. However, these results must be considered with caution given the small number of
individuals and the absence of data related to food resources that influence the body condition of the animals
as well.
iiiTable des matières
Résumé .................................................................................................................................................................. ii
Abstract ................................................................................................................................................................. iii
Table des matières................................................................................................................................................. iv
Liste tableaux......................................................................................................................................................... vi
Liste des figures.................................................................................................................................................... vii
Remerciements...................................................................................................................................................... ix
Avant-propos ........................................................................................................................................................ xii
Introduction générale...............................................................................................................................................1
Impacts comportementaux des parasites sur leurs hôtes......................................................................................3
Impact des parasites sur la santé et la condition corporelle de leurs hôtes ............................................................3
Contrôle des tiques .............................................................................................................................................5
Impact des conditions hivernales sur la condition corporelle .................................................................................6
Modèles de l’étude ..............................................................................................................................................8
Objectifs ........................................................................................................................................................... 10
Approche méthodologique................................................................................................................................. 11
Chapitre 1. Evaluation of a topical acaricide to reduce winter tick loads in wild moose ............................................. 12
Résumé ............................................................................................................................................................ 13
Abstract ............................................................................................................................................................ 14
Introduction....................................................................................................................................................... 15
Methods ........................................................................................................................................................... 16
Study area.................................................................................................................................................... 16
Moose Capture and Acaricide Treatment ....................................................................................................... 16
Statistical Analyses ....................................................................................................................................... 17
Results ............................................................................................................................................................. 18
Discussion ........................................................................................................................................................ 20
Conclusion and Recommendations ................................................................................................................... 21
Acknowledgements ........................................................................................................................................... 22
Chapitre 2. Variation in body condition of moose calves facing contrasting winter severity and tick loads ................. 24
Résumé ............................................................................................................................................................ 25
Abstract ............................................................................................................................................................ 26
Introduction....................................................................................................................................................... 27
ivMethods ........................................................................................................................................................... 29
Study area.................................................................................................................................................... 29
Capture and handling.................................................................................................................................... 30
Body condition estimation ............................................................................................................................. 31
Statistical analyses ....................................................................................................................................... 31
Results ............................................................................................................................................................. 34
Discussion ........................................................................................................................................................ 38
Conclusion........................................................................................................................................................ 41
Acknowledgements ........................................................................................................................................... 41
Conclusion générale ............................................................................................................................................. 43
Efficacité de l’acaricide...................................................................................................................................... 43
Effet de la région et des tiques sur la condition corporelle et le profil sanguin...................................................... 45
Perspectives de recherche ................................................................................................................................ 47
Bibliographie ......................................................................................................................................................... 49
vListe tableaux
Tableau 1. Contribution des différents auteurs des deux articles présentés dans ce mémoire ................................ xiii
Table 2. Hematological parameters used to interpret changes in the body condition of moose calves between winter
and spring in two contrasted regions of New Brunswick, Canada. Arrows in the interpretation column indicate the
direction of the change in the parameter values. Reference values were obtained from the Animal Health Diagnosis
Centre of the College of Veterinary Medicine of Cornell University. (Cornell University 2022b)................................. 33
Table 3. General linear mixed models results on body mass and hematological parameters according to the period of
capture, region and their interaction. Transformations for non-normally distributed variables are indicated next to the
parameter name. The last column is the adjusted p-value for multiple tests using the false discovery rate. Bold
adjusted-p values are significant or present significant tendencies. ......................................................................... 37
viListe des figures
Figure 1. Adaptation du triangle épidémiologique à partir de la représentation graphique par Antonovics (2017) .......7
Figure 2. Cycle de vie de la tique d’hiver (Adapté de; Ministère Forêts Faune et Parcs 2016) ...................................9
Figure 3. Temporal change in A) tick load, B) body mass (kg) and C) hematocrit (%) in moose calves treated with 5
% permethrin (3 ml/45 kg ; solid red line) compared to controls (blue, dashed line) in New Brunswick, Canada from
early winter to spring 2018. Dots represent either A) the mean number of ticks per 10 cm transect counted in January
(ntreated = 10, ncontrol = 8) and in April/May (ntreated = 7, ncontrol = 6) or B) the mass (kg) measured in January (ntreated = 10,
ncontrol = 8) and in April/May (ntreated = 6, ncontrol = 5) or C) the hematocrit (%) measured in January (ntreated = 9, ncontrol = 8)
and in April/May (ntreated = 7, ncontrol = 6). Values are given as means with 95 % confidence intervals for treated and
untreated calves for each period based on a A) a generalized linear mixed model (GLMM) with a Poisson distribution
or B) and C) linear mixed models. Dots occurring only in January or in spring refer either to calves that were not
recaptured in spring (winter death: ntreated = 1, ncontrol = 1, GPS failure: ntreated= 1, no capture opportunities: ntreated = 1,
ncontrol = 1) or missing values at recapture: ntreated = 2, ncontrol = 2. .............................................................................. 19
Figure 4. Study area showing the capture locations in January for moose calves in northern (n = 10) and southern (n
= 8) New Brunswick. ............................................................................................................................................. 30
Figure 5. Comparison of tick loads in January for calves (n = 18) from two populations in northern and southern
New-Brunswick. Black lines represent means. ....................................................................................................... 34
Figure 6. Temporal change in body mass and hematological parameters of moose calves from northern (solid blue
line) and southern (orange dashed line) New Brunswick, Canada between early winter and spring 2018. Dots
represent raw values and values for the same individual are connected with a line. We report the parameter
estimates from mixed linear models as means and 95% confidence intervals for moose calves in each region for
each time period. For interpretation purposes we present the estimates from the linear mixed models on the
untransformed variables after verifying that they were comparable with the model on the normal scale. Dots occurring
only in January refer to calves that either died or were not recaptured in spring. Dots occurring only in spring refer to
variables that were not measured during first capture due to logistic errors. ............................................................ 36
vii“I wish there was a way to know you were in the
good old days before you actually left them.” –
Andrew Bernard (Ed Helms)
viiiRemerciements
Je tiens à débuter cette section en spécifiant qu’heureusement, l’intérêt immédiat qui s’est allumé dans mon
cerveau dès qu’on m’a proposé une recherche sur le parasitisme chez l’orignal ne s’est pas éteint au fil des
ans. C’est vrai que ce n’aura pas toujours été facile, mais une chose est certaine, j’ai toujours continué
d’aimer mon sujet de maîtrise. Ça peut sembler bien peu, mais c’est une chance de rester accrochée par un
projet qui nous suit pendant plusieurs années. Dans mon cas, après 5 années de doutes et de stress, mais
aussi de victoires, de persévérance et de découvertes, j’en suis enfin à rédiger un mot afin de remercier les
gens qui m’ont soutenu tout au long de cette aventure.
Je souhaite commencer en remerciant mon directeur de maîtrise Jean-Pierre Tremblay. C’est un homme
passionné par ses projets de recherche avec qui il est toujours plaisant et enrichissant de discuter de science.
Par contre, ce qui m’aura marqué de Jean-Pierre c’est son positivisme, ses valeurs humaines et sa
disponibilité pour ses étudiants. Je me suis généralement fait un devoir de remercier Jean-Pierre tout au long
de mon parcours pour son soutien constant, mais je souhaite vraiment souligner maintenant à quel point son
aide et sa bonne humeur auront été appréciées. Je veux également remercier Patrick Leighton, mon co-
directeur, pour sa contribution qui, bien qu’à distance, a su apporter un point de vue différent et essentiel à
cette recherche. Il m’a chaleureusement accueilli dans son laboratoire à Saint-Hyacinthe et a toujours été
présent lorsque j’avais besoin de son avis. Je ne pourrais pas non plus oublier de remercier spécialement
Steeve Côté qui a été celui qui m’a proposé mon projet de maîtrise. En plus de me présenter cette opportunité
en or, Steeve a été présent dans toutes les étapes de mon parcours à la maîtrise, que ce soit pour
m’accompagner durant les premiers jours sur le terrain, relire mes manuscrits et se montrer compréhensif
dans mes moments de fragilité.
Lors des captures d’orignaux sur le terrain, j’ai eu la chance de côtoyer des gens passionnés qui m’ont
marqué par leur gentillesse et leur générosité. De chez Irving, merci à John Gilbert pour son accueil
chaleureux et ses histoires toujours divertissantes et Charles Neveu qui m’a accompagné et épaulé tout au
long des captures et même après. Merci Charles, d’avoir pris soin de cette petite étudiante un peu beaucoup
stressée par ses premières captures d’orignaux. Ton aide et ton enthousiasme ont été fort appréciés. Merci
aussi à Mike Bannerman le pilote d’hélicoptère de chez Irving. Merci à Beth Matheson et Mike Reed qui, en
plus d’avoir été extrêmement efficaces, m’ont donné de précieux conseils de vie. Merci à Michelle Oakley
pour son support et sa grande aide lors des captures. Merci à elle et son conjoint Shane d’avoir participé aux
ixcaptures et de m’avoir invité chez eux au Yukon et en Alaska. Je suis très choyée d’avoir travaillé et partagé
des moments inoubliables avec des gens passionnés comme vous.
Je tiens aussi à remercier Cathy Bajzac pour son écoute et ses conseils ainsi que Julien H. Richard pour le
support en ce qui concerne la logistique du matériel de terrain, l’accompagnement sur le terrain ainsi que
pour l’exploration des nombreuses avenues que ce projet de maîtrise a emprunté au fil des ans. Votre
présence, en plus d’être grandement appréciée des étudiants, contribue à l’atmosphère rassembleuse du
labo Côté-Tremblay.
Je poursuivrais en remerciant tous les étudiants qui m’ont vu passer dans le laboratoire Côté-Tremblay. Sans
vous, mon expérience à la maîtrise aurait été beaucoup moins drôle, enrichissante et plaisante. Le labo Côté-
Tremblay, c’est de l’entraide entre collègues, mais aussi des amitiés qui auront rendu la vie plus douce durant
mes dernières années à Québec. Je veux donc remercier Martin Leclerc, Mael LeCorre, Emilie Champagne,
Barbara Vuillaume, Olivier Villemaire-Côté, Michael Bonin, Laurent De Vriendt, Sabrina Plante, Linda Novak,
Florent Déry, Béatrice Carrier, Marianne Gagnon, Jérémie Fuller, Frédéric Dulude-de Broin, Antoine
Boudreau-Leblanc, Aimie Baribeau, Béatrice Capolla, Baptiste Brault et Albert Michaud. Je voudrais aussi
remercier particulièrement mes maîtriseux. Vous avez été là pour toutes les étapes de cette maîtrise, que ce
soit pour des conseils, de l’aide, les fous rires sur l’heure du midi, les soirées, le badminton, les pomodoros
et vous êtes encore là malgré la distance. Evelyne et Alexis, mes compagnons dès la première session,
Sophiane ma grande complice révélée grâce à la pandémie (ça aura au moins permis ça!), Catherine ma
protégée/collègue d’orignaux et coloc de bureau et Rachel la dernière venue, redoutable addition au
regroupement de chasseresses Poivrées aux côtés de Sophiane, Evelyne et moi-même. Merci pour tout.
Durant toutes ces années, il y a eu les amitiés créées par la maîtrise, mais il y a aussi les amitiés qui
perdurent autant à Québec qu’à Mont-Laurier. Des amis qui ont su me soutenir, généralement
silencieusement, parce qu’après quelques mois, la maîtrise devient un sujet un peu tabou dans les soirées
entre amis. Merci pour vos mots d’encouragement et votre intérêt, mais surtout pour tous les moments
passés ensemble qui m’ont permis de m’évader le temps de quelques heures.
En terminant, je me dois de remercier ma famille qui, depuis toujours, constitue mon noyau qui est toujours
présent pour m’encourager et m’écouter lorsque j’en ai besoin. Je veux tout de même faire un merci spécial
à mes parents qui m’ont toujours soutenu dans mes études, qui m’ont particulièrement entendu parler de ce
projet de maîtrise et qui ont démontré un intérêt marqué pour mes avancements. Je remercie ma mère avec
xqui j’ai passé plusieurs heures au téléphone à partager les moments plus difficiles et mes victoires durant
ces dernières années. Merci également de m’avoir accueilli les fins de semaine pour changer le mal de place,
mais aussi pour me surveiller pendant que j’avançais ma rédaction. Sans toi, pour plusieurs raisons, je
n’aurais pas réussi à me rendre jusqu’ici et je te dois beaucoup. Finalement, je tiens à remercier mon père.
C’est lui qui m’a transmis sa passion de la nature à travers nos sorties de chasse, de pêche et de trappe. Il
aura été mon premier professeur de biologie et si ce n’était pas de lui, je n’aurais probablement jamais
poursuivi mes études dans cette branche. C’est avec toi que je préfère échanger mes observations, mes
questionnements et mes raisonnements sur tout ce qui concerne la nature même si ça arrive qu’on ne soit
pas d’accord! Merci pour tout, tout simplement.
xiAvant-propos
Ce mémoire de maîtrise s’intéressant aux effets du parasitisme par la tique d’hiver et des conditions
hivernales sur la condition corporelle de veaux orignaux dans deux régions du Nouveau-Brunswick rapporte
également l’efficacité d’un acaricide en application topique sur la charge de tique d’hiver. Il comporte quatre
sections : une introduction générale, deux chapitres principaux sous la forme d’articles scientifiques rédigés
en anglais ainsi qu’une conclusion générale. Les deux articles scientifiques seront soumis à des revues
scientifiques avec révision par les pairs.
Le premier article Evaluation of a topical acaricide to reduce winter tick loads in wild moose porte sur
l’efficacité d’un acaricide en application topique visant la manipulation de la charge de tiques d’hiver. La
manipulation de charge parasitaire est utile en recherche lorsqu’on tente d’isoler l’effet du parasite des effets
de l’environnement. Le deuxième article Variation in body condition of moose calves facing contrasting
winter severity and tick loads se penche quant à lui sur la variation des paramètres sanguins et de la
masse de deux groupes de veaux orignaux au Nouveau-Brunswick au cours de leur premier hiver. Les deux
groupes se situent sur un gradient latitudinal faisant en sorte que les charges de tiques ainsi que les
conditions hivernales ne sont pas les mêmes et affectent donc différemment les veaux orignaux des deux
régions. Les coauteurs de ses articles sont :
(1) Patrick Leighton, codirecteur de maîtrise, professeur agrégé à la Faculté de médecine vétérinaire,
Université de Montréal
(2) Steeve D. Côté, codirecteur de maîtrise, professeur titulaire au département de biologie de
l’Université Laval.
(3) Jean-Pierre Tremblay, directeur de maîtrise, professeur titulaire au département de biologie de
l’Université Laval.
J’ai réalisé la collecte de données, les analyses statistiques, l’interprétation des résultats et la rédaction des
deux articles en tant que première auteure en collaboration avec mon codirecteur Patrick Leighton
(professeurs agrégé à la Faculté de médecine vétérinaire, Université de Montréal), Steeve D. Côté
(professeur titulaire au Département de biologie de l’Université Laval) et mon directeur Jean-Pierre Tremblay
(professeur titulaire au Département de biologie de l’Université Laval). La contribution des auteurs au projet
est présentée dans le Tableau 1.
xiiTableau 1. Contribution des différents auteurs des deux articles présentés dans ce mémoire
D. De Pierre J-P. Tremblay P. Leighton S.D. Côté
Conception du projet X X X
Financement X X X
Collecte de données X
Analyses statistiques X X
Interprétation des résultats X X
Rédaction X
Révision X X X X
Approbation finale X X X X
*Julien Hénault-Richard a joué un rôle essentiel en ce qui concerne la logistique entourant ce projet notamment les
travaux de terrain auxquels il a aussi participé.
xiiiIntroduction générale
Les changements globaux d’origine anthropique ont des répercussions souvent négatives sur les
écosystèmes (Munson et al. 2018). Contrairement à bien des impacts de la présence humaine qui affectent
une région ou une population en particulier, les conséquences des changements climatiques se manifestent
à l’échelle planétaire. Un des impacts relié aux changements climatiques est la modification dans la
répartition spatiale de certaines espèces autant végétales qu’animales (Parmesan and Yohe 2003).
En effet, la modification des températures, de l’humidité et des régimes de précipitations peut affecter la
productivité (Wu et al. 2021) et les caractéristiques des habitats (Fan et al. 2014). Les populations animales
locales seraient alors susceptibles de se déplacer vers de nouvelles aires répondant davantage à leurs
besoins (Arevall et al. 2018). À long terme, on s’attend à ce que les températures plus chaudes favorisent
l’expansion de la distribution de nombreuses espèces vers les pôles (Conservation of Arctic Flora and Fauna
2013). La rapidité de réaction des espèces aux changements dépend des seuils de tolérances qui sont
propres à chaque espèce. Ainsi, des chevauchements entre les aires de distributions d’espèces qui ne
cohabitaient pas avant sont envisageables (Shepard et al. 2022).
Ces nouveaux chevauchements peuvent perturber les interactions interspécifiques comme les relations
prédateurs-proies ou même créer de la compétition entre des espèces dont les niches écologiques ne se
superposaient pas auparavant (Holm et al. 2016). Les déplacements de populations engendrés par le
réchauffement planétaire peuvent également avoir des répercussions sur les relations parasitaires. Les
parasites transportés par leurs hôtes en déplacement, comme des oiseaux en migration qui sont reconnus
pour être de grands vecteurs d’ectoparasites, pourraient être introduits dans de nouveaux environnements
(Waldenström et al. 2002, Sparagano et al. 2015). Les conséquences de l’introduction d’un parasite non
indigène à une région peuvent grandement affecter les populations animales n’ayant jamais eu de contact
avec le parasite et qui représentent des hôtes potentiels pour ce dernier (Morales-Castilla et al. 2021). Par
exemple, l’arrivée de Philornis downsi qui est une espèce de mouche introduite sur les iles Galapagos cause
de hauts taux de mortalité des couvées (16 à 95 % en 1998 et 2008) de plusieurs espèces de pinsons
communément appelés les pinsons de Darwin (Britton 2013).
Le parasitisme est une relation étroite entre un organisme et son hôte où celui-ci devient la principale source
de nourriture du parasite (Combes 1995, Poulin and Morand 2000), mais aussi son habitat. Au terme de la
coévolution qui s’opère entre le parasite et son hôte, il est attendu que l’équilibre atteint favorise la survie de
1l’hôte permettant ainsi au parasite d’en tirer un maximum de ressources (Combes 1997). Il en découle parfois
une coévolution entre l’hôte et son parasite et une course à l’armement (Richner 1998). Durant cette
évolution, l’hôte développe des défenses physiologiques et comportementales pour éviter l’infection alors
que le parasite tente de contourner ces défenses (Mendel et al. 2010). L’équilibre est en quelque sorte atteint
lorsque les défenses acquises par l’espèce hôte réussissent à combattre le parasite et ainsi lui permettre
d’accomplir ses activités habituelles sans pour autant l’éradiquer complètement, permettant à l’espèce
parasite de se reproduire et se maintenir dans l’environnement (Ebert and Hamilton 1996).
Il est possible de diviser les parasites en deux grandes classes ; les endoparasites qui s’installent dans les
tissus internes de l’animal et les ectoparasites qui se retrouvent sur la peau ou dans le poil de leurs hôtes
(Mehlhorn 2016). Parmi les ectoparasites, on retrouve des insectes piqueurs comme les moustiques, les
puces et les tiques. Ces dernières retiennent présentement l’attention en raison du fait qu’elles sont
responsables de la transmission de bactéries qui peuvent causer des maladies chez la faune transmissible
à l’humain ; des zoonoses (Nicholson et al. 2019). Un des exemples les plus connus est certainement celui
de la tique à pattes noires (Ixodes scapularis), vectrice de la bactérie Borrelia Burgdorferi qui est porteuse
de la maladie de Lyme (Brownstein et al. 2005). Cette tique se trouvait auparavant davantage aux États-
Unis, mais sa distribution ne cesse de s’étendre vers le nord dans la région des provinces de l’est (Leighton
et al. 2012). I. scapularis ne sera probablement pas le seul ectoparasite à étendre sa distribution dans ces
régions. On compte 9 espèces de tiques présentes au Québec, dont la tique étoilée d’Amérique (Amblyomma
americanum) qui semble en expansion (Villeneuve 2012) ainsi que Dermacentor variabilis dont la distribution
s’est déplacée notamment dans le nord-ouest en Saskatchewan (Dergousoff et al. 2013).
Il est difficile de prévoir comment les changements climatiques pourront favoriser ou non le déplacement des
espèces de tiques et dans quelle mesure, mais c’est une situation qui risque de se produire et qu’il est
nécessaire d’anticiper. Souvent, les cycles vitaux des parasites comprennent plusieurs stades et plusieurs
hôtes (Mehlhorn 2016). À certains stades vitaux, les parasites se retrouvent libres dans l’environnement et
sont alors vulnérables aux aléas climatiques (Hoar et al. 2012). Bien que les endoparasites possèdent des
stades vitaux libres, les ectoparasites sont davantage limités par la température ambiante et le climat. En
effet, la ponte des œufs, l’éclosion ainsi que la première période du stade larvaire ont généralement lieu au
sol plutôt qu’à l’intérieur de l’hôte comme chez les endoparasites (Mehlhorn 2016). Dans le cas précis des
tiques, si les conditions sont trop sèches, si la température est trop froide, ou si une couche de neige recouvre
le sol pendant les stades hors de l’hôte, il arrive qu’elles ne puissent pas survivre (Holmes et al. 2018). Ainsi,
2les changements climatiques impliquant des hivers plus courts favorisent l’apparition de zones propices à
l’établissement de certains parasites y compris des tiques pour qui l’environnement était autrefois trop froid
(Kutz et al. 2004). À l’inverse, des étés plus secs pourraient limiter les populations de tiques (Garner and
Wilton 1993). Ainsi, pour que des tiques réussissent à étendre ou déplacer leur aire de répartition, plusieurs
facteurs doivent se rencontrer et atteindre un équilibre.
Impacts comportementaux des parasites sur leurs hôtes
Outre la possibilité qu’elles soient vectrices d’une bactérie ou d’un virus, la présence de tiques sur un hôte
entraine des réponses chez ce dernier qui peuvent avoir des impacts sur sa condition corporelle. Par
exemple, plusieurs grands herbivores d’Amérique du Nord comme le bison (Bison bison), le wapiti (Cervus
canadensis) et le cerf de Virginie (Odocoileus virginianus) s’adonnent à des séances de toilettage dites
programmées (Mooring and Samuel 1998b, Mooring et al. 2006). En effet, il a été démontré que certaines
espèces passent plus de temps à se toiletter à l’automne, au moment où les tiques grimpent sur leur hôte,
et ce avant même que les tiques ne se soient nourries. Ce comportement n’est pas engendré par un stimulus
physique, car à ce stade, les tiques ne sont pas détectables par l’animal. Il s’agit plutôt d’un comportement
sélectionné au cours d’une coévolution, et qu’on dit alors programmé (Hart 2000). Cependant, il existe des
espèces pour lesquelles le toilettage sera plutôt déclenché par les stimuli provenant des morsures de tiques
(Hart 2000). Il se pourrait qu’on retrouve cette tactique chez les espèces d’hôtes naïfs qui n’ont pas co-évolué
avec des ectoparasites et qui cohabitent moins bien avec les tiques (Mooring and Samuel 1998a). Par
exemple, les morsures de tiques provoquent une irritation cutanée chez l’orignal (Alces alces) (Franzmann
and Schwartz 1998) et les démangeaisons qui en découlent peuvent enclencher une modification du
comportement de l’hôte, particulièrement un temps de toilettage accru (Barnard 1990). Ainsi, les individus
qui passent plus de temps à se toiletter ont conséquemment moins de temps à consacrer pour se nourrir,
être en vigilance ou se reposer (Skorupka 1999). Cette perturbation des activités de l’hôte par les
ectoparasites, dont les tiques, et leur consommation de sang entrainent d’autres conséquences sur la santé
de leurs hôtes.
Impact des parasites sur la santé et la condition corporelle de leurs hôtes
Plusieurs études sur des animaux domestiques ou en captivité rapportent des pertes de masse ou de
moindres gains de masse chez les individus infestés par des tiques (Corrier et al. 1979, Glines and Samuel
1989). Par exemple, Little (1963) a observé qu’une charge quotidienne de 50 tiques (Boophilus microplus)
pendant 16 semaines pouvait créer une réduction annuelle du taux de croissance de 3674 g par tique chez
3des génisses (Bos primigenius). Cela peut s’expliquer à la fois par la perte de sang entrainant une dépense
d’énergie supplémentaire pour le renouvellement des globules rouges (Musante et al. 2007) et par la
perturbation des activités habituelles due au toilettage engendré par les morsures de tiques (Skorupka 1999).
Tel qu’observé par Musante et al. (2007), les veaux orignaux souffrant d’une sévère infestation par la tique
d’hiver (Dermacentor albipictus) (70 000 tiques) pouvaient perdre l’équivalent de l’énergie généralement
métabolisée en 3 à 5 jours durant la période d’engorgement des tiques adultes femelles (8 semaines). Cette
situation est particulièrement inquiétante chez les populations vivant dans des environnements subissant
des variations saisonnières extrêmes pour lesquelles la survie est généralement étroitement liée aux
réserves lipidiques (Monteith et al. 2013). En effet, les périodes associées à une réduction de la disponibilité
des aliments, comme l’hiver, font en sorte que les animaux doivent accumuler des réserves durant la période
d’abondance (Monteith et al. 2013). Ce sont les réserves de gras sous-cutanées qui sont les premières à
être mobilisées (Mautz 1978). Le gras stocké sous forme de triglycérides contient plus de deux fois l’énergie
contenue dans la même quantité de muscles (Robbins 1983). Dans le cas où un animal ne réussit pas à
accumuler suffisamment de réserves pour l’hiver, l’organisme peut alors puiser l’énergie stockée dans la
masse musculaire (Kiessling and Kiessling 1984). La dégradation des fibres musculaires s’accompagne de
certains déchets métaboliques comme la créatinine. Ainsi, plus un animal dégrade de masse musculaire,
plus le niveau de créatinine sanguin augmentera (Delgiudice et al. 1992). Cependant, cette augmentation de
créatinine est temporaire puisqu’une fois les muscles atrophiés, les taux de créatinine chutent (Czyzowski et
al. 2013, Rosenblatt et al. 2021).
Outre la perte de poids, il est possible d’obtenir des informations sur le parasite et ses effets par un bilan
sanguin de l’hôte (Fidelis et al. 2016). Dans ce contexte, le terme « bilan sanguin » réfère à l’ensemble des
paramètres qui peuvent être évalués à la suite d’une prise de sang et qui renseignent sur l’état de santé de
l’individu. Par exemple, des infestations par la tique ont mené à l’observation d’hématocrite, de taux
d’hémoglobine, de quantité de globules rouges et de protéines plasmatiques moins élevés chez les hôtes
davantage infestés (Little 1963, Addison et al. 1998b, Musante et al. 2007). Ces changements au niveau du
bilan sanguin de l’hôte ont notamment été liés à des problèmes d’anémie chez les animaux parasités (Jellison
and Kohls 1938, Corrier et al. 1979, Musante et al. 2007). On peut détecter une anémie par l’hématocrite
c’est-à-dire le volume occupé par les globules rouges sur le volume de sang total ou le niveau d’hémoglobine
dans le sang (Billet 1990). On peut distinguer plusieurs types d’anémies qui se différencient les unes des
autres par la cause et les répercussions sur les composantes sanguines. Par exemple, on peut établir un
diagnostic sur la base de la taille des globules rouges, aussi appelé le volume globulaire moyen (VGM). La
4taille des globules rouges peut être en dessous de la moyenne, dans la moyenne ou au-dessus de la
moyenne, selon ce résultat, on identifiera une anémie microcytaire, normocytaire ou macrocytaire (Conrad
1990). Une anémie microcytaire sous-entend que la production de globules rouges est freinée, souvent par
un manque de fer (Conrad 1990). On peut aussi séparer les anémies régénératives des anémies
arégénératives. On parle d’une anémie arégénérative lorsque la cause de l’anémie affecte la production de
globules rouges et que le corps ne peut donc pas compenser le faible nombre de globules rouges en en
produisant de nouveaux (Bessman 1990). Au contraire, lorsque la cause de l’anémie n’est pas en lien avec
la production de globules rouges, mais plutôt régie par une perte de globules rouges comme une hémorragie
ou de l’hémolyse, le corps tente de compenser cette perte (Bessman 1990). La moelle osseuse tente alors
de compenser la perte de globules rouges en en produisant de nouveaux (Schalm 1961). À ce moment, on
peut observer un taux de réticulocytes, i.e. jeunes globules rouges, plus élevé que la normale. C’est ce qui
a été observé entre autres dans une étude de Cériac et al. (2017) alors que des chèvres infestés par un
nématode gastro-intestinal (Haemonchus contortus) présentaient une anémie régénérative avec de hauts
taux de réticulocytes.
Contrôle des tiques
Étant donné la proximité de l’humain aux animaux domestiques, mais aussi la densité de population des
élevages qui favorise les infestations, plusieurs d’études sur le parasitisme ont été réalisées sur les animaux
d’élevage. Pour pallier les conséquences du parasitisme sur les animaux domestiques qui affecte la
productivité du troupeau, les éleveurs doivent intervenir. Une des premières solutions employées fut
l’utilisation de mélanges contenant du sulfure, du kérosène, du pétrole et diverses huiles appliqués sur le
corps des animaux à l’aide de linges imbibés des mélanges (Mohler 1906). De nos jours, il existe une grande
variété de pesticides qui varient en fonction de l’ectoparasite visé, du mode d’action et de la méthode
d’administration (George et al. 2004). Alors que certains éléments actifs s’attaquent directement au système
nerveux des invertébrés, d’autres interfèrent dans le processus de respiration mitochondriale ou dans le
développement et la croissance des organismes (Dekeyser 2005). Pour atteindre les tiques, certains
acaricides s’administrent par injection ou par voie orale et sont alors consommés par la tique lorsqu’elle se
nourrit du sang de l’hôte (Nicholson et al. 2019). D’autres sont appliqués directement sur la peau de l’hôte
sous forme de solution plus ou moins diluée ou via une étiquette apposée sur l’oreille et agissent lorsqu’en
contact direct avec un invertébré (George et al. 2008). Comme les tiques s’attaquent autant aux animaux
domestiques que sauvages, certains acaricides ont été testés sur la faune en nature (Solberg et al. 2003,
5Carroll et al. 2009, Grear et al. 2014). Un dispositif qui fut employé par le passé est celui impliquant des
mangeoires pour cerfs de Virginie équipés de rouleaux imbibés d’acaricide. Le cerf doit nécessairement
insérer sa tête entre les rouleaux pour avoir accès à la nourriture et il s’applique le traitement sur la tête et le
cou par la même occasion. Divers acaricides ont été utilisés comme l’amitraz et la perméthrine qui s’attaquent
au système nerveux (Page 2008). Les études utilisant ce dispositif rapportent des résultats concluants quant
au contrôle local des populations de tiques à pattes noires (Solberg et al. 2003, Carroll et al. 2009, Grear et
al. 2014).
Impact des conditions hivernales sur la condition corporelle
Bien que les animaux domestiques et les animaux sauvages peuvent être parasités, les animaux sauvages
ne reçoivent pas de traitement pour les soulager. D’ailleurs, en plus de subir les conséquences du
parasitisme présentées plus haut, ils doivent en plus composer avec les impacts de l’environnement. Les
parasites ne sont pas les seuls facteurs influant sur la condition corporelle des organismes hôtes et on peut
considérer le parasitisme comme un facteur s’ajoutant aux conditions qui affectent d’emblée la santé des
animaux. Les principaux facteurs affectant le bilan énergétique d’un individu sont la quantité et la qualité de
la nourriture ingérée ainsi que le mouvement et les déplacements (Hudson and White 1985). Il existe
plusieurs paramètres qui peuvent causer un déséquilibre du bilan énergétique. Par exemple, la compétition
pour la nourriture entre les individus ou les espèces peut impacter négativement leur alimentation (Wauters
et al. 2000) alors que la prédation peut quant à elle induire plus de mouvement chez des proies (Mitchell and
Lima 2002) et réduire le temps alloué à l’alimentation (Desportes et al. 1990). On peut aussi penser au climat
qui, lors d’une sécheresse, réduit la nourriture chez les herbivores (Cain et al. 2017) ou tout simplement à la
saison hivernale qui diminue l’accessibilité à de la nourriture de qualité (Massé and Côté 2012) en plus
d’augmenter les dépenses énergétiques liées aux déplacements dans la neige (Crampe et al. 2002) et à la
thermorégulation (Renecker and Hudson 1986). Si on s’attarde aux herbivores, les feuilles et les bourgeons
qui sont à leurs plus nutritifs au début de l’été laissent place à de la matière végétale moins nutritive et plus
difficile à assimiler (Parikh et al. 2017). De plus, l’augmentation des dépenses énergétiques associées aux
déplacements dans la neige de certaines espèces d’ongulés comme le wapiti et le cerf mulet (Odocoileus
hemionus) a été détaillée dans le papier de Parker et al. (1984). Si on ajoute cette dépense d’énergie à de
la nourriture plus rare et moins nutritive, on peut assister à une perte de masse chez les individus (Mautz
1978).
6Ainsi, on est à même de constater que les animaux parasités faisant également face à des conditions
environnementales difficiles pourraient subir les conséquences combinées de ces deux facteurs. Ce concept
est d’ailleurs illustré grâce au triangle épidémiologique (Figure 1) qui représente les liens reliant les
populations hôtes à leur charge parasitaire ainsi qu’aux conditions environnementales (Antonovics 2017).
Lorsqu’on souhaite identifier les effets du parasitisme sur la santé des hôtes, il importe d’avoir des individus
avec des charges parasitaires variables. Un moyen d’y arriver pourrait être d’étudier des hôtes dans plusieurs
régions reconnues pour avoir des charges parasitaires contrastées. On peut, par exemple, sélectionner des
régions le long d’un gradient latitudinal étant donné que les parasites sont affectés par les conditions
environnementales et risquent donc d’être moins abondants en climat froid. Cependant, dans ce contexte,
les groupes d’hôtes font potentiellement face à des conditions environnementales distinctes qui peuvent les
affecter. C’est pourquoi l’utilisation d’antiparasitaires sur des hôtes au sein d’une même population faisant
face aux mêmes conditions climatiques est un moyen efficace d’évaluer les conséquences du parasitisme
sur les hôtes (Albon et al. 2002, Devevey and Christe 2009). On peut ainsi manipuler la charge parasitaire
en traitant une partie des individus et en maintenant des individus témoins afin d’isoler les conséquences du
parasitisme.
Figure 1. Adaptation du triangle épidémiologique à partir de la représentation graphique par Antonovics (2017)
7Modèles de l’étude
L’orignal est le plus gros cervidé mammifère présent en Amérique du Nord. C’est un cervidé bien adapté aux
conditions hivernales neigeuses qu’on retrouve donc plus au nord que plusieurs autres espèces de cervidés.
Contrairement au cerf de virginie, sa grande taille lui permet de plus facilement préserver sa chaleur
(Bergmann 1848) et ses longues pattes fortes facilitent ses déplacements dans la neige (Bubenik 1998).
Cela ne veut pas dire que les orignaux ne ressentent pas les effets de l’hiver, particulièrement les jeunes
nés au cours du printemps précédent. Alors que les adultes accumulent des réserves en prévision de l’hiver,
les jeunes orignaux ont utilisé une grande partie de l’énergie ingérée au cours de leur courte vie à leur
croissance (Schwartz et al. 1991). D’ailleurs, bien qu’ils aient subi une poussée de croissance au cours des
mois précédents l’hiver, ils ne sont quand même pas aussi grands que leurs congénères adultes. Ainsi, ils
font face à l’hiver avec des défis plus grands que les adultes, spécialement en ce qui a trait à leurs réserves
d’énergie qui sont minces.
En plus des défis prévisibles auxquels font face les orignaux et principalement les jeunes, s’ajoute l’effet d’un
parasite qui pourrait être en expansion dans la province. Il s’agit de la tique d’hiver qui se distingue des autres
tiques par le fait qu’elle est principalement active durant la saison hivernale (Figure 2) mais aussi, car elle ne
possède généralement qu’un seul hôte. Qui plus est, elle ne vit qu’une seule année (Samuel 2004c) alors
que chez beaucoup d’espèces de tiques il y a deux ou trois hôtes différents associée aux trois stades vitaux
(larve, nymphe et adulte ; annexe 1) qui s’étendent sur un cycle vital de plus de 2 ans (Bowman and Nuttall
2008). La tique d’hiver s’accroche aux poils de son hôte sous forme de larve et s’alimente de sang avant ses
changements de stades ainsi qu’avant la ponte des œufs, et ce sur le même individu (Samuel 2004c). Ce
mode de vie hivernal amplifie l’effet de la tique sur l’orignal parce qu’il s’agit d’une période critique pour celui-
ci (Schwartz and Renecker 1998). Les ressources sont limitantes en hiver en Amérique du Nord et les
orignaux doivent faire des compromis face aux facteurs limitants que sont la disponibilité des ressources,
mais aussi la profondeur de neige et la prédation (Dussault et al. 2005). Cette situation fait que les orignaux
doivent généralement puiser dans leurs réserves pour survivre à l’hiver (Moen et al. 1997).
8Vous pouvez aussi lire

















































