Les champignons ectomycorhiziens des forêts naturelles et des plantations d'Afrique de l'Ouest : une source de champignons comestibles
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
51
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
Les champignons ectomycorhiziens
des forêts naturelles et des
plantations d’Afrique de l’Ouest :
une source de champignons
comestibles
Marc Ducousso
Laboratoire
des symbioses tropicales
et méditerranéennes
TA 10/J
34398 Montpellier Cedex
France
Amadou Moustapha Bâ
Université des Antilles
et de la Guyane
Pointe-à-Pitre Peu de données sont actuellement disponibles sur les champignons
Guadeloupe ectomycorhiziens comestibles en Afrique de l’Ouest, alors qu’ils constituent une source de
France revenus potentielle pour les populations. Des recherches permettant d’allier une gestion
forestière durable à la production de champignons comestibles, notamment en plantations,
Daniel Thoen devraient être entreprises dans cette partie du continent africain.
Fondation universitaire
luxembourgeoise
185, avenue de Longwy
6700 Arlon
Belgique
Lactarius gymnocarpoides, espèce de champignon comestible mentionnée en Afrique centrale
et australe et également trouvée en Afrique de l’Ouest.
Lactarius gymnocarpoides, species of edible fungus reported in central and southern Africa is
also found in West Africa.
Photo : D. Thoen.
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
52 Marc Ducousso
FOCUS / ECTOMYCORRHIZAL FUNGI
Amadou Moustapha Bâ
Daniel Thoen
RÉSUMÉ ABSTRACT RESUMEN
LES CHAMPIGNONS ECTOMYCORRHIZAL FUNGI LOS HONGOS ECTOMICORRÍCICOS
ECTOMYCORHIZIENS DES FORÊTS ASSOCIATED WITH NATIVE AND DE LOS BOSQUES NATURALES Y DE
NATURELLES ET DES PLANTATIONS PLANTED TREE SPECIES IN WEST LAS PLANTACIONES DE ÁFRICA
D’AFRIQUE DE L’OUEST : UNE AFRICA: A POTENTIAL SOURCE OF OCCIDENTAL: UNA FUENTE DE
SOURCE DE CHAMPIGNONS EDIBLE MUSHROOMS HONGOS COMESTIBLES
COMESTIBLES
West African forests extend from Los bosques de África Occidental se
Les forêts d’Afrique de l’Ouest s’éten- southern Senegal to Côte d’Ivoire. In extienden desde el sur de Senegal
dent depuis le sud du Sénégal jus- these forests, ectomycorrhizal trees hasta Costa de Marfil. En estos bos-
qu’en Côte d’Ivoire. Dans ces forêts, are locally diversified and/or abun- ques, los árboles ectomicorrícicos se
les arbres ectomycorhiziens sont dant. They belong to the following encuentran localmente diversificados
localement diversifiés et/ou abon- botanical families: Caesalpiniaceae o abundantes. Pertenecen a las
dants. Ils appartiennent aux familles (Tribes Detariae and Amherstieae), siguientes familias botánicas:
botaniques des Caesalpiniaceae (tri- Dipterocarpaceae (Genus Monotes) Caesalpiniaceae (tribus Detarieae y
bus des Detarieae et des Amhers- and Uapacaceae (Genus Uapaca). The Amherstieae), Dipterocarpaceae
tieae), Dipterocarpaceae (genre ectomycorrhizal fungi linked to these (género Monotes) y Uapacaceae
Monotes) et Uapacaceae (genre trees are highly diverse, and include (género Uapaca). La diversidad de
Uapaca). La diversité des champi- many species of Russula, Lactarius, hongos ectomicorrícicos asociados a
gnons ectomycorhiziens associés à Amanita, Cantharellus, Boletus (s.l.). estos árboles es importante, espe-
ces arbres est importante, notam- Many Russula, Amanita and Cantha- cialmente entre los géneros Russula,
ment parmi les genres Russula, Lacta- rellus species of West Africa are defi- Lactarius, Amanita, Cantharellus y
rius, Amanita, Cantharellus et Boletus nitely edible, as most of them are Boletus (s. l.). Numerosas especies
(s. l.). De nombreuses espèces pré- consumed elsewhere, in Central, presentes en África Occidental, perte-
sentes en Afrique de l’Ouest, appar- Eastern or Southern Africa. However, necientes a los géneros Russula,
tenant aux genres Russula, Amanita in West Africa, consumption of edible Amanita y Cantharellus son, proba-
et Cantharellus, sont probablement fungi by local peoples is not yet well blemente, comestibles ya que se con-
comestibles car elles sont consom- documented. As non-timber forest sumen en África Oriental y en África
mées en Afrique de l’Est et en Afrique products, fungi are under-exploited in Austral. Sin embargo, en África
australe. Cependant, en Afrique de this part of Africa. It can also be Occidental se dispone de muy poca
l’Ouest, très peu d’information est assumed that, as elsewhere in Africa, información sobre el consumo de
disponible sur la consommation des people living in or near forest areas hongos comestibles por las poblacio-
champignons comestibles par les are likely to be more tolerant of fungi nes locales. Al igual que la mayoría
populations locales. Comme la plu- than savannah people, who are gen- de los productos forestales no leño-
part des produits forestiers non erally mycophobic. sos, los hongos están subexplotados
ligneux, les champignons sont sous- en esta parte de África. También es
exploités dans cette partie de Keywords: ectomycorhizal fungi, edi- probable que, como en otras partes
l’Afrique. Il est aussi probable que, ble fungi, natural forest, plantation, de África, los habitantes de zonas
comme ailleurs en Afrique, les popu- West Africa. forestales o colindantes estén más
lations vivant dans ou au voisinage familiarizados con los hongos que los
des zones forestières soient plus de las sabanas, que suelen ser micó-
familières avec les champignons que fobos.
celles des savanes qui sont générale-
ment mycophobes. Palabras clave: hongo ectomicorrí-
cico, hongo comestible, bosque natu-
Mots-clés : champignon ectomycorhi- ral, plantación, África Occidental.
zien, champignon comestible, forêt
naturelle, plantation, Afrique de
l’Ouest.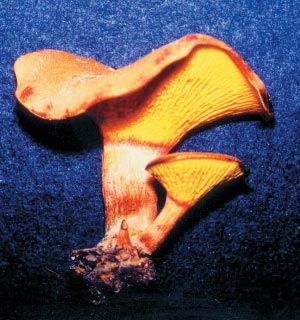
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
53
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
Introduction Situation
géographique
De nombreuses espèces de second temps, de relier ces sources de la zone d’étude
champignons ectomycorhiziens sont potentielles aux récentes données
des comestibles de valeur. Parmi sur les champignons comestibles des
celles-ci, la truffe noire du Périgord, parties centrale, orientale et australe Notre propos se limite à la par-
le matsutake, les chanterelles et les de l’Afrique. Pour cela, deux éléments tie occidentale de l’Afrique de
bolets sont, mondialement, les plus sont à prendre en compte. Le premier l’Ouest, entre 5 et 10° Nord et 3 et 16°
connus des champignons forestiers. est la disponibilité de données Est. Ces limites incluent le sud du
Cependant, la plupart de nos don- récentes sur les champignons comes- Sénégal et du Burkina, la Côte
nées viennent des pays tempérés. tibles en Afrique centrale et australe, d’Ivoire et la Guinée, pays pour les-
Très peu de connaissances sont le second est constitué par nos quels des informations sont dispo-
actuellement disponibles sur la connaissances, acquises depuis plus nibles. Nos conclusions pourront être
comestibilité des champignons tropi- de vingt ans, sur les champignons extrapolées aux pays voisins, comme
caux ; cela est particulièrement vrai ectomycorhiziens en Afrique de le Bénin et le Nigeria, pour les parties
en Afrique de l’Ouest, même si une l’Ouest. La synthèse de ces deux élé- de ce pays ayant un climat et une
étude sur les champignons ectomyco- ments nous permettra de mettre en végétation similaires (figure 1). En
rhiziens comestibles a été récemment évidence les possibilités de produc- effet, même si quelques spécificités
entreprise au Bénin (De Kesel et al., tion de champignons ectomycorhi- existent au sein de la flore ectomyco-
2001). ziens comestibles en Afrique de rhizienne ouest-africaine, nous
Les champignons comestibles l’Ouest. n’avons, jusqu’à présent, aucune
sont mieux connus en Afrique cen-
trale et en Afrique de l’Est (notam-
ment Parent, Thoen, 1978 ; Buyck,
1994 ; Harkönen et al., 1995). Il faut
dire que leur découverte est beau-
coup plus ancienne. En effet, c’est en
1867 que David Livingstone rapporte
pour la première fois, en Afrique, la
consommation de quantités impor-
tantes de champignons en Zambie
(Piearce, 1985). Ce dernier auteur
précise que les champignons cités
par Livingstone étaient des espèces
ectomycorhiziennes. Dans une syn-
thèse récente sur les champignons
comestibles africains (Rammeloo,
Walleyn, 1993), 151 références sont
citées. Seulement trois concernent
l’Afrique de l’Ouest (Heim, 1936 a Pays étudiés :
et b ; Locquin, 1954). Aucun travail
récent traitant des champignons Sénégal, Guinée, Côte d’Ivoire,
comestibles en Afrique de l’Ouest Burkina Faso
n’est disponible. Ce manque de don-
nées n’est pas dû à une absence de
champignons dans cette partie de Pays où les résultats sont
l’Afrique. En effet, les champignons extrapolables
sont régulièrement mentionnés dans
les régimes alimentaires des popula-
tions locales, sans plus de précisions Répartition de la forêt africaine
sur les espèces, leurs origines et les à ectomycorhizes
méthodes de cuisson.
Notre objectif est, dans un pre-
mier temps, de décrire les ressources Absence de champignons
en champignons ectomycorhiziens en ectomycorhiziens
Afrique de l’Ouest puis, dans un Figure 1.
Pays étudiés et zones où les observations présentées peuvent être extrapolées.
Studied countries and other areas to which these observations may be extrapolated.
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
54
FOCUS / ECTOMYCORRHIZAL FUNGI
A B
C D
Figure 2.
Les quatre types de formations forestières ouest-africaines dominées par les ectomycorhizes. A : forêt tropicale dense humide
au sud de la Guinée ; B : forêt soudanienne claire semi-décidue à Lataha, Côte d’Ivoire ; C : forêt-galerie vue d’avion au nord de
la Côte d’Ivoire ; D : plantations d’Eucalyptus au nord de la Côte d’Ivoire.
The four types of West African forest formations with dominant ectomycorrhiza. A: dense tropical rain forest in southern Guinea;
B: Sudanese semi-deciduous clear forest, Lataha, Côte d’Ivoire; C: aerial photograph of a gallery forest in the north of Côte
d’Ivoire; D: Eucalyptus plantations in the north of Côte d’Ivoire.
Photo A : A. M. Bâ ; photos B, C, D : M. Ducousso.
indication quant à l’existence de dif- Les pr incipaux sée par une pluviosité annuelle com-
férences majeures qui la sépareraient prise entre 2 000 et 6 000 mm, une
du reste de la flore ectomycorhi-
types forestier s humidité maximale tout au long de
zienne de l’Afrique tropicale. concer nés l’année et une saison sèche quasi
Toutefois, il convient d’être très pru- inexistante, favorable à l’abondance
dent et de ne pas considérer qu’une Dans la zone d’étude définie, des épiphytes. Le deuxième type est
espèce est comestible dans tous les quatre types de forêts dominées par la forêt soudanienne claire semi-déci-
pays où on la trouve si les informa- les ectomycorhizes sont à considérer. due (figure 2 b). Ce type de forêt est
tions concernant sa comestibilité ne Le premier type est la forêt tropicale souvent dominé par une seule espèce
sont avérées que dans un seul pays. dense (figure 2 a). Elle est caractéri- formant des peuplements purs (cas
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
55
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
A B
C D
E F
Figure 3.
Les six espèces de champignons comestibles mentionnées en Afrique centrale et australe et également trouvées en Afrique de
l’Ouest. A : Cantharellus rufopunctatus ; B : Cantharellus pseudofriesii ; C : Lactarius gymnocarpus ; D : Lactarius
gymnocarpoides ; E : Phlebopus sudanicus et F : Tubosaeta brunneosetosa.
Six species of edible fungi reported in central and southern Africa are also found in West Africa. A: Cantharellus rufopunctatus;
B: Cantharellus pseudofriesii; C: Lactarius gymnocarpus; D: Lactarius gymnocarpoides; E: Phlebopus sudanicus and F:
Tubosaeta brunneosetosa.
Photos A, B : M. Ducousso. ; photos C, D : D. Thoen ; photo E : M. Ducousso ; photo F : D. Thoen.
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
56
FOCUS / ECTOMYCORRHIZAL FUNGI
Tableau I.
Liste des espèces d’arbres à ectomycorhizes rencontrées en Afrique de l’Ouest mentionnant
leur écologie, le pays où la symbiose a été décrite et les références bibliographiques.
Espèce Famille/Tribu Écologie Pays/Auteurs
Afzelia africana Caesalpiniaceae/Detarieae Forêt claire soudanienne, Sénégal (Thoen, Bâ, 1989),
forêt tropicale dense Guinée (Thoen, Ducousso, 1989 a),
Burkina (Sanon et al., 1997),
Côte d’Ivoire (Ducousso et al., 1999)
Afzelia bella Caesalpiniaceae/Detarieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Afzelia bracteata Caesalpiniaceae/Detarieae Forêt claire soudanienne Guinée (Thoen, Ducousso, 1989 a)
Anthonotha crassifolia Caesalpiniaceae/Amherstieae Forêt claire soudanienne Guinée (Thoen, Ducousso, 1989 a)
Anthonotha fragans Caesalpiniaceae/Amherstieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Anthonotha macrophylla Caesalpiniaceae/Amherstieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Berlinia grandiflora Caesalpiniaceae/Amherstieae Forêt claire soudanienne Côte d’Ivoire (Locquin, 1954)
Cryptosepalum tetraphyllum Caesalpiniaceae/Amherstieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Gilbertiodendron limba Caesalpiniaceae/Amherstieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Isoberlinia doka Caesalpiniaceae/Amherstieae Forêt claire soudanienne Burkina (Sanon et al., 1997),
Côte d’Ivoire (Ducousso et al., 1999)
Isoberlinia dalziellii Caesalpiniaceae/Amherstieae Forêt claire soudanienne Burkina (Sanon et al., 1997)
Paramacrolobium coeruleum Caesalpiniaceae/Amherstieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Pellegriniodendron diphyllum Caesalpiniaceae/Amherstieae Forêt tropicale dense Guinée (Bâ et al., 2000)
Monotes kerstingii Dipterocarpaceae/Monotideae Forêt claire soudanienne Burkina (Sanon et al., 1997)
Uapaca chevalieri Uapacaceae Forêt claire soudanienne Guinée (Thoen, Ducousso, 1989 a)
Uapaca guineensis Uapacaceae Forêt-galerie, Sénégal (Thoen, Bâ, 1989),
forêt tropicale dense Guinée (Thoen, Ducousso, 1989 a),
Burkina (Sanon et al., 1997),
Côte d’Ivoire (Ducousso et al., 1999)
Uapaca heudelottii Uapacaceae Forêt tropicale dense Côte d’Ivoire (Ducousso et al., 1999),
Guinée (Bâ et al., 2000)
Uapaca somon Uapacaceae Forêt tropicale dense, Burkina (Sanon et al., 1997),
forêt claire soudanienne Guinée (Bâ et al., 2000)
Uapaca togoensis Uapacaceae Forêt claire soudanienne Côte d’Ivoire (Ducousso et al., 1999)
Uapaca sp. Uapacaceae Forêt-galerie Guinée (Thoen, Ducousso, 1989 a),
Burkina (Sanon et al., 1997)
d’Isoberlinia doka au nord de la Côte de la tribu des Amherstieae. Le qua- Les espèces
d’Ivoire). Dans ces régions, la pluvio- trième type de boisement est formé d’arbres impliquées
sité annuelle varie de 800 à 2 000 mm par les plantations (figure 2 d). On les
et les épiphytes deviennent rares. Le trouve à peu près partout, étant faci-
dans les symbioses
troisième type de forêt est constitué lement reconnaissables, essentielle- ectomycorhiziennes
par les galeries forestières (figure ment du fait de leur caractère généra-
2 c). Ces formations particulières lement monospécifique. Quelques Les trois premiers types de forêt
croissent le long des cours d’eau tem- espèces de plantations comme les décrits ci-dessus sont des formations
poraires ou permanents, principale- Eucalyptus, les Acacia australiens et naturelles d’Afrique de l’Ouest. Les
ment dans la zone soudanienne de les pins sont ectomycorhiziennes et arbres ectomycorhiziens de ces forêts
l’Afrique de l’Ouest. Elles sont domi- maintenant très répandues en Afrique appartiennent à trois familles bota-
nées principalement par les Uapaca, de l’Ouest. niques : Caesalpiniaceae, Dipterocar-
les Afzelia et quelques autres genres paceae et Uapacaceae (tableau I).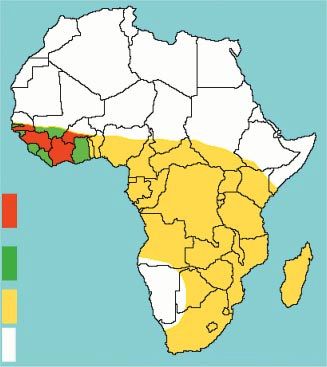
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
57
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
Tableau II. Liste des espèces d’arbres ectomycorhiziens introduites
en Afrique de l’Ouest avec mention du pays d’introduction et des
références bibliographiques ayant rapporté leur ectomycorhization.
Espèce Famille Pays/Auteurs
Casuarina equisetifolia Casuarinaceae Sénégal (Bâ et al., 1987)
Cinnamomum zeylanicum Lauraceae Guinée (Thoen, Ducousso, 1989 a)
Eucalyptus apodophyla Myrtaceae Sénégal (Ducousso, 1991)
Parmi les Caesalpiniaceae, deux
tribus, les Amherstieae et les Eucalyptus camaldulensis Myrtaceae Sénégal (Ducousso, 1991),
Detarieae, ont des espèces à ectomy- Guinée (Thoen, Ducousso, 1989 a),
corhizes. La première possède un Burkina (Thoen et al., 2001),
Côte d’Ivoire (Lesueur, Ducousso, 1995)
nombre relativement important de
genres et d’espèces en Afrique de Eucalyptus citriodora Myrtaceae Sénégal (Ducousso, 1991)
l’Ouest tandis que la seconde n’est Eucalyptus grandis Myrtaceae Sénégal (Ducousso, 1991)
représentée que par un seul genre,
Afzelia. Eucalyptus pentaleuca Myrtaceae Sénégal (Ducousso, 1991)
Le genre Uapaca a longtemps Eucalyptus tereticornis Myrtaceae Sénégal (Ducousso, 1991)
été classé parmi les Euphorbiaceae.
Au moins six espèces d’Uapaca sont Melaleuca leucodendron Myrtaceae Sénégal (Ducousso, 1991)
présentes en Afrique de l’Ouest. Melaleuca viridifolia Myrtaceae Sénégal (Ducousso, 1991)
Les Monotoideae constituent la
Acacia auriculiformis Mimosaceae Sénégal (Ducousso, 1991),
tribu africaine des Dipterocarpaceae.
Côte d’Ivoire (Lesueur, Ducousso, 1995)
Une seule espèce du genre Monotes
a été décrite comme ectomycorhi- Acacia holosericea Mimosaceae Sénégal (Ducousso, 1991),
zienne en Afrique de l’Ouest. Côte d’Ivoire (Lesueur, Ducousso, 1995)
D’autres espèces ouest-afri- Acacia mangium Mimosaceae Sénégal (Ducousso, 1991),
caines, non mentionnées dans le Côte d’Ivoire (Lesueur, Ducousso, 1995)
tableau I, sont probablement aussi à
Acacia trachycarpa Mimosaceae Sénégal (Ducousso, 1991)
ectomycorhizes. Cela est certaine-
ment vrai pour d’autres espèces des Pinus caribaea Pinaceae Sénégal (Ducousso, 1991),
genres déjà cités dans le même Côte d’Ivoire (Lesueur, Ducousso, 1995)
tableau, mais aussi pour des genres Pinus kesiya Pinaceae Guinée (Ducousso, 1991)
comme Aphanocalyx, Didelotia,
Microberlinia, Monopetalanthus et Pinus patula Pinaceae Guinée (Ducousso, 1991)
Tetraberlinia (Newbery et al., 1988),
dont la capacité à former des ectomy-
corhizes n’a pas encore été recher- La comestibilité : considérés comme non comestibles
chée en Afrique de l’Ouest. un concept lorsqu’ils présentent un caractère
D’après la Fao (Pandey, 1997), toxique. La toxicité peut être expri-
les plantations de pins, d’Eucalyptus, complexe mée par des désordres physiolo-
d’Acacia et de Casuarina, genres giques plus ou moins importants,
connus pour leur capacité à former Une approche négative du principalement d’ordre digestif ou
des ectomycorhizes, couvrent plus de concept de comestibilité peut aider à neuroleptique. Bien sûr, les champi-
100 000 ha en Afrique de l’Ouest. sa meilleure compréhension. Un gnons mortels ne sont jamais consi-
Cela ne représente que 0,1 % de la champignon peut être considéré dérés comme comestibles et, le plus
surface du Burkina, de la Côte d’Ivoire comme non comestible lorsque, souvent, les espèces proches sont
et du Sénégal réunis. D’autres genres notamment, sa consistance est trop également rejetées. Les champignons
à ectomycorhizes ont été plantés en dure ou trop visqueuse. Certains ayant des fructifications de petite
Afrique de l’Ouest, mais les surfaces champignons trop durs comme taille ainsi que les champignons fruc-
existant actuellement sont peu Ganoderma lucidum (Jordan, tifiant rarement ou très peu sont éga-
importantes. Une liste des espèces Wheeler, 1998) et quelques autres lement négligés du fait de la difficulté
utilisées en plantation en Afrique de Polyporaceae sont utilisés comme de leur récolte.
l’Ouest, pour lesquelles des ectomy- remèdes en médecine traditionnelle Dans une approche positive, les
corhizes ont été décrites, est présen- (notamment Gérault, Thoen, 1992). deux principales qualités d’un cham-
tée dans le tableau II. Les champignons sont également pignon comestible résident dans son
BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
58
FOCUS / ECTOMYCORRHIZAL FUNGI
70 espèces associées principalement à
Isoberlinia doka ont été décrites
(Ducousso et al., 1999). En Guinée, 38
espèces ont été trouvées au Fouta-Dja-
lon (Thoen, Ducousso, 1989 a) et 110
dans la forêt dense humide du sud du
pays (Rivière et al., 2001). Au Sénégal,
43 espèces ont été décrites comme
associées à Uapaca guineensis et 34
avec Afzelia africana (Thoen, Bâ, 1989).
Parmi ces 322 champignons
cités, les deux tiers n’ont pas pu être
identifiés jusqu’au niveau de l’es-
pèce et 12 espèces nouvelles ont été
décrites. Il est cer tain que ces
récoltes sont encore incomplètes
mais elles illustrent déjà le manque
de données sur les macromycètes en
Afrique de l’Ouest et le besoin de
développer la mycologie dans cette
partie de l’Afrique.
La mention des champignons
dans le régime alimentaire des popu-
lations locales est une indication de
l’importance de cette ressource.
Toutefois, notre manque de connais-
sance des champignons ectomycorhi-
ziens ne permet pas d’identifier ces
Figure 4.
champignons et, de fait, réduit consi-
Cantharellus floridulus récolté sous Isoberlinia doka en Côte d’Ivoire. dérablement les possibilités de déve-
Cantharellus floridulus harvested under Isoberlinia doka in Côte d’Ivoire. lopper cette ressource.
Photo M. Ducousso. Parmi les espèces trouvées en
Afrique de l’Ouest, Cantharellus rufo-
punctatus, C. pseudofriesii, Lactarius
gymnocarpus, L. gymnocarpoides
Les espèces (Verbeken, 1997), Phlebopus sudani-
cus et Tubosaeta brunneosetosa sont
caractère charnu et dans son goût. de champignons mentionnées comme comestibles en
Une consistance plus ou moins vis- impliquées dans Afrique centrale et australe (Parent,
queuse peut être appréciée ici et Thoen, 1978 ; figure 3). La comestibi-
totalement rejetée ailleurs. Il en est
les symbioses lité de ces espèces doit encore être
de même pour le goût. Par exemple, ectomycorhiziennes confirmée. Toutefois, la liste des cham-
au nord de la Côte d’Ivoire, une rus- pignons ectomycorhiziens comestibles
sule amère est plus appréciée qu’une Les champignons n’est certainement pas restreinte à ces
espèce similaire plus douce. De associés à des espèces six espèces. D’après les données
même, en Indonésie, les scléro- locales ouest-afr icaines d’Afrique centrale et australe, il est très
dermes récoltés dans les plantations probable que d’autres espèces ecto-
d’Eucalyptus sont commercialisés, La diversité des champignons mycorhiziennes, notamment dans les
alors qu’en Afrique les espèces de ce ectomycorhiziens associés à des genres Amanita, Cantharellus et Rus-
genre ne sont pas connues pour être espèces locales est très importante. sula, sont comestibles. En dépit de la
consommées. La comestibilité d’une Les genres Russula, Lactarius, Ama- petite taille de ses fructifications qui
espèce de champignon est un carac- nita, Cantharellus, Inocybe et Boletus rend difficile sa cueillette en quantité
tère variable d’un lieu à un autre en sont parmi les plus représentés. Dans suffisante, Cantharellus floridulus
fonction des coutumes locales. En la partie méridionale du Burkina, 27 (figure 4) est consommé au Ghana.
conséquence, la comestibilité ne peut espèces ont été mentionnées (Sanon Des russules blanches sont également
être constatée qu’a posteriori. et al., 1997). En Côte d’Ivoire, consommées en Côte d’Ivoire.BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
59
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
Figure 5.
Pisolithus sp. sous Acacia holosericea.
Sénégal, région de Dakar. Figure 6.
Pisolithus sp. under Acacia holosericea. Suillus granulatus récolté sous Pinus kesiya. Fouta-Djalon, Guinée.
Senegal, Dakar region. Suillus granulatus harvested under Pinus kesiya. Fouta Djalon, Guinea.
Photo M. Ducousso. Photo M. Ducousso.
Les champignons folia a été trouvé en symbiose avec Le succès de l’introduction des
associés à des espèces Pisolithus sp. et Phlebopus sudani- pins en plantation a été possible
de planta tions cus au Sénégal (Ducousso, 1991). grâce à leur inoculation, comme cela a
En Corse (France), les jeunes été le cas en Guinée au début du
e
Les espèces d’arbres originaires fructifications de Pisolithus sp. sont xx siècle (Chevalier, 1947). Depuis
d’Australie comme les Eucalyptus, les consommées sous la dénomination lors, la litière des plantations âgées et
Acacia et les Casuarina n’ont jamais de « truffe de Corse ». Les fructifica- parfaitement bien établies est utilisée
fait l’objet d’une inoculation volon- tions plus matures sont utilisées pour pour l’inoculation des pépinières. De
taire par des champignons ectomyco- la teinture de la laine et des tissus. la sorte, la diversité des champignons
rhiziens dans notre zone d’étude. Aucun de ces usages n’est mentionné ectomycorhiziens est sensiblement
Leurs associations ectomycorhi- en Afrique. Les Phlebopus sudanicus plus importante que sous les espèces
ziennes éventuelles sont spontanées récoltés dans les plantations litto- australiennes, même si, comme pour
et totalement incontrôlées. Il a été rales du sud du Sénégal sont vendus ce dernier cas, l’introduction des
établi que les champignons ectomy- localement dans les restaurants tou- champignons n’a absolument pas été
corhiziens des forêts naturelles afri- ristiques. contrôlée. Ainsi, toutes sortes de
caines ne permettent pas de former A Madagascar, Cantharellus champignons ont été introduites
des ectomycorhizes réellement fonc- eucalyptorum ined. et des russules indistinctement, comprenant aussi
tionnelles avec les espèces ligneuses rouges récoltés dans les plantations bien des espèces toxiques, mortelles,
australiennes (Malajczuk et al., d’Eucalyptus robusta font l’objet d’un comestibles ou pathogènes. La pré-
1990 ; Martin et al., 1998). Les cham- commerce important (Ducousso, sence sous les pins de champignons
pignons ectomycorhiziens de ces 1999). Une espèce comestible de très toxiques comme l’amanite phal-
espèces auraient donc été introduits Xerocomus a également été trouvée loïde (Pegler, Piearce, 1980) incite à
fortuitement à partir de leur aire sous Acacia mangium (Ducousso, la plus grande prudence. Les popula-
d’origine en Australie. Leur diversité 1999). Les Eucalyptus et les Acacia tions locales le savent parfaitement et
est de ce fait très limitée. Les partagent des partenaires ectomyco- récoltent très rarement sous les pins,
Eucalyptus ont été trouvés en sym- rhiziens. Il est donc envisageable malgré l’abondance de champignons
biose avec Pisolithus sp., Sclero- d’utiliser les mêmes souches dans comestibles comme Suillus granula-
derma capense, S. verrucosum et des essais d’inoculation contrôlée. tus (Ducousso, 1991 ; figure 6). Cette
Phlebopus sudanicus. Pour les Les possibilités d’utiliser ces arbres espèce, vendue sous forme de conserve
Acacia, seul Pisolithus sp. est men- comme support pour la production en Europe et en Amérique du Nord,
tionné comme espèce ectomycorhi- contrôlée de champignons comes- est totalement négligée par les popu-
zienne (figure 5). Casuarina equiseti- tibles doivent être explorées. lations ouest-africaines. Il en est deBOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
60
FOCUS / ECTOMYCORRHIZAL FUNGI
Quelques cas
par ticulier s
Phlebopus sudanicus
Cette espèce ubiquiste, trouvée
aussi bien sous les espèces locales
qu’introduites (Thoen, Ducousso,
1989 b), est sujette à des interpréta-
tions contradictoires quant à sa comes-
tibilité : « Apprécié par les populations
indigènes » (Hariot, Patouillard,
1909) ; « Said to cause intoxication in
West Africa » (Pegler, Rayner, 1969 ;
Morris, 1990). Une partie de la
réponse réside probablement dans
l’ambiguïté de la description de cette
espèce. En effet, comme cela est pré-
senté en figure 7, de très importantes
variations morphologiques existent
entre trois spécimens récoltés au
Sénégal. Cela suggère la nécessité
d’une révision du genre Phlebopus
avant de se prononcer sur le caractère
comestible de ces espèces. Au
Sénégal, les Européens consomment
régulièrement et sans problème
Phlebopus sudanicus qui est très
appréciée. L’existence de variétés voire
d’espèces toxiques au sein du genre
Phlebopus n’est toutefois pas à écar-
ter. Apparemment, les populations
sénégalaises ne consomment pas
Phlebopus sudanicus et ne semblent
pas disposer d’informations sur sa
comestibilité et sa diversité.
Amanita bingensis
Figure 7.
Trois morphotypes différents de Phlebopus sudanicus récoltés au Sénégal. Cette amanite trouvée dans les
Three different morphotypes of Phlebopus sudanicus harvested in Senegal. forêts soudaniennes claires semi-déci-
Photos D. Thoen & M. Ducousso. dues fait également l’objet d’interpré-
tations contradictoires quant à sa
même pour Phallus (Dictyophora) Des essais d’inoculation contrô- comestibilité et il est clair qu’une pré-
indusiatus qui est commercialisée en lée sur Acacia mangium ont été mis cision univoque de la description des
Asie (Chang, Miles, 1989) et des en place en Afrique à des fins de amanites jaunes est indispensable. En
quatre autres espèces Amanita sp., recherche. Effectués sur de petites effet, d’après Morris, « a number of
Corditubera sp., Scleroderma cepa et surfaces, ils n’ont pas été orientés yellow Amanita are considered edible
Strobilomyces luteolus, trouvées sous vers la production de champignons in Central Africa » (Morris, 1990),
les pins en Guinée (Ducousso, 1991). comestibles. Toutefois, les potentiali- d’après Heim, « Amanita bingensis est
Dans le sud de la France, les tés des Eucalyptus, des Acacia aus- très toxique » (Heim, 1940), enfin
possibilités d’assurer la production traliens et des pins tropicaux comme d’après Dolacinski (communication
de Lactarius deliciosus en association support pour la production de cham- personnelle), Amanita bingensis est
avec des pins ont été explorées avec pignons comestibles méritent une mortelle. Les propos de Morris méri-
succès (Guerin-Laguette, 1998). Cet attention particulière, afin d’accroître tent d’être vérifiés et précisés avant de
exemple, développé en zone tempé- l’intérêt des populations pour ces statuer sur la comestibilité de cer-
rée, pourrait être adapté en Afrique. plantations en Afrique de l’Ouest. taines amanites jaunes.BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
61
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
Conclusion
Gyrodon inter medius
ined.
Ce champignon (figure 8) trouvé Les champignons ectomycorhi- champignons comestibles devraient
au Sénégal, exclusivement dans les ziens comestibles constituent une être entreprises en Afrique de
galeries forestières, est servi dans richesse potentielle en Afrique de l’Ouest, comme c’est le cas aux États-
quelques restaurants touristiques l’Ouest, où ils ne font pas l’objet d’un Unis depuis une quinzaine d’années.
comme un bolet local. Il est très commerce important, comme c’est le Les possibilités qu’une approche de
apprécié par les Européens et, avec cas en Afrique centrale, orientale et ce type permette de réduire, au moins
des collègues, nous avions pour habi- australe. Nos connaissances sur ces localement, la dégradation des forêts
tude, au début de la saison des champignons sont encore très insuffi- d’Afrique de l’Ouest sont à explorer.
pluies, de consommer de grandes santes. L’intervention de mycologues Les plantations peuvent être uti-
quantités de ce délicieux champi- pour caractériser la ressource est lisées comme support pour la pro-
gnon. À ce jour, il s’agit de la première indispensable, afin d’en assurer le duction de champignons comestibles.
mention du caractère comestible développement ultérieur. Les techniques d’inoculation et de
d’une espèce de Gyrodon en Afrique. La production de champignons suivi des souches sont déjà bien maî-
Aucune information sur la consom- ectomycorhiziens ne peut être assu- trisées dans les régions tempérées et
mation de cette espèce n’a été rele- rée qu’en forêt dans des sites très également, dans une certaine
vée auprès des populations sénéga- peu ou pas dégradés. Des recherches mesure, dans les régions tropicales
laises vivant à proximité de ces permettant d’allier une gestion fores- (Brunck et al., 1990). Bien sûr, de
galeries forestières. tière durable à la production de nouvelles souches adaptées aux
conditions écologiques de l’Afrique
de l’Ouest devront être sélection-
nées. Sous certaines conditions, l’in-
troduction de champignons sélection-
nés à l’origine de la plantation permet
de générer des revenus annuels qui
peuvent rapidement dépasser ceux
du bois, comme c’est le cas dans le
sud de la France, avec l’association
du chêne vert et de la truffe du
Périgord.
L’utilisation des champignons
ectomycorhiziens comestibles pour
promouvoir le développement
durable des forêts tropicales et des
plantations ne devrait plus être
négligé par les forestiers.
Figure 8.
Gyrodon intermedius ined., vendu comme bolet dans divers restaurants
touristiques sénégalais.
Gyrodon intermedius ined., served as a boletus in tourist restaurants in Senegal.
Photo M. Ducousso.BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
62
FOCUS / ECTOMYCORRHIZAL FUNGI
Références
bibliographiques
GUERIN-LAGUETTE A., 1998. Les lactaires PANDEY D., 1997. Hardwood plantations in
à lait rouge : mycorhization contrôlée des the tropics and subtropics. Tropical forest
BÂ A. M., SOUGOUFARA B., THOEN D., pins et caractérisation moléculaire. Appli- plantation areas 1995. Rome, Italie, Food
1987. The triple symbiosis of Casuarina cation à l’étude de la compétence écolo- and Agriculture Organisation of the United
equisetifolia in Senegal. In : Mycorrhizae gique et de la compétitivité d’isolats de Nations, 58 p.
in the next decade. Sylvia D. M., Hung L. L., Lactarius deliciosus. Thèse, École natio-
Graham J. H. (éd.). Gainsville, États-Unis, nale supérieure agronomique de Montpel- PARENT G., THOEN D., 1978. Food value of
University of Florida, p. 121. lier, France, 116 p. edible mushrooms from Upper-Shaba
region. Economic Botany, 31 (4) : 436-445.
BÂ A. M., BUYCK B., DESCHÈRES P., EYS- HARIOT P., PATOUILLARD N., 1909. Collec-
SARTIER G., IFONO F. G., DUCOUSSO M., tions recueillies au Congo français par M. PEGLER D. N., PIEARCE G. D., 1980. The
WEY J., GIRAUD E., FONTANA A., DIALLO M. A. Chevalier. Les champignons de la région edible mushrooms of Zambia. Kew Bull.,
A. K., DREYFUS B., 2000. Diversity and use Chari-Tchad. Bull. Mus. Hist. Nat. Paris, 35 : 475-491.
of ectomycorrhizal fungi in Guinea tropical 84-91. PEGLER D. N., RAYNER R. W., 1969. A
rain forest. In : Selection strategies for
HARKÖNEN M., SAARIMÄKI T., MWASUM- contribution to the Agaric flora of Kenya.
plant-beneficial microorganisms, Cost
BI L., 1995. Edible mushrooms of Tanzania. Kew Bull., 23 : 347-412.
830, 3-5 avril 2000, Nancy, France.
Karstenia, 35 (suppl.), 92 p. PIEARCE G. D., 1985. Livingstone and fungi
BRUNCK F., COLONNA J.-P., DOMMERGUES
HEIM R., 1936 a. Aperçu sur les champi- in tropical Africa. Bull. Brit. Mycol. Soc.,
Y.-R., DUCOUSSO M., GALIANA A., PRIN Y.,
gnons toxiques et comestibles des colo- 19 : 39-50.
ROEDERER Y., SOUGOUFARA B., 1990. La
maîtrise de l’inoculation des arbres avec nies françaises. Pathol. Exot. Vétérin. RAMMELOO J., WALLEYN R., 1993. The
leurs symbioses racinaires. Synthèse Comp., 3 : 1-31. edible fungi of Africa south of the Sahara :
d’une sélection d’essais au champ en zone a literature survey. National Botanic Gar-
HEIM R., 1936 b. Observations sur la flore
tropicale. Bois For. Trop., 223 : 24-42. den of Belgium, Scripta Botanica Belgica,
mycologique malgache. III. Trois bolets
gigantesques d’Afrique et de Madagascar. 5, 62 p.
BUYCK B., 1994. Ubwoba : les champi-
gnons comestibles de l’ouest du Burundi. Rev. Mycol. (Paris), 1 : 1-18. RIVIÈRE T., DIABATÉ M., DUCOUSSO M.,
Bruxelles, Belgique, Agcd, Publication PRIN Y., DREYFUS B., BUYCK B., EYSSAR-
HEIM R., 1940. Une amanite mortelle de
agricole n° 34, 123 p. TIER G., VERBEKEN A., DESCHÈRES P., BÂ
l’Afrique tropicale. Rev. Mycol. (Paris), 5 :
22-28. A. M., 2001. Diversity of ectomycorrhizal
CHANG S. T., MILES P. G., 1989. Edible
fungi associated with some native trees in
mushrooms and their cultivation. Boca
JORDAN P., WHEELER S., 1998. The ulti- Southern Guinea. Third International
Raton, Floride, États-Unis, Crc Press, 345 p.
mate mushroom book. Londres, Royaume- Conference on Mycorrhizas, 8-13 juillet
CHEVALIER A., 1947. Causes qui ont per- Uni, Anness Publishing Limited, 256 p. 2001, Adelaïde, Australie.
mis l’établissement et le maintien sans
LESUEUR D., DUCOUSSO M., 1995. Étude SANON K. B., BÂ A. M., DEXHEIMER J.,
soin d’une biocénose d’arbres exotiques
des associations symbiotiques dans les 1997. Mycorrhizal status of some fungi
au Jardin botanique de Dalaba. C. R. Acad.
essais agroforestiers réalisés dans les sta- fruiting beneath indigenous trees in Burki-
Sc. Paris, 224 : 1315-1318.
tions d’Oumé et de Korhogo en Côte na Faso. For. Ecol. Manag., 98 : 61-69.
DE KESEL A., YOROU S. N., SINSIN B., d’Ivoire. Rapport de mission Fed-Acp-Rpr
269, 23 p. THOEN D., BÂ A. M., 1989. Ectomycorrhi-
CODJIA J. T. C., 2001. Étude de la producti-
zae and putative ectomycorrhizal fungi of
vité des champignons comestibles de la
LOCQUIN M., 1954. Une chanterelle Afzelia africana Sm. and of Uapaca gui-
forêt classée de Wari-Maro (Bénin). La bio-
comestible de la Côte d’Ivoire : Hygropho- neensis Müll. Arg. in southern Senegal.
diversité botanique et l’expertise belge, 19
ropsis mangenotii sp. nov. J. Agric. Bot. New Phytol., 113 : 549-559.
et 20 octobre 2001, National Botanic Gar-
Trop. Appl., 1 : 359-361.
den of Belgium, Meise. THOEN D., DUCOUSSO M., 1989 a. Cham-
MALAJCZUK N., LAPEYRIE F., GARBAYE J., pignons et ectomycorhizes du Fouta-Dja-
DUCOUSSO M., 1991. Importance des sym-
1990. Infectivity of pine and eucalypt iso- lon. Bois For. Trop., 221 : 45-63.
bioses racinaires pour l’utilisation des
lates of Pisolithus tinctorius on roots of
acacias en Afrique de l’Ouest. Thèse, uni- THOEN D., DUCOUSSO M., 1989 b. Mycor-
Eucalyptus urophylla in vitro. New Phytol.,
versité de Lyon I. Nogent-sur-Marne, rhizal habit and sclerogenesis of Phlebo-
114 : 627-631.
France, Dakar, Sénégal, Cirad/Isra, 205 p. pus sudanicus (Gyrodontaceae) in Sene-
MARTIN F., DELARUELLE C., IVORY M., gal. Agric. Ecosys. Envir., 28 : 519-523.
DUCOUSSO M., 1999. Rapport d’une mis-
1998. Genetic variability in intergenic spa-
sion à Madagascar. Montpellier, France, THOEN D., ABOUROUH M., DUCOUSSO M.,
cers of ribosomal DNA in Pisolithus iso-
Cirad-forêt, 23 p. 2001. Ectomycorrhizal fungi associated
lates associates with pine, Eucalyptus and
Afzelia in lowland Kenyan forests. New with Eucalyptus in Africa and Madagascar.
DUCOUSSO M., LOUPPE D., OUATTARA N.,
Phytol., 139 : 341-352. Third International Conference on Mycorrhi-
EYSSARTIER G., BUYCK B., 1999. Des
zas, 8-13 juillet 2001, Adelaïde, Australie.
mycorhizes très diversifiées dans les
MORRIS B., 1990. An annotated check-list
jachères naturelles au nord de la Côte VERBEKEN A., 1997. Biodiversity of the
of the macrofungi of Malawi. Kirkia, 13 :
d’Ivoire. In : La jachère en Afrique tropi- Genus Lactarius Pers. in tropical Africa.
323-364.
cale, rôle, aménagements, alternatives, Thesis, University of Gent (Belgium).
13-16 avril 1999, Dakar, Sénégal, p. 126. NEWBERY D. M., ALEXANDER I. J., THO- Part 1 : text, 342 p. Part 2 : plates (269)
MAS D. W., GARTLAN J. S., 1988. Ectomy- and maps (29).
GÉRAULT A., THOEN D., 1992. Les champi-
corrhizal rain-forest legumes and soil
gnons dans les pharmacopées tradition-
phosphorus in Korup National Park, Came-
nelles de l’Afrique de l’Ouest. Rev. Méd.
roon. New Phytol., 109 : 433-455.
Pharm. Afr., 1 : 45-53.BOIS ET FORÊTS DES TROPIQUES, 2003, N° 275 (1)
63
CHAMPIGNONS ECTOMYCORHIZIENS / LE POINT SUR…
Synopsis
ECTOMYCORRHIZAL FUNGI
Forest types and tree species associated Ectomycorrhizal fungi
ASSOCIATED WITH NATIVE
with ectomycorrhizal fungi Many species belonging to the genera
AND PLANTED TREE SPECIES
West Africa has four types of forests dom- Russula, Lactarius, Amanita, Cantharellus,
IN WEST AFRICA:
inated by ectomycorrhizal trees: 1- dense Inocybe and Boletus (already over 200 sp.)
A POTENTIAL SOURCE
tropical rainforest, 2- Sudanese semi- are frequently associated with native
OF EDIBLE MUSHR OOMS
deciduous clear forest, 3- gallery forest, trees. Two thirds of sporophores collected
4- some plantations. until now have not been named with cer-
Marc DUCOUSSO
The first three types of forests are domi- tainty, and 12 new species have been
Amadou Moustapha BÂ
nated by three botanical families, Cae- recognised. It is also certain that these col-
Daniel THOEN
salpiniaceae, Dipterocarpaceae and lections are still incomplete, illustrating
Euphorbiaceae, all native to West Africa. the lack of data in the field of mycology in
Ectomycorrhizal fungi are
Within these families, two tribes of Cae- West Africa. Among species already found
natural compulsory symbiotic partners of
salpiniaceae: Amherstieae (10 sp.) and in West Africa, Cantharellus rufopunctatus,
some native and introduced tree species
Detarieae (3 sp.), one genus of Diptero- C. pseudofriesii, Lactarius gymnocarpus,
in West Africa. Several ectomycorrhizal
carpaceae: Monotes (1 sp.) and one genus L. gymnocarpoides, Phlebopus sudanicus
fungi are valuable edible mushrooms, like
of Euphorbiaceae: Uapaca (6 sp.) are and Tubosaeta brunneosetosa are men-
the Périgord black truffle or the Matsu-
known to be ectomycorrhizal. It is very tioned as edible species in Central and
take. However, the bulk of our data con-
likely that other species, especially Southern Africa. Other edible ectomycor-
cern fungi in temperate countries. The aim
belonging to genera Aphanocalyx, Didelo- rhizal species are expected to be found,
of this paper is to describe this resource in
tia, Microberlinia, Monopetalanthus and especially within the genera Amanita, Can-
West Africa.
Tetraberlinia (which have not yet been tharellus and Russula.
examined for ectomycorrhiza), are also Associated ectomycorrhizal flora in planta-
Current knowledge
ectomycorrhizal in West Africa. tions is very limited. Only ten species
A recent review on African edible fungi
In plantations, Pinus, Eucalyptus, Acacia (Amanita sp., Corditubera sp., Phallus
mentions 151 references. Only 3 of these
and Casuarina are known for their abilities (Dictyophora) indusiatus, Phlebopus
concern West Africa, the most recent hav-
to form ectomycorrhizas. These cover sudanicus, Pisolithus sp., Scleroderma
ing been published by Locquin in 1954.
about 100,000 ha in West Africa. Some capense, Sc. cepa, Sc. verrucosum Stro-
Edible mushrooms are better known in
other species have been recorded as ecto- bilomyces luteolus and Suillus granulatus)
Central and Eastern Africa than in West
mycorrhizal in West Africa but their plant- have been found. Some are commer-
Africa, where no recent work on edible
ed areas are negligible. cialised, like Suillus granulatus in Europe
mushrooms is available. Nevertheless,
and North America as yellow boletus, or
mushrooms are regularly cited in the diet
Edibility Phallus indusiatus in Asia as Zhu Sheng.
of local people, although there is no spe-
The two main qualities of an edible fungus Young sporophores of Pisolithus are eaten
cific data on species, origin and cooking
are fleshy consistency and tasty flavour. in Corsica (France) as “Corsican truffle”
methods.
Nevertheless, appreciation of the qualities and mature sporophores were used in
An approach based on recent data con-
of a fungus varies from one place to anoth- Europe as dye for wool and textiles. None
cerning edible mushrooms in Central,
er. A bitter taste or gummy consistency can of these uses are reported in West Africa.
Eastern and South Africa and on knowl-
be appreciated locally, but if the consis- Phlebopus sudanicus from coastal Casuar-
edge acquired over the last 20 years on
tency is too hard or too gummy, the fungi ina equisetifolia plantations in South
ectomycorrhizal fungi in West Africa has
are generally considered inedible, and Senegal is the only harvested species. It is
been developed to assess possibilities for
they rarely fruit. Possibilities for profitable served as a local boletus in some tourist
the production of edible ectomycorrhizal
harvests have to be considered in the light restaurants.
fungi in West Africa.
of edibility. Toxic and lethal fungi, includ- In Madagascar, Cantharellus eucalyptorum
ing closely related species, are rejected, ined. and some red Russula harvested
even if their toxicity has never been under Eucalyptus robusta are intensively
proven. In any case, edibility can only be traded. An edible, abundantly fruiting
confirmed after the event, which makes species of Xerocomus has also been found
local knowledge of prime importance. under Acacia mangium. The potential of
Eucalypts, Australian Acacia and tropical
pines to support the production of edible
mushrooms is worth exploring.
Edible ectomycorrhizal fungi are a poten-
tial resource for West Africa. They are
diversified and are not subject to intensive
trading. Their use in tropical forests and
plantations should be promoted.Vous pouvez aussi lire

















































