La périodicité des populations d'oiseaux au Québec : patrons et causes possibles - Mémoire Noémie Lacroix-Dubois Maîtrise en sciences forestières ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
La périodicité des populations d'oiseaux au Québec :
patrons et causes possibles
Mémoire
Noémie Lacroix-Dubois
Maîtrise en sciences forestières - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Noémie Lacroix-Dubois, 2022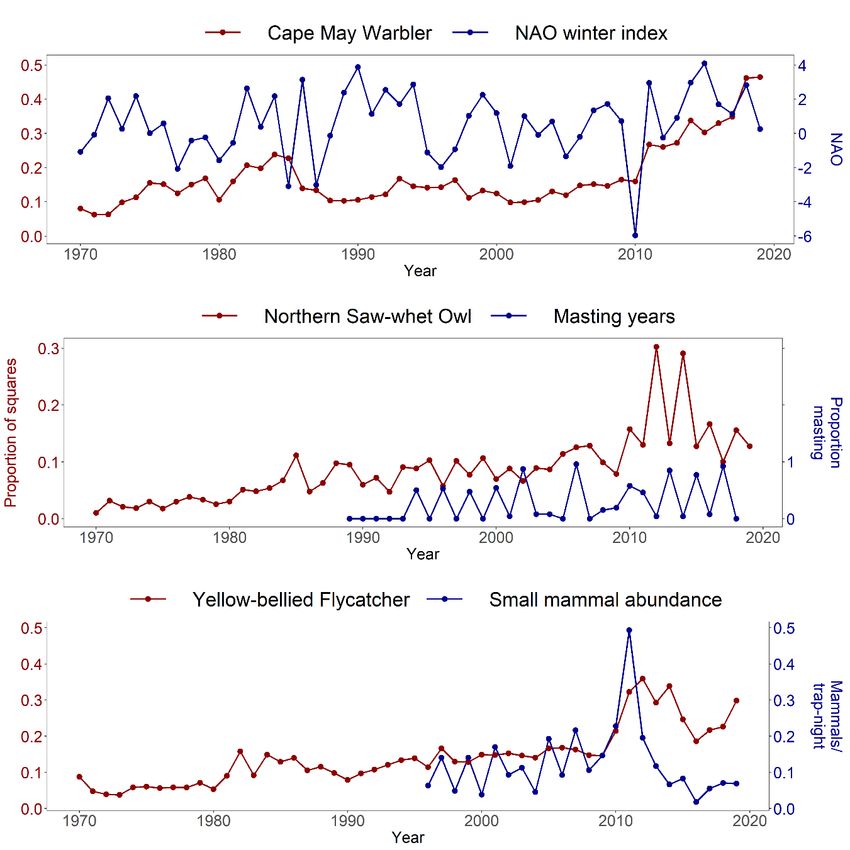
La périodicité des populations d’oiseaux au
Québec : patrons et causes possibles
Mémoire
Noémie Lacroix-Dubois
Sous la direction de :
André Desrochers, directeur de recherche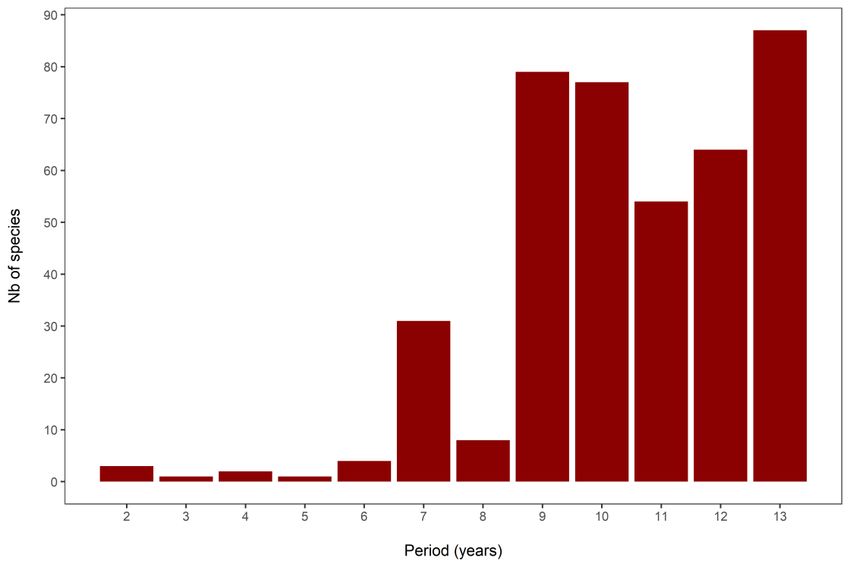
Résumé
Le dernier rapport sur l’État des oiseaux du Canada (2019), publié par un consortium
d’organismes gouvernementaux et d’OBNL, souligne le déclin de plusieurs espèces et la
hausse de plusieurs autres, depuis 50 ans. Plusieurs facteurs sont invoqués pour expliquer ces
tendances, mais leur importance demeure nébuleuse, faute de données expérimentales. À
défaut de données expérimentales, une connaissance détaillée des tendances demeure
pertinente à l’avancement de notre compréhension des causes de ces tendances. Un des
aspects les moins connus des tendances des oiseaux est leur périodicité. J’ai évalué la
périodicité des tendances des 250 espèces d’oiseaux les plus souvent observées dans la
province de Québec, lors des 50 dernières années (1970-2019). La métrique utilisée était la
proportion annuelle des carrés de 10 km x 10 km échantillonnés où l’espèce était observée.
Les données provenaient des programmes de science participative ÉPOQ et eBird. J’ai utilisé
des tests de randomisation et des régressions harmoniques pour évaluer des cycles allant de
2 à 13 ans. L’objectif secondaire de l’étude était d’explorer les causes possibles de la
périodicité. Pour ce faire, j’ai évalué l’association entre les patrons d’abondance annuelle et
l’Oscillation Nord-Atlantique, les années semencières du sapin baumier ainsi que
l’abondance des micromammifères. Parmi les 250 espèces étudiées, 149 montraient au moins
un cycle significatif entre 2 et 13 ans. Le cycle de 13 ans était le plus fréquent (87 espèces).
Les cycles de 138 espèces étaient corrélés avec l’Oscillation Nord Atlantique, ceux de 36
espèces avec les années semencières et ceux de 84 espèces avec l’abondance des
micromammifères. De nature exploratoire, cette étude pose les premiers jalons de notre
compréhension de la périodicité chez les populations d’oiseaux dans la province de Québec.
Cette étude permet d’énoncer des hypothèses plus précises, certaines desquelles sont
discutées en conclusion de ce mémoire
iiAbstract
The latest State of Canada’s Birds report (2019), published by a consortium of government
agencies and non-profit organizations, highlights the decline of several species and the
increase of several others over the past 50 years. Several factors may explain these trends,
but their importance remains unclear due to the lack of experimental data. In the absence of
experimental data, detailed knowledge of trends remains relevant to advancing our
understanding of the causes of these trends. One of the lesser-known aspects of bird trends
is their periodicity. I evaluated the periodicity of the trends of the 250 species of birds most
often observed in the province of Quebec, in the last 50 years (1970-2019). The metric used
was the annual proportion of the sampled 10 km x 10 km squares where the species was
observed. The data came from birdwatching programs ÉPOQ and eBird. I used harmonic
regressions to evaluate cycles ranging from 2 to 13 years. A secondary objective of the study
was to explore the possible causes of the periodicity. To do this, I evaluated the association
between annual abundance patterns and the North Atlantic Oscillation, balsam fir masting
years as well as the abundance of micro mammals. Among the 250 species studied, 149
showed at least one significant cycle between 2 and 13 years. The 13-year cycle was the most
frequent (87 species). The cycles of 138 species were correlated with the North Atlantic
Oscillation, those from 36 species with masting years, and those from 84 species with small
mammal abundance. This exploratory study lays the groundwork for our understanding of
periodicity in bird populations in the province of Quebec. This study makes it possible to
formulate more precise hypotheses, some of which are discussed at the conclusion of this
thesis.
iiiTable des matières
Résumé ................................................................................................................................................ ii
Abstract .............................................................................................................................................. iii
Table des matières .............................................................................................................................. iv
Liste des figures ................................................................................................................................. vi
Liste des tableaux .............................................................................................................................. vii
Liste des abréviations ....................................................................................................................... viii
Remerciements ................................................................................................................................... ix
Avant-propos ....................................................................................................................................... x
Introduction générale .......................................................................................................................... 1
Mise en contexte ............................................................................................................................. 2
Le phénomène de périodicité chez les oiseaux ........................................................................... 2
Les données sur la faune aviaire au Québec................................................................................ 6
Les tendances, sous plusieurs angles statistiques ........................................................................ 9
Description du problème de recherche .......................................................................................... 10
Objectifs de l’étude ....................................................................................................................... 10
Chapitre 1 – Periodicity in bird populations of Québec and possible causes .................................... 12
1.1 Résumé ........................................................................................................................................ 13
1.2 Abstract ....................................................................................................................................... 14
1.3 Introduction ................................................................................................................................. 15
1.4 Methods ....................................................................................................................................... 16
1.4.1 Statistical analysis ................................................................................................................ 18
1.5. Results ........................................................................................................................................ 19
1.5.1 General trends and periodicity ............................................................................................. 19
1.5.2 Correlates of periodicity ....................................................................................................... 23
1.6 Discussion ................................................................................................................................... 26
1.6.1 Possible causes of periodicity .............................................................................................. 26
1.7 Acknowledgments ....................................................................................................................... 30
Conclusion générale .......................................................................................................................... 31
Portée du projet ............................................................................................................................. 31
Périodicité et causes possibles....................................................................................................... 31
Approches et dérives statistiques .............................................................................................. 32
Comment utiliser ces résultats dans un contexte de conservation ............................................. 33
Perspectives d’avenir .................................................................................................................... 33
ivBibliographie ..................................................................................................................................... 35
Annexe 1 – Noms français, anglais et scientifiques des espèces analysées dans le cadre de l’étude.
Les espèces sont triées par ordre alphabétique. ................................................................................. 40
Annexe 2 – Espèces périodiques et statistiques associées à chaque période. Les espèces sont triées
par ordre alphabétique. ...................................................................................................................... 51
Annexe 3 – Espèces périodiques et statistiques associées à l’Oscillation Nord-Atlantique, les années
semencières et l’abondance des micromammifères. Les espèces sont triées par ordre alphabétique.
........................................................................................................................................................... 62
vListe des figures
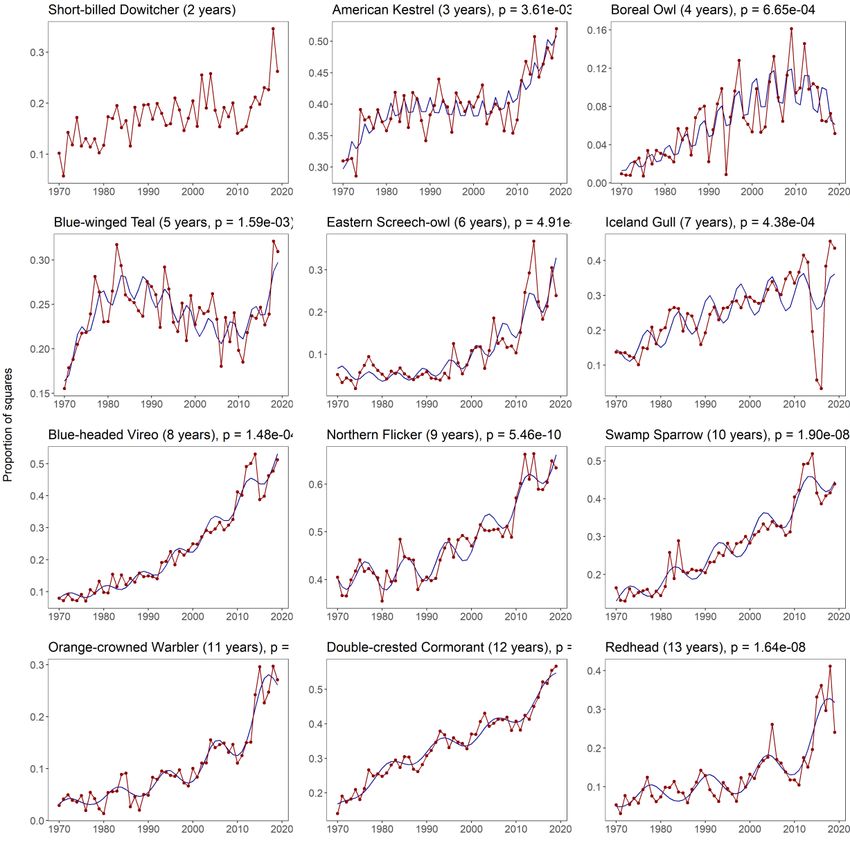
Figure 1.1 Number of species with a significant cycle with periods from 2 to 13 years. Species with
multiple significant cycles were counted multiple times. ................................................................. 21
Figure 1.2 Most pronounced cycles from 3 to 13 years. Red lines represent annual abundance, blue
lines represent fitted values from the model. Only the annual abundance is presented for the Short-
billed Dowitcher because it was tested by randomization rather than generalized linear modeling (see
explanation in the main text). ............................................................................................................ 22
Figure 1.3 Associations between the abundance of (1) Cape May Warbler and the North-Atlantic
Oscillation winter index, (2) Northern Saw-whet Owl and masting years and (3) Yellow-bellied
Flycatcher abundance and small mammal abundance. ..................................................................... 25
viListe des tableaux
Table 1.1 Number of species that exhibited significant periodicities for each period (columns, in
years) and for each family. Species sometimes had more than one significant periodicity. ............. 20
Table 1.2 Number of bird species that were positively and negatively associated with North-Atlantic
Oscillation, masting years and small mammal abundance for every family. .................................... 24
viiListe des abréviations
AMQ Atlas des Micromammifères du Québec
BBS Relevé des oiseaux nicheurs d’Amérique du Nord – Breeding Bird Survey
ENSO El Niño Oscillation Australe – El Niño Southern Oscillation
ÉPOQ Études de Populations d’oiseaux du Québec
GAM Modèle Additif Généralisé – Generalized Additive Model
GAMM Modèle Additif Généralisé Mixte – Generalized Additive Mixed Model
GLM Modèle Linéaire Généralisé – Generalized Linear Model
ICOAN Initiative de conservation des oiseaux de l’Amérique du Nord
MFFP Ministère des Forêts, de la Faune et des Parcs
NAO North Atlantic Oscillation
NCAR National Center for Atmospheric Research
ONA Oscillation Nord-Atlantique
RH Régression Harmonique
OBNL Organisme à but non lucratif
viiiRemerciements
J’adresse mes remerciements à ceux qui ont contribué à la réalisation de mon mémoire.
Premièrement, j’aimerais remercier QuébecOiseaux pour avoir financé cette étude par
l’entremise de la bourse du Fonds Atlas, ainsi que l’Observatoire d’oiseaux de Tadoussac
pour m’avoir octroyé généreusement une bourse. J’aimerais ensuite remercier André
Desrochers, mon directeur de recherche, pour son soutien tout au long de ma maîtrise, pour
tout le temps qu’il m’a consacré et pour ses conseils qui m’ont guidé dans mon travail. Merci
aussi à Junior Tremblay et à Pascal Côté, qui ont répondu à mes interrogations lorsque j’en
avais et pour m’avoir apporté des précisions dans leur domaine d’expertise. J’aimerais aussi
remercier Mathieu Bouchard ainsi qu’Alexandre Anctil de m’avoir fourni des données
essentielles à la réalisation de mon mémoire. Finalement, j’aimerais remercier Jacques
Larivée, qui a posé les premiers jalons de l’ornithologie comme science participative au
Québec avec le programme ÉPOQ.
ixAvant-propos
Ce projet d’étude a été financé par la bourse du Fonds Atlas de QuébecOiseaux et par
l’Observatoire d’Oiseaux de Tadoussac. Je certifie être l’auteure principale de ce document.
André Desrochers, mon directeur de recherche, a proposé les grandes lignes de l’analyse
statistique, à partir desquelles j’ai réalisé l’analyse finale. J’ai également réalisé la rédaction
ce me mémoire. Ce mémoire contient un article qui sera une pré-publication (preprint) dans
BioRxiv en 2021, et explique la raison pour laquelle le chapitre principal est rédigé en
anglais. André Desrochers m’a aidé à élaborer ce projet et m’a conseillé lors de la réalisation
de chacune des étapes. Pascal Côté m’a également apporté son aide dans la rédaction et m’a
donné de nombreux conseils. Par conséquent ils seront coauteurs de l’article.
xIntroduction générale
Dans un article récent, Rosenberg et al. (2019) rapportent une perte nette de 2,9 milliards
d’oiseaux en Amérique du Nord depuis les années 1970. Selon ces auteurs, le biome des
prairies ainsi que celui de la forêt boréale seraient ceux ayant les plus grandes pertes nettes
d’individus. Par exemple, ils estiment que 50 % des espèces présentes en forêt boréale sont
en déclin, ce qui représenterait une perte nette de 500 millions d’individus. L’initiative de
conservation des oiseaux de l’Amérique du Nord (ICOAN) présente un portrait plus nuancé
dans son dernier rapport sur l’état des populations d’oiseaux au Canada. En effet, l’organisme
rapporte des groupes d’espèces comme la sauvagine, qui ont augmenté et d’autres groupes,
comme les insectivores aériens, qui ont diminué. En ce qui concerne les oiseaux forestiers,
l’ICOAN rapporte 49 espèces en augmentation, 30 espèces stables et 44 espèces en
diminution (ICOAN 2019).
On utilise souvent les tendances des populations aviaires à titre d’indicateurs de l’état de
« santé » du milieu. Par exemple, une dégradation de l’habitat peut causer une baisse du
succès de reproduction, une hausse de la prédation des nids, une baisse du succès
d’appariement et une réduction du succès de nidification, ce qui entraine le déclin des
populations d’oiseaux (Robinson et al. 1995; Austen et al. 2001). Un tel déclin peut
également être associé à la présence d’espèces exotiques envahissantes (Gurevitch et Padilla
2004). Cependant, la diminution des populations d’oiseaux peut également être sans lien avec
la « santé » de l’écosystème, et être dû à l’action de phénomènes naturels, tels que le climat,
le parasitisme, la prédation et d’autres relations trophiques (Savard et al. 2015; Ottersen et
al. 2001; Nott et al. 2002; Morrissette et al. 2010). Malgré tout, une diminution des
populations d’oiseaux, qu’elle soit de nature anthropique ou naturelle, pourrait entrainer des
effets négatifs dans l’environnement en raison des services écosystémiques que ces espèces
rendent (Inger et al. 2015). La meilleure manière de déterminer quelles sont les causes du
déclin chez une espèce serait par expérimentation, ce qui permettrait de contrôler les effets
confondants. Cependant, il est impossible d’effectuer des expériences contrôlées à l’échelle
de populations entières puisqu’on peut difficilement manipuler les paramètres
environnementaux, encore moins répliquer de telles manipulations (Morrissette et al. 2010).
1À défaut d’expérimenter, une manière d’améliorer notre compréhension est d’analyser plus
en détails les tendances. L’étude de l’abondance chez les populations d’oiseaux est donc
essentielle dans la compréhension des différents écosystèmes, tout comme l’examen des
causes derrière les fluctuations de populations (Morrissette et al. 2010).
Mise en contexte
Le phénomène de périodicité chez les oiseaux
Au-delà des tendances, certaines espèces fauniques présentent une périodicité1, c’est-à-dire
que leur abondance fluctue à des intervalles plus réguliers que ceux auxquels on s’attendrait
par hasard (Moss et Watson 2001). Le phénomène de périodicité est connu chez plusieurs
taxons. Les petits mammifères sont reconnus pour leurs cycles de populations qui ont été
documentés dans plusieurs régions, particulièrement dans la région de Fennoscandie, avec
des cycles de populations variants de 3 à 7 ans (Hörnfeldt 1978; Cheveau et al. 2004).
Ailleurs en Europe et en Amérique du Nord, les fluctuations étaient plutôt reconnues comme
étant irrégulières, ou ayant des amplitudes plus faibles (Hanski et al. 1991). Cependant,
Cheveau et al. (2004) ont démontré que des cycles de 4 ans étaient présents dans la forêt
boréale de l’Est de l’Amérique du Nord chez certaines espèces de petits mammifères, ce qui
est similaire aux résultats observés en Fennoscandie. Des cycles de populations sont
également présents chez les Lépidoptères. En effet, certaines espèces connaissent des cycles
variant entre 8 et 11 ans, et les populations peuvent être synchrones sur de grandes échelles
géographiques (Myers 1998; Myers et Cory 2013). De plus, les cycles des Lepidoptères sont
plus apparents dans des environnements plus rigoureux (Myers 2018). Cependant, un des
exemples les plus connus en biologie demeure celui du lièvre et du lynx. En effet, les
populations de ces deux espèces montrent des cycles d’environ 10 ans dans les forêts boréales
d’Amérique du Nord (Keith et Winberg 1978; Yan et al. 2013). Dans l’exemple du lynx et
du lièvre, les fluctuations sont principalement causées par un facteur dépendant de la densité,
1
Dans ce mémoire, le terme « périodicité » est utilisé pour décrire le phénomène général par lequel l’abondance
fluctue selon des intervalles plus ou moins réguliers tandis que le terme « cycle » est utilisé pour décrire la
succession relative d’une période donnée (ex : cycle de 2 ans).
2soit le taux de prédation par le lynx. Dans d’autres systèmes, les principaux facteurs
dépendants de la densité peuvent être la compétition ou le parasitisme. Ces facteurs régulent
les populations en provoquant une diminution de population lorsque les effectifs sont élevés
et le contraire lorsque les effectifs sont bas (Newton 2007). Prenons l’exemple de la
compétition pour la nourriture : si la nourriture est abondante, la population augmente. Cette
augmentation de la population provoque une diminution de la disponibilité de la nourriture
per capita, et donc une diminution de la fécondité ou de la survie des individus ; la population
diminue. Une fois les effectifs réduits, la disponibilité des ressources augmente, et ainsi de
suite (Williams et al. 1993). Un autre exemple de facteur dépendant de la densité pouvant
causer des cycles de populations est la saisonnalité. En effet, une étude de Betini et al. (2017)
a démontré que certains traits chez les animaux, tel que la taille par exemple, permettait de
produire une progéniture plus importante durant la saison de reproduction et diminuait les
chances de survie lorsque les ressources sont plus limitées. Ainsi, les individus de plus grande
taille sont plus abondants lorsque les effectifs de population sont plus faibles et qu’il y a
suffisamment de ressources. Inversement, les individus de plus petite taille sont plus
abondants lorsque les effectifs de population sont plus élevés. Ces changements causés par
la saisonnalité pourraient ainsi provoquer des cycles de populations prévisibles, comme
Betini et al. (2017) l’ont démontré avec des populations de Drosophila melanogaster.
Des phénomènes périodiques indépendants de la densité peuvent aussi avoir un effet sur les
populations fauniques, notamment le climat et les relations trophiques indirectes. Ces
phénomènes affectent donc les populations, sans aucun lien avec le nombre d’individus.
Cependant, bien que ces mécanismes puissent avoir un impact important sur les populations
à l’échelle locale, ils ne provoquent pas nécessairement de variation au niveau de la
population. En effet, si ces facteurs ont une influence localement mais ne sont pas synchrones
spatialement, ces facteurs ne vont pas affecter la dynamique de la population à grande échelle
(Morrissette et al. 2010).
Bien que le phénomène de périodicité soit connu chez plusieurs taxons, il demeure très peu
étudié chez les populations d’oiseaux, et peu d’espèces sont reconnues comme montrant des
cycles de populations (Cheveau et al. 2004; Koenig 2001; Moss et Watson 2001; Shaw et al.
2004). Ceci est particulièrement vrai pour les fluctuations à très long terme, dont la mise en
3évidence nécessite des estimations d’abondance remontant à plusieurs décennies, voire des
siècles, indisponibles excepté quelques cas comme ceux provenant de l’accumulation de
guano (Duffy 1983) et de marqueurs isotopiques (Hargan et al. 2019).
Plusieurs facteurs peuvent être à l’origine de la périodicité chez les populations d’oiseaux.
Le reste de cette introduction présentera la perspective particulière du Québec. L’Oscillation
Nord-Atlantique (ONA), les années semencières ainsi que l’abondance de
micromammifères sont des phénomènes naturels présents dans la province de Québec qui
peuvent montrer une périodicité et qui sont reconnus comme ayant une influence sur les
populations d’oiseaux. Il existe plusieurs phénomènes climatiques et atmosphériques
associées à la variabilité du climat globale à court terme, tels que El Niño (ENSO – El Niño
Southern Oscillation) et l’ONA (Nott et al. 2002). Ces phénomènes peuvent avoir un impact
sur les écosystèmes marins, terrestres et limniques, en plus d’influencer plusieurs processus
écologiques et modifier les traits phénotypiques chez plusieurs espèces (Stenseth et al. 2003;
Hurrell et Deser 2009; Hallett et al. 2004). Bien que ENSO exerce son influence de façon
globale, l’ONA représente le phénomène climatique dominant dans la région Nord
Atlantique (Stenseth et al. 2003). En effet, l’ONA est un phénomène océanique et
atmosphérique qui affecte la force et la direction des vents, ce qui provoque des changements
dans les températures et les précipitations dans l’hémisphère Nord, incluant les différents
écosystèmes du Québec (Savard et al. 2015). Bien que l’ONA ne possède pas de cycle
dominant, plus de 75% de la variance de cet indice se produit sur une échelle de temps
inférieure à une décennie. Plusieurs indices ont été définis dans le passé afin de caractériser
ce phénomène, mais l’indice régulier hivernal est celui qui semble avoir le plus d’influence
sur les écosystèmes (Ottersen et al. 2001). Dans la forêt boréale de l’Est de l’Amérique du
Nord, un indice hivernal positif signifie un hiver plus froid avec peu de précipitations tandis
qu’un indice négatif indique un hiver doux et des précipitations importantes (Savard et al.
2015). Par exemple, cela peut entrainer des variations dans la productivité primaire et la
quantité d’insectes disponibles, ce qui affecte ensuite les populations d’oiseaux forestiers et
leur succès reproducteur (Nott et al. 2002). Le taux de survie des jeunes, la croissance des
individus, la densité des populations ainsi que la phénologie de reproduction peuvent
également être affectés par l’ONA (Ottersen et al. 2001). Au Québec, l’abondance de
4l’Épervier brun2 était positivement corrélée avec l’indice de l’ONA de l’hiver précédent entre
1996 et 2010, ce qui suggère l’influence du climat sur la dynamique des écosystèmes de la
province (Savard et al. 2015). Cependant, le lien entre les conditions climatiques locales et
l’indice hivernal n’est pas toujours évident. En effet, l’utilisation d’un tel indice comporte
des désavantages, tels que la variation spatiale, la saisonnalité, la non-stationnarité, la non-
linéarité et le manque de corrélation. Il est donc important d’avoir une bonne connaissance
de ce phénomène climatologique et d’utiliser l’indice approprié au système écologique à
l’étude (Stenseth et al. 2003).
Les années semencières sont un autre phénomène qui affecte l’abondance des populations
d’oiseaux. Ce sont les années où la production de graines et de cônes est plus importante chez
les populations de conifères et de plantes vivaces. Cette production est intermittente puisque
les coûts énergétiques engendrés par une telle production une année mènent à une production
réduite l’année suivante. En forêt boréale, des cycles de 2 à 3 ans sont généralement observés
chez les conifères (Strong et al. 2015). Ces fluctuations de production ont une influence
directe sur les espèces aviaires qui dépendent de ces ressources. En effet, les années de forte
production permettent aux individus de se reproduire sur une plus longue période l’année
suivante et de produire davantage de couvées, ce qui mène à une augmentation du nombre
de jeunes. On observe ensuite une forte diminution de la reproduction les années
subséquentes (Strong et al. 2015). Cette réduction s’explique par le fait que de hautes densités
de populations résultant de fortes productions de graines sont ensuite confrontées à une faible
quantité de nourriture l’automne suivant (Koenig et Knops 2001). Les années semencières
peuvent également causer le déplacement d’individus, aussi appelé « migration éruptive »
(Dunn 2019). En effet, la production de graines peut être synchronisée sur plus de 1 000 km,
ce qui force certaines espèces d’oiseaux en quête de cette ressource à se déplacer sur des
distances de cet ordre de grandeur (Dunn 2019). L’abondance de plusieurs espèces d’oiseaux
forestiers est synchrone avec les années semencières. Les irruptions de populations de la
Mésange à tête noire en Ontario étaient synchrones avec la production de cônes entre 1951
et 1968 (Bock et Lepthien 1976). Une étude de Koenig (2001) a également démontré que
2
L’utilisation des majuscules dans ce document est conforme au document technique suivant : Chabot J, David
N, 1986. « La majuscule dans la nomenclature biologique ». Can J Zool 64 :2072-2073
5plusieurs espèces de l’est de l’Amérique du Nord montraient une périodicité correspondant
au patron de production de graines en forêt boréale. Parmi ces espèces, on retrouve la Sittelle
à poitrine rousse, le Sizerin flammé ainsi que le Tarin des pins.
L’abondance des micromammifères peut aussi affecter l’abondance de certaines espèces
d’oiseaux. Les micromammifères sont reconnus pour montrer des cycles qui varient de 3 à 7
ans dans plusieurs pays de l’hémisphère Nord (Cheveau et al. 2004). Ainsi, les oiseaux
prédateurs qui se nourrissent de ce mammifère peuvent être affectées par les fluctuations
d’abondance de leurs proies. C’est le cas de la Nyctale de Tengmalm, dont l’abondance était
fortement corrélée avec celle du Campagnol à dos roux entre 1990 et 2000 au Québec. En
effet, l’abondance de la Nyctale de Tengmalm montrait des cycles de populations de 4 ans
qui correspondaient avec celui de sa proie (Cheveau et al. 2004). L’abondance du Grand-duc
d’Amérique était également corrélée avec celle du Lièvre d’Amérique au Yukon (Boutin et
al. 1995).
Les données sur la faune aviaire au Québec
Les données ornithologiques du Québec se regroupent en deux grandes catégories, soit les
relevés standardisés et les programmes de science participative, aussi appelée science
citoyenne. Parmi les relevés standardisés, on retrouve notamment le Relevé des oiseaux
nicheurs d’Amérique du Nord (BBS – Breeding Bird Survey). Mis en place en 1966, le BBS
constituait jusqu’à récemment la source principale de donnés sur les espèces d’oiseaux
nichant en Amérique du Nord (Gouvernement du Canada 2018). En effet, une grande partie
des oiseaux chanteurs qui se reproduisent en Amérique du Nord est suivie par ce programme,
au moyen de relevés de bord de routes effectués durant la saison de reproduction. Cependant,
plusieurs espèces sont sous représentées par les relevés du BBS. En effet, les espèces qui
nichent de façon clairsemée ou localement ne sont pas couvertes par un nombre suffisant de
routes, comme c’est le cas pour plusieurs rapaces et espèces coloniales. Certains oiseaux
nichent également dans des régions éloignées où le réseau routier est moins développé,
notamment dans le nord de la forêt boréale (Dunn et al. 1996). De plus, ce programme de
suivi repose sur l’aide de bénévoles, qui sont moins nombreux en forêt boréale que dans la
vallée du Saint-Laurent (Savard et Ibarzabal 2001). Enfin, les tendances obtenues à partir des
6relevés du BBS sont susceptibles d’être biaisées en raison de la sélection des sites par les
bénévoles. Ainsi, des déclins de populations peuvent être observés dans les routes BBS,
même s’il n’y a pas de déclin généralisé (Fournier et al. 2019). Les espèces nocturnes, les
espèces rares, les oiseaux de rivages, les oiseaux aquatiques, les oiseaux de milieux humides
ainsi que les espèces qui se reproduisent dans les milieux arctiques sont également sous
représentées (Walker et Taylor 2020). C’est pourquoi des méthodes alternatives s’avèrent
attrayantes pour effectuer le suivi des oiseaux dans la forêt boréale du Québec.
Les programmes de science participative constituent une solution de plus en plus intéressante
dans l’étude des tendances de populations d’oiseaux. On retrouve plusieurs programmes de
ce genre à travers le monde. BirdTrack est un programme qui vise à suivre la migration des
espèces d’oiseaux en Grande-Bretagne et en Irlande. Cette base de données provient
d’observations récoltées par des citoyens et des ornithologues de tous les niveaux. DOF-
basen au Danemark ainsi que Artportalen en Suisse sont des programmes similaires (Baillie
et al. 2006). Le Québec est un pionnier mondial en matière de science ornithologique
participative. Dès les années 1940, des ornithologues québécois prirent l’habitude de noter
systématiquement leurs observations; Victor Gaboriault fut une réelle inspiration en cette
matière (David 1978). Avec la popularisation des ordinateurs, ces observations allaient être
saisies numériquement et gérées avec le programme ÉPOQ (Études des Populations
d’oiseaux du Québec), mis sur pied en 1975 par un professeur du Cégep de Rimouski,
Jacques Larivée (Droege et al. 1998). ÉPOQ fonctionnait avec les technologies de l’époque
(listes en papier, courrier postal, etc.) sensiblement de la même façon que le programme eBird
(Dunn et al. 1996). À sa fermeture, ÉPOQ contenait 10 025 915 mentions d’espèces
d’oiseaux (Desrochers 2019).
Désormais, les ornithologues québécois utilisent eBird, qui a progressivement remplacé
ÉPOQ et incorporé ses données. Le programme eBird a été mis sur pied par le Cornell Lab
of Ornithology et la National Audubon Society en 2002 (Sullivan et al. 2009). Ce programme
regroupe les observations d’ornithologues amateurs et professionnels au moyen de listes
d’observations. Ainsi, chaque personne observant un ou plusieurs oiseaux est en mesure de
collecter ces données et les rendre disponibles au public (Sullivan et al. 2009). En 2017, il y
avait plus de 18,8 millions de listes compilées (Walker et Taylor 2017). Au moment d’écrire
7ces lignes, eBird comptait 53,8 millions de listes, provenant de toutes les régions du globe,
dont 1,6 millions provenant du Québec. En raison de la quantité importante de données qu’il
contient, le programme eBird constitue un outil pouvant servir à la communauté scientifique,
que ce soit pour examiner la distribution de certaines espèces, l’abondance relative ou bien
pour analyser des tendances (Sullivan et al. 2009).
L’utilisation de listes d’observations provenant des programmes de science participative
montre plusieurs avantages. En effet, lorsqu’elles sont standardisées et récoltées en quantité
suffisante, les listes d’observations permettent d’analyser la phénologie, les tendances des
populations, la structure des communautés ainsi que la richesse en espèces (Droege et al.
1998). De plus, ces programmes permettent de couvrir des vastes territoires où certaines
données sont manquantes et des saisons durant lesquelles les relevés traditionnels n’offrent
pas de donnés. Par définition, la science participative est également économique puisqu’elle
ne requiert presque pas de personnel rémunéré. Finalement, ces programmes couvrent des
séries temporelles qui ne serait pas possible d’avoir avec des relevés standardisés d’un point
de vue économique et logistique (Dunn et al. 1996; Walker et Taylor 2017). Toutefois, les
données provenant de la science participative comprennent plusieurs sources de variabilité.
Parmi celles-ci, on retrouve l’effort d’échantillonnage par observateur, la couverture
géographique très inégale ains que la capacité de détection et la compétence des participants
(Walker et Taylor 2017). De plus, comme ces données sont récoltées de façon opportuniste,
à n’importe quel moment et à n’importe quel endroit, ces dernières comportent du bruit, c’est
à dire de l’information sans intérêt. De plus, les observations ne sont pas réparties également
dans le temps puisque les ornithologues se concentrent sur les périodes de forte diversité en
espèces, particulièrement lors des migrations (Dunn et al. 1996).
Heureusement, plusieurs approches statistiques permettent de prendre en compte ces biais et
ces variabilités (Walker et Taylor 2017). Dans une étude récente, Walker et Taylor (2020)
ont comparé les tendances obtenues avec les données eBird avec celles obtenus à partir des
données du BBS. Les résultats démontraient que les tendances calculées à partir des indices
annuels provenant d’eBird étaient fortement corrélés avec celles obtenues à partir des
données du BBS, ce qui démontre l’utilité du programme eBird pour l’étude des tendances
des populations aviaires.
8Les tendances, sous plusieurs angles statistiques
Les biostatisticiens tendent actuellement à mesurer les tendances avec des modèles linéaires
généralisés (GLM) ainsi que des modèles additifs généralisés (GAM). Les GLM et les GAM
ont comme avantage de tenir compte de structures de données distribuées de façon non
Gaussienne (non-normales). Ils sont donc utiles pour développer des modèles écologiques
qui représentent mieux les données sous-jacentes, ce qui augmente notre compréhension des
systèmes écologiques (Guisan et al. 2002). Plusieurs autres méthodes ont été développées
pour analyser les tendances. Premièrement, on peut mentionner l’utilisation de GAMMs
(Modèles linéaires additifs mixtes – Generalized Additive Mixed Models) afin de modéliser
des tendances non linéaires. Cette technique permet de séparer les fluctuations à court terme
des changements à long terme, ce qui réduit la détection erronée des changements de
population à long terme comparativement à un GAM (Knape 2016). La méthode « List-
Length only » est une autre méthode qui utilise le nombre d’espèces observées par listes
d’observation comme approximation de l’effort, appliqué à un modèle logistique. Comme
cette méthode requiert peu d’informations, elle peut être appliquée à plusieurs sources de
données différentes, ce qui représente un avantage (Horns et al. 2018). Les régressions
linaires Gaussiennes ont également été utilisées dans le passé afin d’estimer les tendances de
populations d’oiseaux. Cependant, une telle méthode peut avoir des limites conceptuelles et
pratiques importantes, ce qui peut mener à des estimations biaisées des tendances de
populations (Sauer et al. 2004). Bien que plusieurs de ces approches permettent de tester la
périodicité de façon explicite, nous n’en connaissons pas d’exemples dans la littérature
scientifique. Par exemple, les GAM sont plus souvent utilisés de manière phénoménologique,
c’est-à-dire dans une optique de curve-fitting, c’est-à-dire la production de paramètres
hautement flexibles sans fondement conceptuel simple. Afin d’évaluer la périodicité des
tendances de manière explicite, il est plus simple et pertinent de la formuler avec des
fonctions trigonométriques (sinus et cosinus). De tels modèles se regroupent sous le vocable
de régressions harmoniques (RH; Lück et al. 2014). L’utilisation des RH est avantageuse
puisqu’elles représentent un outil relativement simple qui permet de calculer la phase et
l’amplitude pour une période ou fréquence donnée.
9Description du problème de recherche
Bien qu’on connaisse les tendances des populations d’oiseaux présents au Québec, les
analyses réalisées jusqu’à maintenant ne permettent pas d’évaluer le phénomène de
périodicité chez les populations d’oiseaux. Pourtant, la connaissance de ce phénomène
pourrait être un outil important dans la gestion des espèces montrant une périodicité. En effet,
l’étude de la périodicité nous permet de mieux comprendre la dynamique des populations.
De plus, l’influence de l’ONA, des années semencières ainsi que l’abondance des
micromammifères est connu pour peu d’espèces. Il est donc difficile de faire des constats
généraux. En effet, l’influence de ces phénomènes sur l’ensemble des espèces d’oiseaux
présentes au Québec n’a pas fait l’objet de recherches ou d’études à ma connaissance. De
plus, nous ne savons pas s’il y a une interaction entre les différents phénomènes. Finalement,
les données eBird ont été utilisées dans plusieurs études, mais rarement dans la modélisation
de tendances à long terme (Walker et Taylor 2017; Walker et Taylor 2020).
Les ornithologues du Québec ont généré une quantité de données possiblement inégalée, ce
qui nous permet de faire de telles analyses. En effet, le Québec est une des rares régions du
globe avec des données couvrant l’entièreté de l’année sur plus de 50 ans. L’ONA, les années
semencières ainsi que l’abondance des micromammifères sont déjà reconnus comme ayant
une influence sur certaines espèces d’oiseaux. Ainsi, il est possible de faire cette analyse pour
l’ensemble des espèces afin d’avoir une vision globale de l’impact qu’ont ces phénomènes
sur la dynamique des populations.
Objectifs de l’étude
L’objectif principal de cette étude est de décrire et d’amorcer une réflexion sur la périodicité
chez les populations d’oiseaux du Québec. De nature exploratoire, cette étude permettra de
poser des hypothèses plus précises quant à l’évolution récente des populations du Québec.
Cette étude a également plusieurs objectifs secondaires. Le premier objectif secondaire
consiste à généraliser l’application de modèles de régressions harmoniques à des
distributions non-Gaussiennes. Le second est d’explorer les causes possibles pouvant
expliquer les patrons de périodicité chez les espèces d’oiseaux, notamment l’Oscillation
10Nord-Atlantique, les années semencières ainsi que l’abondance des micromammifères.
Finalement, cette étude mettra en évidence la pertinence des programmes de science
participative dans la compréhension des tendances de la faune aviaire.
11Chapitre 1 – Periodicity in bird populations of
Québec and possible causes
121.1 Résumé
La périodicité est un phénomène bien connu chez plusieurs taxons, mais demeure peu étudié
chez les oiseaux. Cette étude vise à développer une meilleure connaissance des tendances des
populations d’oiseaux au Québec, plus particulièrement sur le phénomène de périodicité.
Nous avons utilisé les données provenant de deux sources de science participative, ÉPOQ et
eBird, afin d’évaluer des cycles allant de 2 à 13 ans pour les 250 espèces les plus
fréquemment observées dans la province. Pour ce faire, nous avons utilisé des modèles
linéaires harmoniques. Nous avons testé si l’Oscillation Nord Atlantique, les années
semencières et l’abondance des micromammifères sont associées avec la périodicité. Parmi
les 250 espèces étudiées, 149 montraient au moins un cycle significatif entre 2 et 13 ans. Le
cycle de 13 ans était le plus fréquent, avec 87 espèces. Parmi les espèces périodiques, 138
étaient corrélées avec l’Oscillation Nord Atlantique, 36 avec les années semencières et 84
avec l’abondance des micromammifères. Les résultats obtenus montrent que le phénomène
de périodicité est bien présent chez plusieurs espèces d’oiseaux au Québec et que certains
phénomènes régulent fortement les effectifs de ces mêmes espèces. Cependant, cette étude
ne pose que les premiers jalons de notre compréhension de la périodicité chez les populations
d’oiseaux au Québec. Nous souhaitons que des études ultérieures puissent se baser sur la
présente étude afin d’approfondir nos connaissances de la périodicité des populations
d’oiseaux.
131.2 Abstract
Periodicity is a well-known phenomenon in several taxa but remains little studied in birds.
This study aims to deepen our understanding of bird population trends in Quebec, with a
focus on periodicity. We used data from ÉPOQ and eBird to determine whether cycles
ranging from 2 to 13 years exist in the 250 most frequently observed species in the province.
To do this, we used linear harmonic models. We tested whether the North Atlantic
Oscillation, masting years and small mammal abundance are associated to periodicity.
Among the 250 species studied, 149 showed at least one significant cycle between 2 and 13
years. The 13-year cycle was the most frequent, with 87 species. Of the periodic species, 138
were correlated with the North Atlantic Oscillation, 36 with masting years, and 84 with small
mammal abundance. The results obtained show that the phenomenon of periodicity is present
in several species of birds in Quebec and that certain phenomena strongly regulate the
numbers of these same species. However, this study only provides early steps in our
understanding of periodicity in bird populations in Quebec. Future work will hopefully build
on the current study to increase our understanding of avian periodicity.
141.3 Introduction
In a recent paper, Rosenberg et al. (2019) highlight the decline of several species of the North
American avifauna over the past 50 years. The latest State of Canada’s Birds (2019) also
reports a decline in several groups of species, and an increase in other groups. One of the
lesser-known aspects of bird trends is their periodicity. Periodicity occurs when a population
fluctuates more regularly than expected by chance (Moss and Watson 2001). Periodicity is
well documented for some taxa, but only few bird species are known to show population
cycles (Cheveau et al. 2004; Koenig 2001; Moss et Watson 2001; Shaw et al. 2004).
Density-dependence can generate periodicity in avian populations, e.g. in response to
predation or food limitation (Newton 2007; Morrissette et al. 2010). Density-dependent and
density-independent factors can be periodic and thus induce cycles in bird populations. For
example, small mammal abundance often shows cycles varying from 3 to 7 years (Cheveau
et al. 2004), which can have a direct effect on bird populations above and beyond density-
dependent factors. Birds of prey in Canadian boreal forest sometimes exhibit abundance
cycles similar to those of their prey, such as the Great-horned Owl (Bubo virginianus) (Boutin
Mo 1995) and the Boreal Owl (Aegolius funereus) (Cheveau et al. 2004). Masting years, i.e.,
years when trees produce abundant seed and cones, can also induce cycles in bird
populations. In the boreal forest, 2- to 3-year masting cycles are observed, particularly among
conifers (Strong et al. 2015), and they are associated to the abundance of bird species such
as the Red-breasted Nuthatch (Sitta canadensis), the Common Redpoll (Acanthis flammea)
and the Pine Siskin (Spinus pinus) (Koenig 2001). Periodicity in bird populations could also
be linked to climate because of its influence on primary production and trophic links. The
North Atlantic Oscillation (NAO) is a climatic and atmospheric phenomenon that affects
precipitations and temperature in the North hemisphere, and thus in the boreal ecosystems
(Savard et al. 2015). NAO does not have a dominant cycle, but 75% of its variation occurs
within decades (Ottersen et al. 2001). Because of its influence on the climate, NAO can affect
bird populations and their reproductive success (Nott et al. 2002). In Québec, Sharp-shinned
Hawk (Accipiter striatus) abundance is positively correlated with the NAO (Savard et al.
152015). We know that NAO, masting years and small mammal abundance have an impact on
populations, but our understanding of their general influence on bird species is very limited.
The analysis of periodicity requires long and detailed time series. Fortunately, birds are one
of the most documented taxa and there are several data sources available. In Quebec, birders
have routinely sent their monthly observations to bird clubs since the 1970s, leading to the
first birdwatching checklist program in North America and perhaps the world, Étude des
Populations d’Oiseaux du Québec (ÉPOQ; Larivée 2001). This program and other similar
endeavours inspired the creation of eBird, launched in 2002 by the Cornell Lab of
Ornithology and the National Audubon Society (Sullivan et al. 2009). Although these data
contain substantial sources of variability (Dunn et al. 1996; Walker et Taylor 2017), they
remain reliable for trend analyses (Walker et Taylor 2020).
The objectives of this study are to (1) quantify trends to determine the proportion of the bird
species observed in Quebec since 1970 that show cycles with periods ranging from 2 to 13
years, and (2) explore three possible causes of the periodicity: North Atlantic Oscillation,
masting years and small mammal abundance.
1.4 Methods
We selected the province of Québec, Canada as the geographic region of interest. In this
study, we used data from eBird to draw up the Québec bird species trends and to determine
if the populations of those species show a periodicity. The eBird dataset is available online
and we downloaded the 2019 dataset (eBird basic dataset 2019). We added ÉPOQ checklists
to the eBird dataset to complement eBird especially preceding 2002, when eBird was
launched. Also, most of the data from the two Quebec bird atlases are included. The
checklists from both datasets contain information about the number of individuals observed,
species, age, sex of the individuals, the date, the location, and the protocol type. We excluded
checklists before 1970 as they were too sparse prior that year. Therefore, we have data for
each year, from 1970 to 2019. Although studies using eBird data often use only checklists
with known effort (protocol, duration, etc.), we decided to include all checklists in our
analysis (with and without known effort), as did Walker and Taylor (2017). One of the
16Vous pouvez aussi lire

















































