PLEINS FEUX (ARCHIVÉ) CONDITIONS PROPICES À L'ÉMERGENCE DE NOUVEAUX VARIANTS DU SRAS-COV-2
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
PLEINS FEUX
(ARCHIVÉ) Conditions propices à l’émergence
de nouveaux variants du SRAS-CoV-2
Publication : mars 2022
Archivé : décembre 2023
ARCHIVÉ
VÉ
Ce matériel archivé est disponible uniquement à des fins de recherche historique et de référence.
Celui-ci n'est plus mis à jour et il se peut qu’il ne reflète plus les directives actuelles.
I
CH
AR
Principales constatations
La transmission continue est le principal moteur de l’évolution du coronavirus du syndrome
respiratoire aigu sévère 2 (SRAS-CoV-2). Des taux élevés de transmission ouvrent la voie à
l’émergence de nouveaux variants préoccupants (VP) en raison d’un phénomène de sélection
naturelle.
Des facteurs interreliés comme l’émergence de sources zoonotiques et la transmission
interspécifique, de même que l’infection chronique chez les patients immunosupprimés,
peuvent accroître davantage le risque de voir apparaître de nouveaux variants.
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 1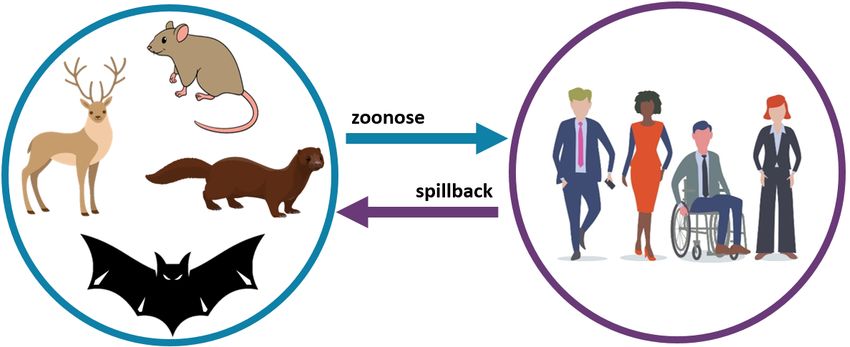
Une vaccination rapide et globale de la population, associée à la mise en œuvre de mesures
sanitaires et à la surveillance, constitue une démarche proactive prudente pour réduire
l’émergence et la propagation de nouveaux variants du SRAS-CoV-2.
L’émergence de variants dans les régions où le taux de vaccination est faible touche autant les
régions à taux élevé de vaccination que les régions à faible taux de vaccination, ce qui renforce
l’importance de l’équité mondiale en matière de vaccination pour réduire le risque continu
d’émergence de nouveaux variants préoccupants.
Introduction
L’émergence de variants du SRAS-CoV-2 survient généralement en raison de mutations pendant
la réplication virale, et moins fréquemment par recombinaison génétique (recombinaison
lorsque différentes souches virales infectent le même hôte) et l’édition de l’acide ribonucléique
VÉ
(ARN) par passage sur l’hôte. À l’instar d’autres virus à ARN, les coronavirus, dont le SRAS-CoV-2,
sont sujets à de fréquentes mutations, bien que dans une moindre mesure que l’influenzavirus,
en raison de la présence d’un domaine d’exonucléase de relecture.1 Avec le temps, certaines
modifications génétiques sont naturellement sélectionnées et peuvent mener à des altérations
de la transmissibilité, de la virulence, de l’échappement immunitaire et de l’efficacité vaccinale.
Les mutations peuvent soit augmenter, réduire ou être sans effet sur l’infectivité virale. Les variants
qui ont évolué pour devenir plus transmissibles, causer une maladie plus grave ou contribuer à la
I
diminution de l’efficacité des mesures de santé publique s’appellent des « variants préoccupants
(VP) ». Par ailleurs, les variants qui ont des mutations dont on ignore si elles augmentent la
CH
transmissibilité ou la virulence s’appellent des « variants d’intérêt (VI) » et ils peuvent
éventuellement passer au statut de variant préoccupant en présence de données en matière de
transmissibilité ou de virulence.2 Plusieurs VP ont émergé de la lignée ancestrale du SRAS-CoV-2,
incluant Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2 et ses sous-lignées AY) et
plus récemment, Omicron (B.1.1.529 et ses sous-lignées BA) et son sous-variant BA.2.
À l’heure actuelle, les variants préoccupants et les variants d’intérêt du SRAS-CoV-2 ont en
commun plusieurs mutations qui ont permis à des variants ultérieurs de se propager dans des
AR
populations largement vaccinées, tout en conservant ou en augmentant leur capacité
réplicative, c’est-à-dire leur capacité de se répliquer dans un environnement donné.3 Le concept
de sélection naturelle suggère que des mutations qui privilégient une capacité réplicative
accrue, notamment une réplication ou une transmissibilité améliorée, seront favorisées.4
La majorité des mutations du SRAS-CoV-2 surviennent dans le gène de spicule, ce qui
correspond à la protéine de spicule (S) qui est située à la surface de l’enveloppe virale.5,6 La
protéine S joue un rôle de premier plan dans la reconnaissance du récepteur hôte (enzyme de
conversion de l’angiotensine 2 [ACE2]) et la pénétration cellulaire du virus.7 De plus, la protéine
S constitue une cible essentielle pour les anticorps neutralisants, et les vaccins actuels utilisent
la protéine S comme une substance immunogène, ou une substance déclenchant une réponse
immunitaire, soit seule ou avec d’autres protéines du SRAS-CoV-2.7
Les taux élevés de transmission continue du variant Omicron ont suscité des préoccupations
concernant la formation de variants nouveaux ou émergents susceptibles d’affecter l’efficacité
vaccinale et d’accroître davantage la transmissibilité et la gravité du SRAS-CoV-2.
Le présent document vise à présenter un résumé des données probantes sur les conditions et
les facteurs de risque propices à l’émergence de nouveaux variants du SRAS-CoV-2.
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 2Méthodologie
Des recherches ont été effectuées dans Ovid Medline et Google Scholar et une recherche de
prépublications a également été menée dans le dossier sur la COVID-19 du National Institutes of Health
(NIH). Les concepts de recherche suivants étaient inclus : SRAS-CoV-2/COVID-19 et variants émergents.
Résultats
Au niveau populationnel, nous ne connaissons pas tous les facteurs de risque d’émergence de nouveaux
variants. À la lumière des conclusions préliminaires de l’évaluation du SRAS-CoV-2 et de nos
connaissances sur les autres virus à ARN, nombre de facteurs peuvent favoriser l’émergence de
nouveaux variants du SRAS-CoV-2. De façon générale, les virus à ARN sont sujets à des mutations
aléatoires, en raison de l’absence d’activité exonucléase de relecture des ARN polymérases codées du
virus. Les mutations surviennent naturellement comme un sous-produit de la réplication virale. Le taux
VÉ
de mutation dans le génome du SRAS-CoV-2 a été estimé à 1,87 x 10-6 substitutions de nucléotides par
site par jour, ce qui est approximativement cinq fois moins élevé que l’infuenza A/H3N2, mais plus élevé
que de nombreux autres virus à ARN monocaténaires. Par conséquent, pour l’ensemble des ~30 000
paires de bases du génome du SRAS-CoV-2, une lignée subit approximativement 20 modifications
génétiques par année.1,8 Chaque mutation additionnelle s’accompagne d’un risque accru d’émergence
d’un nouveau variant. La transmission et la propagation rapide du SRAS-CoV-2 à l’échelle mondiale,
entraînant une infection et une réplication virales, fournit au virus de nombreuses occasions de
procéder à la sélection naturelle de mutations si rares, mais favorables. La sélection naturelle favorise
I
l’émergence de variants préoccupants qui échappent à l’immunité conférée par l’infection ou la
vaccination, en particulier dans le contexte d’une immunité partielle de la population.9
CH
Facteurs augmentant l’occurrence de mutations favorables
Plusieurs facteurs peuvent accroître l’occurrence de mutations favorables dans la population.10,11
Ces facteurs sont énumérés et décrits ci-dessous.
NOMBRE DE PERSONNES INFECTÉES
Plus le virus se répand, plus il a d’occasions de muter. Bien que la majorité des mutations qui
AR
surviennent dans le SRAS-CoV-2 soient néfastes pour le virus et rapidement éliminées de la population
ou encore relativement neutres, une faible proportion d’entre elles touchera des propriétés
fonctionnelles susceptibles de modifier l’infectivité, la gravité de la maladie ou les interactions avec le
degré d’immunité de l’hôte.
SUPERPROPAGATEURS
Certaines personnes peuvent transmettre le SRAS-CoV-2 à un taux disproportionnellement élevé par
rapport à d’autres, ce sont les superpropagateurs. Les scénarios de superpropagation pourraient jouer
un rôle important en favorisant certains variants du SRAS-CoV-2, accroissant ainsi leur prédominance
dans la population.12
HÔTES IMMUNOSUPPRIMÉS
Les mutations se produisent au moment de la réplication du virus dans un hôte, et quand un nouveau
variant apparaît pendant une infection, les mutations favorables seront naturellement sélectionnées.
Dans le cas du SRAS-CoV-2, ces processus évolutifs ont été le mieux documentés chez les patients
immunosupprimés.13 Ces derniers peuvent maintenir des charges virales élevées pendant des périodes
prolongées, créant ainsi davantage d’occasions de réplication et de sélection virales, et conduisant à des
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 3taux de mutation élevés. De plus, une infection qui se prolonge peut augmenter la durée d’excrétion des
nouveaux variants.14,15
Grâce au séquençage du SRAS-CoV-2 à de multiples points temporels, plusieurs études ont réussi à
cerner des modifications rapides dans la composition de la population virale chez un patient au fil des
jours.16–18 Ces modifications sont plus rapides que prévu par dérive génétique dans une vaste
population, et portent à croire que pour le virus, la sélection naturelle consiste à se répliquer ou à
échapper à un système immunitaire affaibli. Les conséquences de santé publique pour les patients
immunosupprimés infectés par le SRAS-CoV-2 s’en trouvent ainsi exacerbées. La vaccination des
patients immunosupprimés et de leurs contacts étroits, ainsi que l’observance stricte de l’isolement en
cas d’infection devraient être hautement prioritaires.13
TRANSMISSION INTERSPÉCIFIQUE
Le SRAS-CoV-2 pourrait provenir d’un coronavirus zoonotique, possiblement présent dans des chauves-
VÉ
souris qui l’auraient transmis aux humains par un mammifère hôte non humain intermédiaire.19–21
L’évolution des coronavirus, y compris le SRAS-CoV-2, est compliquée par la vaste gamme d’hôtes
potentiels et leur transmissibilité connue de l’homme aux animaux et des animaux à l’homme.22–24 Les
coronavirus peuvent subir et un cycle enzootique, dans lequel un virus est transmis à des animaux
autres qu’humains, et un cycle épidémique, dans lequel un virus est transmis entre humains. Puisque les
animaux sont des réservoirs pour le SRAS-CoV-2, la sélection d’une transmission supérieure à l’intérieur
de ces espèces pourrait générer des caractéristiques pathologiques différentes et contribuer à accélérer
l’évolution du virus.25 S’il y a infection d’une population d’animaux autres qu’humains par une source
I
humaine du SRAS-CoV-2 (phénomène connu sous le nom zoonose inverse ou spillback), suivie de
mutations virales à l’intérieur de la population animale non humaine et d’une transmission secondaire
CH
des animaux vers les humains (phénomène connu sous le nom de zoonose ou spillover), le cycle de
transmission interspécifique peut contribuer à l’émergence de nouveaux variants du SRAS-CoV-2 (voir
Figure 1).14 Par exemple, des rapports ont fait état de la transmission d'animal à humain de nouveaux
variants du SRAS-CoV-2 à partir de visons d’élevage dans les Pays-Bas et le Danemark;26 cependant, ces
variants n’étaient pas hautement transmissibles.27
Figure 1 : Cycles proposés de transmission enzootique et épidémique du SRAS-CoV-2
AR
[IMAGE : zoonosis = zoonose; spillback = spillback]
Le SRAS-CoV-2 est probablement apparu chez les chauves-souris, avec transmission à un hôte intermédiaire
encore inconnu ou directement aux humains. Le cycle enzootique inclut la transmission entre et à l’intérieur de
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 4populations d’espèces sauvages (p. ex., cerfs de Virginie, rongeurs) et d’animaux domestiques ou en captivité (p.
ex., visons). Le cycle épidémique inclut la transmission du SRAS-CoV-2 entre humains. Bien que la transmission des
humains aux animaux (spillback) ait été documentée, les voies de transmission des animaux aux humains (zoonose
ou spillover) restent à préciser. Termes clés : enzootique, transmission d’un agent pathogène entre des animaux
autres qu’humains aux taux prévus, avec peu de décès chez les animaux infectés; épidémique, transmission d’un
agent pathogène entre des humains au-delà des taux prévus; zoonose, transmission d’un agent pathogène
d’animaux autres qu’humains à des humains (aussi appelée spillover ou spillover zoonotique); spillback,
transmission d’un agent pathogène d’humains à des animaux autres qu’humains (aussi appelée zoonose inverse ou
zooanthroponose).
Plus récemment, des analyses moléculaires ont révélé la possibilité que le variant Omicron du SRAS-CoV-
2 ait une origine murine, dans laquelle il pourrait y avoir eu spillback des humains aux souris dès le
milieu de l’année 2020, et transmission inverse vers les humains à la fin de l’année 2021, ce qui pourrait
expliquer le nombre relativement élevé de mutations dans le virus et les différences évolutives par
rapport aux précédents variants préoccupants, notamment le variant Delta.28 Toutefois, l’absence d’une
VÉ
infectivité accrue dans les modèles de rongeurs n'appuie pas l’origine murine du variant Omicron pour
l'instant.29 Il a été rapporté que plusieurs autres animaux hébergeaient et transmettaient le SRAS-CoV-2,
comme les furets, les chats, les ratons-laveurs, les cerfs de Virginie et les hamster de Syrie.30–32
TRAITEMENT PHARMACOLOGIQUE, BIOLOGIQUE ET PAR PLASMA CONVALESCENT
Le recours à certains médicaments et agents biologiques pour traiter la COVID-19 pourrait en théorie
appliquer une pression de sélection propice à l’émergence de nouveaux variants.33,34 Des séries de cas
ont par exemple indiqué que le traitement par le bamlanivimab, un anticorps monoclonal dirigé contre
I
la protéine de spicule du SRAS-CoV-2, avait entraîné l’émergence de la mutation résistante au
CH
bamlanivimab E484K chez plusieurs patients, laquelle peut provoquer une infection persistante, en
particulier chez des patients immunosupprimés.35,36 De même, des cas d’infection continue causée par
des variants d’échappement immunitaire ayant émergé durant un traitement par plasma convalescent
ont été signalés.33 Outre le traitement à base d’anticorps, la résistance aux médicaments constitués de
petites molécules comme le remdésivir a été rapportée.37 Cependant, les données cliniques sur la
prévalence de tels mutants et l’existence de différences en ce qui a trait à leur risque de transmissibilité
ou leur virulence sont insuffisantes. Étant donné que la pharmacothérapie n’est pas le principal
mécanisme de contrôle du SRAS-CoV-2, il est peu probable qu’elle joue un rôle significatif dans
l’émergence de nouveaux variants. Par ailleurs, la durée limitée de l’infection et le risque réduit de
AR
réplication virale chez les patients traités indiquent un plus faible risque de développement d’une
résistance en cours de traitement.38 Notamment, l’émergence de nouveaux variants attribuable à
d’autres mécanismes a rendu certains agents biologiques inefficaces, ce qui réduit les options
thérapeutiques contre la COVID-19. Par exemple, le « cocktail » d’anticorps casirivimab/imdevimab n’a
pas d’activité neutralisante contre le variant Omicron.39
RECOMBINAISON AVEC D’AUTRES CORONAVIRUS
Au lieu de mutations périodiques dans le temps, de grandes sections du génome du SRAS-CoV-2
pourraient se recombiner avec un autre coronavirus (dans un hôte humain ou un autre mammifère
hôte) et mener à la création d’un nouveau variant.38 Ce potentiel de recombinaison peut favoriser la
transmission interspécifique.19 Nous savons que d’autres coronavirus subissent des recombinaisons
moins fréquentes, et d’aucuns ont émis l’hypothèse qu’un événement de recombinaison serait un
facteur clé pour déterminer l’origine du SRAS-CoV-2.40 De plus, des analyses phylogénétiques révèlent
que le variant préoccupant Alpha (B.1.1.7) pourrait être issu d’un événement de recombinaison avec des
souches préexistantes du SRAS-CoV-2.41
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 5Rôle de la vaccination
En raison d’une combinaison possible d’échappement immunitaire et de modifications intrinsèques
de la transmission virale, Omicron a entraîné une transmissibilité accrue et des taux supérieurs
d’infections après la vaccination en Ontario et à l’échelle mondiale. Bien que la vaccination soit
notablement moins efficace contre le variant Omicron, par comparaison aux lignées précédentes,
trois doses d’un vaccin contre la COVID-19 offrent néanmoins une protection importante contre
l’infection et la maladie grave causées par le variant préoccupant Omicron.42,43
D’une part, l’évolution du SRAS-CoV-2 dans le contexte d’une population largement, mais non
entièrement vaccinée (ou avec une immunité affaiblie avec le temps) peut mener à la sélection de
variants qui échappent à la protection immunitaire conférée par la vaccination.44 D’autre part, une
population rapidement et largement vaccinée qui est équitablement et géographiquement dispersée
peut réduire l’infection par le SRAS-CoV-2 et sa transmission, et ainsi atténuer la pression de
sélection et freiner la propagation de nouveaux variants. 45
VÉ
Bien que la vaccination puisse être associée à l’émergence de mutants du SRAS-CoV-2 résistants aux
anticorps,46 la rapidité de la vaccination et l’ampleur de la couverture vaccinale ainsi que les mesures
de santé publique comme le port du masque, la distanciation physique et la recherche de contacts,
peuvent contribuer à ralentir la propagation de ces mutants.47 Une étude de modélisation menée par
Rella et coll. a révélé qu’une vaccination rapide de la population réduit la probabilité d’émergence de
souche du SRAS-CoV-2 résistante aux vaccins. Cette probabilité peut être davantage réduite si la
vaccination est associée au respect des mesures sanitaires (p. ex., observance stricte du port du
I
masque et de la distanciation physique) utiles pour freiner la transmission du virus, en particulier à la
CH
suite d’une campagne de vaccination.48 L’émergence de variants dans les régions où les taux de
vaccination sont moins élevés affectera tant les régions où la population est peu vaccinée que celles
où la population est largement vaccinée, renforçant l’importance de l’équité mondiale en matière de
vaccination pour limiter le risque continu d’émergence de nouveaux variants préoccupants.
On s’interrogeait au départ à savoir si les stratégies visant à réduire les doses nécessaires à la
vaccination, comme les intervalles prolongés entre les doses, pouvaient créer un environnement
propice à l’émergence de nouveaux variants en raison de l’immunité partielle de la population.
Cependant, les données probantes sur la vaccination contre le SRAS-CoV-2 et d’autres virus
AR
indiquent que ces stratégies diminuent au contraire le risque d’émergence de nouveaux variants
parce que la vaccination réduit la charge virale ainsi que la durée de l’infection. 49
Il y a lieu de penser que la diversité des lignées de SRAS-CoV-2 est inversement corrélée avec
l’augmentation des taux de vaccination massive de la population. Cela appuie le concept selon
lequel des taux plus élevés de vaccination peuvent contribuer à limiter l’évolution du SRAS-CoV-
2 et ainsi, stopper l’émergence de nouveaux variants.50
Rôle des mesures de santé publique
Comme nous l’avons vu dans l’étude de modélisation de Rella et coll., même avec des taux
élevés de vaccination, les mesures sanitaires non pharmaceutiques peuvent aider à réduire
l’émergence de souches virales résistantes à la vaccination en compensant l’efficacité d’une
vaccination partielle et en atténuant la transmissibilité du SRAS-CoV-2 dans la population.
Il y a lieu de croire que la rigueur des interventions en santé publique (p. ex., mesures de
confinement) n’augmente pas le risque d’émergence de nouveaux variants du SRAS-CoV-2, car
ces mesures tendent à diminuer la transmission virale, quelles que soient les caractéristiques du
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 6virus. En fait, Justo Arevalo et coll. ont observé que l’introduction de mesures sanitaires était
associée à une réduction similaire tant des variants hautement transmissibles (en particulier la
mutation N501Y) que des variants moins transmissibles du SRAS-CoV-2.51
Rôle de la surveillance
Le dépistage proactif et le séquençage du génome (p. ex., dans le cas des voyages à l’étranger) peuvent
permettre de détecter les variants émergents et de déclencher promptement des interventions de santé
publique visant à freiner leur introduction et leur propagation dans une autre communauté.52
Bien que les humains représentent certainement le plus vaste réservoir du SRAS-CoV-2, il faut accroître
la surveillance virale et le séquençage chez les animaux afin d’améliorer la détection précoce de
nouveaux variants zoonotiques susceptibles de contaminer les humains.28
Conclusion
VÉ
L’émergence de nouveaux variants du SRAS-CoV-2 est un phénomène inévitable cependant, les
conditions dans lesquelles ces variants apparaissent et prospèrent restent en grande partie à
déterminer. Par ailleurs, différents mécanismes tous en lien avec le concept de « sélection
naturelle » sont proposés, c’est-à-dire que les mutations qui permettent au virus d’échapper à la
réponse immunitaire sont plus susceptibles de devenir prédominantes avec le temps. La
transmission continue augmente sans équivoque le risque de mutations, lequel est davantage
I
accru par les infections chroniques et le potentiel de transmission zoonotique.
CH
De nouveaux variants continuent d’évoluer en vue d’échapper à la réponse immunitaire conférée
par la vaccination ou par une infection précédente. Cependant, une vaccination mondialement
équitable, rapide et à grande échelle couplée à d’autres interventions en santé publique constitue
la meilleure approche pour freiner l’émergence et la propagation de nouveaux variants.
AR
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 7Références
1. Otto SP, Day T, Arino J, Colijn C, Dushoff J, Li M, et al. The origins and potential future of SARS-CoV-
2 variants of concern in the evolving COVID-19 pandemic. Current Biology. 2021;31:R918–29.
Disponible à : https://doi.org/10.1016/j.cub.2021.06.049
2. Organisation mondiale de la santé. Suivi des variants du SARS-CoV-2 [En ligne]. Genève :
Organisation mondiale de la santé; 2022 [cité le 28 janvier 2022]. Disponible à :
https://www.who.int/fr/activities/tracking-SARS-CoV-2-variants/tracking-SARS-CoV-2-variants
3. Domingo E, Holland JJ. RNA virus mutations and fitness for survival. Annu Rev Microbiol.
1997;51:151-78. Disponible à : https://doi.org/10.1146/annurev.micro.51.1.151
4. Plante JA, Mitchell BM, Plante KS, Debbink K, Weaver SC, Menachery VD. The variant gambit:
VÉ
COVID-19’s next move. Cell Host Microbe. 2021;29(4):508-15. Disponible à :
https://doi.org/10.1016/j.chom.2021.02.020
5. Tao K, Tzou PL, Nouhin J, Gupta RK, de Oliveira T, Kosakovsky Pond SL, et al. The biological and
clinical significance of emerging SARS-CoV-2 variants. Nat Rev Genet. 2021;22:757-73. Disponible à :
https://doi.org/10.1038/s41576-021-00408-x
6. Chen J, Gao K, Wang R, Wei G-W. Revealing the threat of emerging SARS-CoV-2 mutations to
I
antibody therapies. J Mol Biol. 2021;433:167155. Disponible à :
https://doi.org/10.1016/j.jmb.2021.167155
CH
7. Heinz FX, Stiasny K. Distinguishing features of current COVID-19 vaccines: knowns and unknowns of
antigen presentation and modes of action. NPJ Vaccines. 2021;6(1):104. Disponible à :
https://doi.org/10.1038/s41541-021-00369-6
8. Su S, Wong G, Shi W, Liu J, Lai ACK, Zhou J, et al. Epidemiology, genetic recombination, and
pathogenesis of coronaviruses. Trends Microbiol. 2016;24(6):490-502. Disponible à :
https://doi.org/10.1016/j.tim.2016.03.003
AR
9. Gräf T, Bello G, Venas TMM, Pereira EC, Paixão ACD, Appolinario LR, et al. Identification of a novel
SARS-CoV-2 P.1 sub-lineage in Brazil provides new insights about the mechanisms of emergence of
variants of concern. Virus Evol. 2021;7(2):veab091. Disponible à :
https://doi.org/10.1093/ve/veab091
10. Van Egeren D, Novokhodko A, Stoddard M, Tran U, Zetter B, Rogers MS, et al. Controlling long-term
SARS-CoV-2 infections can slow viral evolution and reduce the risk of treatment failure. Sci Rep.
2021;11(1):22630. Disponible à : https://doi.org/10.1038/s41598-021-02148-8
11. Laskar R, Ali S. Differential mutation profile of SARS-CoV-2 proteins across deceased and
asymptomatic patients. Chem Biol Interact. 2021;347:109598. Disponible à :
https://doi.org/10.1016/j.cbi.2021.109598
12. Gómez-Carballa A, Pardo-Seco J, Bello X, Martinón-Torres F, Salas A. Superspreading in the
emergence of COVID-19 variants. Trends Genet. 2021;37(12):1069-80. Disponible à :
https://doi.org/10.1016/j.tig.2021.09.003
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 813. Corey L, Beyrer C, Cohen MS, Michael NL, Bedford T, Rolland M. SARS-CoV-2 variants in patients
with immunosuppression. N Engl J Med. 2021;385(6):562-6. Disponible à :
https://doi.org/10.1056/NEJMsb2104756
14. Burki T. The origin of SARS-CoV-2 variants of concern. Lancet Infect Dis. 2022;22(2):174-5.
Disponible à : https://doi.org/10.1016/s1473-3099(22)00015-9
15. Pan D, Mudalige NL, Sze S, Koeckerling D, Oyefeso O, Barker J, et al. The new UK SARS-CoV-2 variant
and lockdown – causes and consequences. Clin Med. 2021;21:e295-9. Disponible à :
https://doi.org/10.7861/clinmed.2021-0019
16. Choi B, Choudhary MC, Regan J, Sparks JA, Padera RF, Qiu X, et al. Persistence and evolution of
SARS-CoV-2 in an immunocompromised host. N Engl J Med. 2020;383(23):2291-3. Disponible à :
https://doi.org/10.1056/NEJMc2031364
VÉ
17. Avanzato VA, Matson MJ, Seifert SN, Pryce R, Williamson BN, Anzick SL, et al. Case study: prolonged
infectious SARS-CoV-2 shedding from an asymptomatic immunocompromised individual with
cancer. Cell. 2020;183(7):1901-1912.e9. Disponible à : https://doi.org/10.1016/j.cell.2020.10.049
18. Kemp SA, Collier DA, Datir RP, Ferreira IATM, Gayed S, Jahun A, et al. SARS-CoV-2 evolution during
treatment of chronic infection. Nature. 2021;592(7853):277-82. Disponible à :
https://doi.org/10.1038/s41586-021-03291-y
I
19. Na W, Moon H, Song D. A comprehensive review of SARS-CoV-2 genetic mutations and lessons from
CH
animal coronavirus recombination in one health perspective. J Microbiol. 2021;59(3):332-40.
Disponible à : https://doi.org/10.1007/s12275-021-0660-4
20. Terrier O, Si-Tahar M, Ducatez M, Chevalier C, Pizzorno A, Le Goffic R, et al. Influenza viruses and
coronaviruses: Knowns, unknowns, and common research challenges. PLoS Pathog.
2021;17(12):e1010106. Disponible à : https://doi.org/10.1371/journal.ppat.1010106
21. Rehman SU, Shafique L, Ihsan A, Liu Q. Evolutionary trajectory for the emergence of novel
coronavirus SARS-CoV-2. Pathogens. 2020;9(3):E240. Disponible à :
AR
https://doi.org/10.1186/s12985-021-01633-w
22. Damas J, Hughes GM, Keough KC, Painter CA, Persky NS, Corbo M, et al. Broad host range of SARS-
CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc Natl Acad Sci
USA. 2020;117(36):22311-22. Disponible à : https://doii.org/10.1073/pnas.2010146117
23. Kuchipudi SV, Surendran-Nair M, Ruden RM, Yon M, Nissly RH, Nelli RK, et al. Multiple spillovers
and onward transmission of SARS-CoV-2 in free-living and captive white-tailed deer. bioRxiv 466677
[Prépublication]. 1er novembre 2021 [cité le 1er mars 2022]. Disponible à :
https://doi.org/10.1101/2021.10.31.466677
24. Saif LJ. Animal coronaviruses : what can they teach us about the severe acute respiratory
syndrome?. Rev Sci Tech. 2004;23(2):643-60. Disponible à : https://doi.org/10.20506/rst.23.2.1513
25. Bashor L, Gagne RB, Bosco-Lauth A, Bowen R, Stenglein M, VandeWoude S. SARS-CoV-2 evolution in
animals suggests mechanisms for rapid variant selection. Proc Natl Acad Sci U S A.
2021;118(44):e2105253118. Disponible à : https://doi.org/10.1073/pnas.2105253118
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 926. Wang L, Didelot X, Bi Y, Gao GF. Assessing the extent of community spread caused by mink-derived
SARS-CoV-2 variants. Innovation. 2021;2(3):100128. Disponible à :
https://doi.org/10.1016/j.xinn.2021.100128
27. Oreshkova N, Molenaar RJ, Vreman S, Harders F, Oude Munnink BB, Hakze-van der Honing RW, et
al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Eurosurveillance.
2020;25(23):2001005. Disponible à : https://doi.org/10.2807/1560-7917.ES.2020.25.23.2001005
28. Wei C, Shan K-J, Wang W, Zhang S, Huan Q, Qian W. Evidence for a mouse origin of the SARS-CoV-2
Omicron variant. J Genet Genomics. 2021;48:1111-21. Disponible à :
https://doi.org/10.1016/j.jgg.2021.12.003
29. Halfmann PJ, Iida S, Iwatsuki-Horimoto K, Maemura T, Kiso M, Scheaffer SM, et al. SARS-CoV-2
Omicron virus causes attenuated disease in mice and hamsters. Nature. 21 janvier 2022
[Publication électronique avant impression] Disponible à : https://doi.org/10.1038/s41586-022-
VÉ
04441-6
30. Mallapaty S. The search for animals harbouring coronavirus - and why it matters. Nature.
2021;591(7848):26-8. Disponible à : https://doi.org/10.1038/d41586-021-00531-z
31. Mallapaty S. How sneezing hamsters sparked a COVID outbreak in Hong Kong. Nature.
4 février 2022 [Publication électronique avant impression]. Disponible à :
I
https://doi.org/10.1038/d41586-022-00322-0
CH
32. Palermo PM, Orbegozo J, Watts DM, Morrill JC. SARS-CoV-2 Neutralizing antibodies in white-tailed
deer from Texas. Vector Borne Zoonotic Dis. 2022;22(1):62-4. Disponible à :
https://doi.org/10.1089/vbz.2021.0094
33. Focosi D, Maggi F, Franchini M, McConnell S, Casadevall A. Analysis of immune escape variants from
antibody-based therapeutics against COVID-19: a systematic review. Int J Mol Sci. 2021;23(1):29.
Disponible à : https://doi.org/10.3390/ijms23010029
34. Colson P, Devaux CA, Lagier J-C, Gautret P, Raoult D. A possible role of remdesivir and plasma
AR
therapy in the selective sweep and emergence of new SARS-CoV-2 variants. J Clin Med.
2021;10(15):3276. Disponible à : https://doi.org/10.3390/jcm10153276
35. Peiffer-Smadja N, Bridier-Nahmias A, Ferré VM, Charpentier C, Garé M, Rioux C, et al. Emergence of
E484K mutation following bamlanivimab monotherapy among high-risk patients infected with the
Alpha variant of SARS-CoV-2. Viruses. 2021;13(8):1642. Disponible à :
https://doi.org/10.3390/v13081642
36. Jensen B, Luebke N, Feldt T, Keitel V, Brandenburger T, Kindgen-Milles D, et al. Emergence of the
E484K mutation in SARS-COV-2-infected immunocompromised patients treated with bamlanivimab
in Germany. Lancet Reg Health Eur. 2021;8:100164. Disponible à :
https://doi.org/10.1016/j.lanepe.2021.100164
37. Chakraborty C, Bhattacharya M, Sharma AR. Emerging mutations in the SARS-CoV-2 variants and
their role in antibody escape to small molecule-based therapeutic resistance. Curr Opin Pharmacol.
2022;62:64-73. Disponible à : https://doi.org/10.1016/j.coph.2021.11.006
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 1038. Amoutzias GD, Nikolaidis M, Tryfonopoulou E, Chlichlia K, Markoulatos P, Oliver SG. The remarkable
evolutionary plasticity of coronaviruses by mutation and recombination: insights for the COVID-19
pandemic and the future evolutionary paths of SARS-CoV-2. Viruses. 2022;14(1):78. Disponible à :
https://doi.org/10.3390/v14010078
39. Boschi C, Colson P, Bancod A, Moal V, Scola BL. Omicron variant escapes therapeutic mAbs contrary
to eight prior main VOC. bioRxiv 474769 [Prépublication]. 3 janvier 2022 [cité le 1er mars 2022].
Disponible à : https://doi.org/10.1101/2022.01.03.474769
40. Fischer W, Giorgi EE, Chakraborty S, Nguyen K, Bhattacharya T, Theiler J, et al. HIV-1 and SARS-CoV-
2: Patterns in the evolution of two pandemic pathogens. Cell Host Microbe. 2021;29(7):1093-110.
Disponible à : https://doi.org/10.1016/j.chom.2021.05.012
41. Xie X, Lewis T-J, Green N, Wang Z. Phylogenetic network analysis revealed the recombinant origin of
the SARS-CoV-2 VOC202012/01 (B.1.1.7) variant first discovered in U.K. bioRxiv 449840
VÉ
[Prépublication]. 10 juillet 2021 [cité le 9 février 2022]. Disponible à :
https:/doi.org/10.1101/2021.06.24.449840
42. Agence ontarienne de protection et de promotion de la santé (Santé publique Ontario). Variant
préoccupant Omicron de la COVID-19 (B.1.1.529) : Évaluation du risque, 29 décembre 2021 [En
ligne]. Toronto, ON : Imprimeur de la Reine pour l'Ontario; 2021 [cité le 1er mars 2022]. Disponible
à : https://www.publichealthontario.ca/-/media/documents/ncov/voc/2022/01/covid-19-omicron-
I
b11529-risk-assessment-dec-29.pdf?sc_lang=fr
CH
43. Buchan SA, Chung H, Brown KA, Austin PC, Fell DB, Gubbay JB, et al. Effectiveness of COVID-19
vaccines against Omicron or Delta symptomatic infection and severe outcomes. medRxiv 21268565
[Prépublication]. 28 janvier 2022 [cité le 31 janvier 2022]. Disponible à :
https://doi.org/10.1101/2021.12.30.21268565
44. Chen J, Wang R, Wei G-W. Review of the mechanisms of SARS-CoV-2 evolution and transmission.
arXiv 210908148 [Prépublication]. 15 septembre 2021 [cité le 28 janvier 2022]. Disponible à :
http://arxiv.org/abs/2109.08148
AR
45. Sah P, Vilches TN, Moghadas SM, Fitzpatrick MC, Singer BH, Hotez PJ, et al. Accelerated vaccine
rollout is imperative to mitigate highly transmissible COVID-19 variants. EClinicalMedicine.
2021;35:100865. Disponible à : https://doi.org/10.1016/j.eclinm.2021.100865
46. Wang R, Chen J, Wei G-W. Mechanisms of SARS-CoV-2 evolution revealing vaccine-resistant
mutations in Europe and America. J Phys Chem Lett. 2021;12(49):11850-7. Disponible à :
https://doi.org/10.1021/acs.jpclett.1c03380
47. Hughes E, Binny R, Hendy S, James A. Predicting elimination of evolving virus variants. medRxiv
21259501 [Prépublication]. 25 juin 2021 [cité le 9 février 2022]. Disponible à :
https://doi.org/10.1101/2021.06.24.21259501
48. Rella SA, Kulikova YA, Dermitzakis ET, Kondrashov FA. Rates of SARS-CoV-2 transmission and
vaccination impact the fate of vaccine-resistant strains. Sci Rep. 2021;11(1):15729. Disponible à :
https://doi.org/10.1038/s41598-021-95025-3
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 1149. Cobey S, Larremore DB, Grad YH, Lipsitch M. Concerns about SARS-CoV-2 evolution should not hold
back efforts to expand vaccination. Nat Rev Immunol. 2021;21(5):330-5. Disponible à :
https://doi.org/10.1038/s41577-021-00544-9
50. Niesen MJM, Anand P, Silvert E, Suratekar R, Pawlowski C, Ghosh P, et al. COVID-19 vaccines
dampen genomic diversity of SARS-CoV-2: Unvaccinated patients exhibit more antigenic mutational
variance. medRxiv 21259833 [Prépublication]. 5 juillet 2021 [cité le 28 janvier 2022]. Disponible à :
https://doi.org/10.1101/2021.07.01.21259833
51. Justo Arevalo S, Zapata Sifuentes D, J. Huallpa C, Landa Bianchi G, Castillo Chávez A, Garavito-Salini
Casas R, et al. Dynamics of SARS-CoV-2 mutations reveals regional-specificity and similar trends of
N501 and high-frequency mutation N501Y in different levels of control measures. Sci Rep.
2021;11(1):17755. Disponible à : https://doi.org/10.1038/s41598-021-97267-7
52. Williams GH, Llewelyn A, Brandao R, Chowdhary K, Hardisty K-M, Loddo M. SARS-CoV-2 testing and
VÉ
sequencing for international arrivals reveals significant cross border transmission of high risk
variants into the United Kingdom. EClinicalMedicine. 2021;38:101021. Disponible à :
https://doi.org/10.1016/j.eclinm.2021.101021
I
CH
AR
(ARCHIVÉ) Conditions propices à l’émergence de nouveaux variants du SRAS-CoV-2 12Modèle proposé pour citer le document
Agence ontarienne de protection et de promotion de la santé (Santé publique Ontario). Conditions
propices à l’émergence de nouveaux variants du SRAS-CoV-2. Toronto, ON : Imprimeur de la Reine pour
l’Ontario; 2022.
Avis de non-responsabilité
Santé publique Ontario (SPO) a conçu le présent document. SPO offre des conseils scientifiques et
techniques au gouvernement, aux agences de santé publique et aux fournisseurs de soins de santé de
l’Ontario. Les travaux de SPO s’appuient sur les meilleures données probantes disponibles au moment
de leur publication. L’application et l’utilisation du présent document relèvent de la responsabilité des
utilisateurs. SPO n’assume aucune responsabilité relativement aux conséquences de l’application ou de
l’utilisation du document par quiconque. Le présent document peut être reproduit sans permission à
VÉ
des fins non commerciales seulement, sous réserve d’une mention appropriée de Santé publique
Ontario. Aucun changement ni aucune modification ne peuvent être apportés à ce document sans la
permission écrite explicite de Santé publique Ontario.
Santé publique Ontario
Santé publique Ontario est un organisme du gouvernement de l’Ontario voué à la protection et à la
promotion de la santé de l’ensemble de la population ontarienne, ainsi qu’à la réduction des iniquités en
I
matière de santé. Santé publique Ontario met les connaissances et les renseignements scientifiques les
CH
plus pointus du monde entier à la portée des professionnels de la santé publique, des travailleurs de la
santé de première ligne et des chercheurs.
Pour en savoir plus sur SPO, veuillez consulter : publichealthontario.ca.
©Imprimeur de la Reine pour l’Ontario, 2022
ARVous pouvez aussi lire

















































