UE 1 BIOCHIMIE FICHE DE COURS 1 STRUCTURE DES PROTEINES - CPCM
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
UNIVERSITE VERSAILLES SAINT QUENTIN
1er semestre 2019-2020 – Valérie Brocheriou
UE 1
BIOCHIMIE
FICHE DE COURS 1
STRUCTURE DES PROTEINES
CPCM – 106 Bd Saint Germain 75006 PARIS – Tel : 01.46.34.52.25
contact@prepa-cpcm.com / www.prepa-cpcm.com
Page | 1
CPCM- UE1B- FC1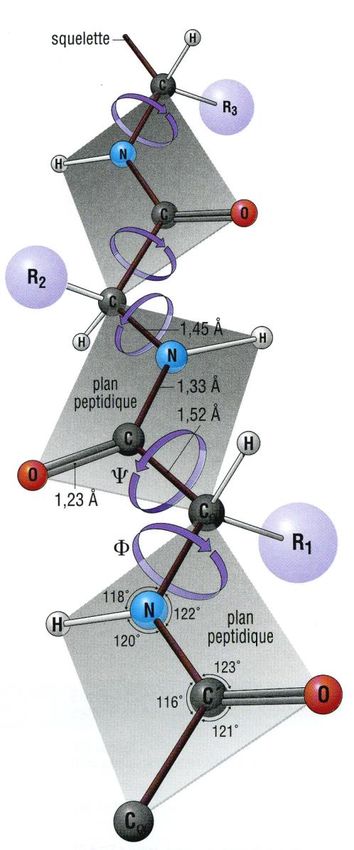
STRUCTURE DES PROTEINES
Table des matières
I. Les acides alpha- aminés p.3
1) Classification p.3
2) Les propriétés physico- chimiques des acides aminés p.5
3) Les modifications post- traductionnelles des acides aminés dans les protéines
p.7
4) Les rôles des acides aminés p.8
II. Les protéines p.8
1) Définition p.8
2) Liaison peptidique p.8
3) Les différents niveaux de structure des protéines p.9
4) La structure tertiaire p.15
5) Structure 3D et chaperons p.18
6) La structure quaternaire p.18
7) Pathologies de conformation des protéines p.19
Page | 2
CPCM- UE1B- FC1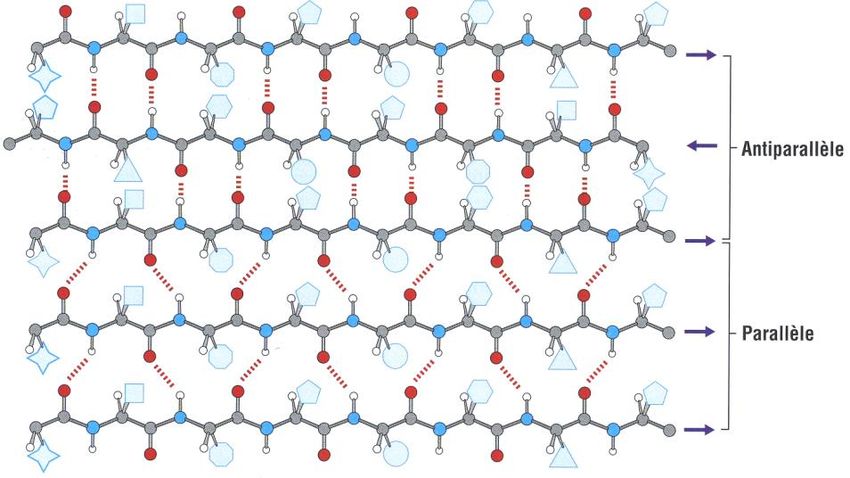
STRUCTURE DES PROTEINES
Les protéines sont composées d’une ou plusieurs chaînes peptidiques, identiques ou
différentes, composées chacune de l’enchainement d’acides alpha aminés. Cette chaîne peut
ensuite être modifiée par ajout d’ions, de sucres ou encore de lipides.
Les protéines font de 5000 à plusieurs millions de Daltons (unité de masse des protéines).
La chaîne d’acides aminés va adopter une certaine forme dans l’espace, qui lui confèrera sa
fonction (conformation native). Les acides aminés hydrophobes seront responsables au sein
de cette structure d’interactions hydrophobes, les acides aminés hydrophiles neutres de
liaisons hydrogènes, et les hydrophiles chargés de liaisons ioniques.
I. Les acides -aminés
.
Les acides -aminés sont les constituants majeurs des protéines. Le carbone qui porte la
fonction carboxyle, la plus oxydée, est nommé carbone alpha.
1) La classification : elle se fait selon la nature physico-chimique du reste (radical)
R.
Attention : Dans votre classification, polaire = hydrophile et apolaire = hydrophobe
En fonction de la polarité du radical R, on peut distinguer 4 classes d’acides aminés :
Classe 1 : Ni polaire ni apolaire, la glycine
Classe 2 : Les apolaires : Ala, Val, Leu, Ile (radical aliphatique) et Met, Pro, Phe, Trp
Classe 3 : Les polaires neutres : Ser ; Thr ; Cys ; Tyr ; Asn : Gln
Classe 4 : Les polaires chargés His, Lys ; Arg (chargés +) et Asp ; Glu (chargés -)
Cette polarité des radicaux va conditionner le reploiement tridimensionnel de la protéine.
1.1- Les acides -aminés à reste R apolaire aliphatique
Page | 3
CPCM- UE1B- FC1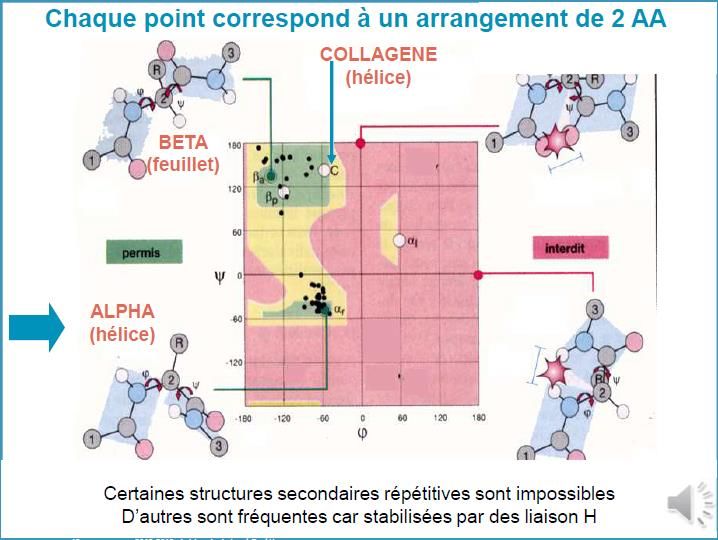
Remarques :
- La glycine ne possède pas d’atome de carbone asymétrique
- La glycine n’est ni polaire ni apolaire = classe 1 des acides aminés
- La glycine possède le radical R le plus petit possible (R = H) ; pas de carbone
asymétrique pas de pouvoir de déviation de la lumière polarisée
- L’isoleucine possède deux atomes de carbone asymétriques
- La proline possède un reste R cyclique ; amine secondaire
- La proline est le seul acide -aminé naturel à fonction amine secondaire
Page | 4
CPCM- UE1B- FC1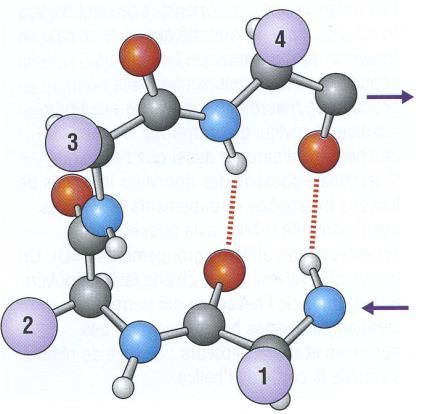
1.2- Les acides -aminés à reste R aromatique
Phe et Trp sont apolaires
Tyr est polaire
L’histidine est aussi un acide aminé aromatique, mais dont l’absorbance est négligeable.
1.3- Les acides -aminés à reste R polaire non ionisable
1.4- Les acides -aminés à reste R polaire ionisable
Page | 5
CPCM- UE1B- FC1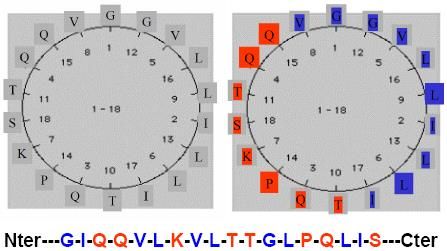
1.5- Les acides -aminés à reste R à fonction alcool ou soufrée
1.6- Les acides non -aminés
Ils ne sont pas constitutifs des protéines mais peuvent intervenir dans le métabolisme,
ou comme neurotransmetteurs :
- Ornithine et citrulline interviennent dans la synthèse de l’urée, le cycle de l’urée
permettant l’élimination de l’azote « toxique » chez l’homme
- L’acide glutamique peut être décarboxylé en acide -aminobutyrique (GABA,
neurotransmetteur)
- L’histidine peut être décarboxylée en histamine (neuromédiateur, impliquée dans les
phénomènes allergiques)
- La tyrosine peut être hydroxylée sur le noyau aromatique pour donner la DOPA, qui
sera décarboxylée pour former la dopamine
2) Les propriétés physico-chimiques des acides aminés
2.1- Les propriétés acido-basiques
Page | 6
CPCM- UE1B- FC1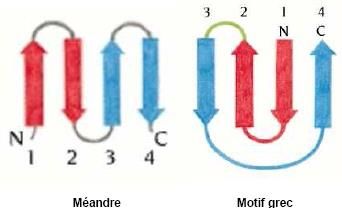
La présence d’une fonction acide carboxylique et d’une fonction basique (amine) confère aux
acides aminés un caractère ampholyte.
On pose pour simplifier que pKa COOH / COO- = 2,0 et que
pKa NH3+ / NH2 = 9,2
Page | 7
CPCM- UE1B- FC12.2- Les propriétés spectrophotométriques
On retiendra que les acides -aminés aromatiques (Phe, Tyr, Trp) absorbent dans
l’U.V. à max = 280nm (Phe plus précisément à 260 nm). On considèrera que les autres
acides -aminés n’absorbent pas dans l’U.V. Les acides aminés aromatiques augmentent
fortement l’absorption des protéines dans l’U.V. Ils permettent le dosage des protéines selon
la loi de Beer- Lambert DO = lC.
3) Les modifications post-traductionnelles des acides aminés dans les
protéines
De nombreuses modifications post-traductionnelles sont réalisables sur les acides
aminés dans les protéines. Seules les plus importantes figurent dans le tableau ci-dessous.
Les principales modifications post-traductionnelles des acides aminés
Ponts
Phosphorylation Acétylation Hydroxylation Méthylation Glycosylation
disulfures
Sérine OUI OUI
Thréonine OUI OUI
Tyrosine OUI
OUI OUI OUI
Lysine
(Ex : Histones) (Ex : Collagènes) (Ex : Histones)
Histidine OUI
OUI
Proline
(Ex : Collagènes)
Asparagine OUI
Cystéine OUI
L’extrémité NH2 terminale des acides aminés peut être protégée par acétylation
L’acide aspartique et l’acide glutamique peuvent subir des réactions de carboxylation
La cystéine est le seul acide aminé capable de réaliser des ponts disulfures (les ponts disulfures
s’établissent entre deux cystéines cystine)
Page | 8
CPCM- UE1B- FC14) Rôles des acides aminés
Structural : leur enchaînement linéaire confère la structure primaire de la protéine
Fonctionnel :
o GABA : neurotransmetteur
o Précurseurs d’hormones, de neurotrznsmetteurs, d’acides nucléiques, etc…
(Tyr DOPA)
o Métabolique :
Transportent et éliminent l’azote (cycle de l’urée)
Substrats énergétiques : production d’ATP (- Une liaison forte (covalente), polaire, non chargée, sans propriété acido-basique (ne
s’ionise pas dans l’eau, ne se protone pas dans l’eau)
- Très résistante à l’hydrolyse chimique (voir méthodes)
- Une liaison plane : la stabilisation par résonance confère à la liaison peptidique un
caractère partiel de liaison double. Ainsi la liaison C–N a une longueur intermédiaire
entre liaison simple et liaison double
- De configuration trans privilégiée car plus stable (cis possible, surtout dans le cas de
la proline mais reste minoritaire) : en effet, le caractère liaison double partielle
entraîne un empêchement de rotation autour de la liaison C-N.
- Capable d’établir des liaisons hydrogène (CO accepteur, NH donneur)
3) Les différents niveaux de structure des protéines (ou des peptides)
A partir du gène, on obtient la séquence primaire de la protéine puis, par intervention parfois
de chaperons, le reploiement de la protéine. Les différentes structures indiquent l’organisation
respective des acides aminés.
3.1- La structure primaire des protéines
C’est l’enchaînement simple des acides aminés (qui composent le peptide ou la
protéine) les uns à la suite des autres par liaisons peptidiques. La structure primaire est
génétiquement déterminée. Elle conditionne les structures secondaires et tertiaires des
protéines. Une protéine de structure primaire n’a généralement pas d’activité biologique.
3.2- Les structures secondaires des protéines
Cette structure définit la position relative de deux acides aminés contigus ; si cette position est
répétitive, on obtient des structures régulières en hélice ou en feuillet.
Les propriétés physico-chimiques de la liaison peptidique impliquent des arrangements
spatiaux particuliers pour la chaîne polypeptidique (voir schéma ci-dessous).
En effet, le caractère partiel de double liaison empêche la libre rotation autour de la liaison
peptidique C-N. On peut donc définir la position relative des restes R de deux acides aminés
consécutifs par les angles de torsion , tels que :
- définit la rotation autour de la liaison C – C
- définit la « rotation » autour de la liaison peptidique CO – N : 180° trans ou 0° cis
(les atomes C, O, N et H formant la liaison peptidique étant coplanaires)
- définit la rotation autour de la liaison N – C2.
Page | 10
CPCM- UE1B- FC1Une chaîne polypeptidique va ainsi s’organiser en segments dans lesquels un couple d’angle
(,) peut se répéter, définissant ainsi une structure secondaire régulière (si le couple se
répète) ou irrégulière.
Seul un nombre limité de couples (,) est permis, de par la taille des différents restes R des
acides aminés qui composent la chaîne polypeptidique.
Ces couples sont traduits graphiquement par les diagrammes de Ramachandran et
correspondent aux interactions minimales entre ces restes R
Chaîne polypeptidique : mise en évidence des angles de torsion ,
Page | 11
CPCM- UE1B- FC1Diagramme de Ramachandran
3.2.1- Les structures secondaires régulières de type hélice
Il existe de nombreuses structures en hélice, les caractéristiques des principales étant décrites
dans le tableau ci-dessous. Les hélices étant les plus communément rencontrées, elles sont
décrites par la suite plus en détail.
Hélice Hélice 310 Hélice de collagène
(//) (-57/-47/180) (-49/-26/180) (-80 à -57 / 130 à /155/180)
Pas de l’hélice 1,5Å / 100° 2,0Å 3,1 Å
Résidus / tour 3,6 3,0 3
Sens de rotation Droite (gauche Droite Gauche (triple hélice droite)
rare)
Liaisons H Entre n et n+4 Entre n et n+3
Les principaux types d’hélice dans les protéines
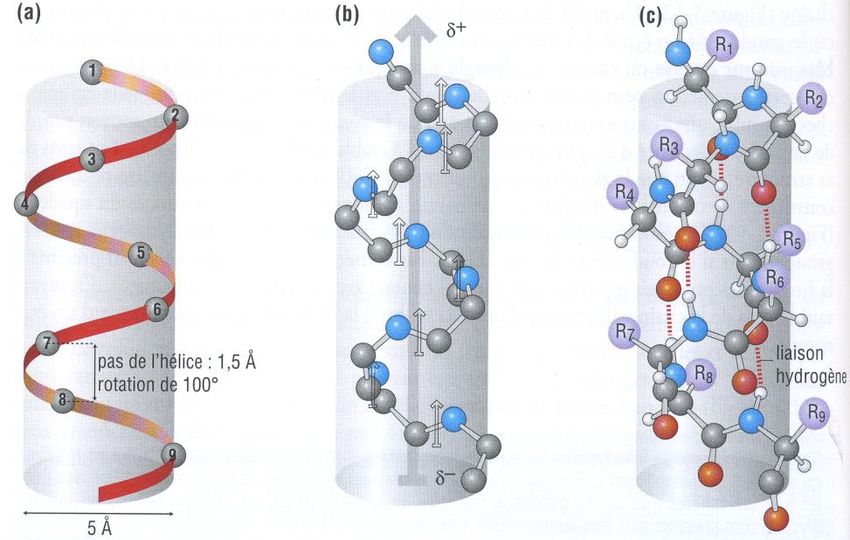
3.2.1.1- L’hélice
- Les carbones des acides aminés forment le squelette de l’hélice (schéma (a)),
consistant en une hélice droite le plus souvent (s’enroule dans le sens des aiguilles
d’une montre)
Page | 12
CPCM- UE1B- FC1- L’hélice est stabilisée par liaisons hydrogène (liaisons faibles) entre les C=O et les
NH des liaisons peptidiques (NH de l’acide aminé n° i et CO de l’acide aminé n° i +
4, schéma (c))
- Les liaisons hydrogènes sont toutes orientées dans le même sens, parallèles à l’axe de
l’hélice (formation de dipôles peptidiques alignés, schéma (b))
- Les radicaux R des acides aminés pointent vers l’extérieur de l’hélice avec un
décalage de 100°
- La proline est interdite dans l’hélice car elle ne peut pas engager sa liaison
peptidique (amide tertiaire donc pas de H) dans une liaison hydrogène
- La glycine est interdite dans l’hélice car son radical R ne peut satisfaire les
conditions (,) imposées par la structure
- Des AA de mêmes charges ou trop volumineux en i et i+4 défavorisent la structure
- Une hélice peut être totalement hydrophobe (ne comporte que des acides aminés
hydrophobes) : les hélices forment fréquemment les passages transmembranaires
des protéines
- Une hélice peut être totalement hydrophile (ne comporte que des acides aminés
hydrophiles)
- Une hélice peut être amphiphile : une face hydrophile et une face hydrophobe : un
ensemble d’hélices amphiphiles forment fréquemment les pores des canaux
ioniques de telle façon que les faces hydrophobes sont au contact de la bicouche
lipidique et les faces hydrophiles forment l’intérieur du canal
Hélice : squelette (a), dipôle (b), liaisons hydrogène (c)
3.2.1.2- Caractérisation d’une hélice : méthode de la roue hélicoïdale
Cette technique de représentation permet la mise en évidence du caractère hydrophobe,
hydrophile ou amphiphile d’une hélice (voir schéma ci-dessous). Dans cette méthode,
chaque résidu d’acide aminé est projeté sur un cercle dans un plan perpendiculaire à l’axe de
l’hélice. On obtient la roue en se rappelant que les restes R de deux acides aminés consécutifs
sont décalés d’un angle de 100°.
Page | 13
CPCM- UE1B- FC1Lorsque la roue est tracée et que vous devez disposer correctement les acides aminés dessus,
l’acide aminé n°2 est à 5 points du n°1, le n°3 est à 5 points du n°2…(voir le schéma ci-
dessous).
Hélice amphiphile (= amphipathique) : les acides aminés hydrophobes sont groupés du côté
droit de la projection, les acides aminés hydrophiles sont du côté gauche
3.2.1.3- L’hélice de collagène
- Riche en prolines et en glycines : Répétition de triplets motif de séquence (Gly – X –
Y)n X et Y étant des acides aminés quelconques (souvent proline pour X ou hydroxy-
proline / hydroxy-lysine pour Y)
- Riche en acides aminés atypiques 3-hydroxy-proline, 4-hydroxy-proline, 5-
hydroxy-lysine obtenus par modifications post-traductionnelles
- Hélice gauche : Association de 3 hélices gauches en décalage de 1/3 pour former une
triple hélice droite = tropocollagène
- Stabilisation de l’hélice gauche par liaisons hydrophobes : la proline réalisant des
liaisons peptidiques amides tertiaires, il n’y a pas de H disponible pour réaliser des
liaisons hydrogène. De plus, l’encombrement du cycle proline interdit les couples
(,) favorables à l’hélice
- Stabilisation de la triple hélice droite par :
- Liaisons hydrogènes inter-chaînes entre les C=O et les NH des
liaisons peptidiques (Pro- Gly ; Lys- Gly ; HO-Pro- Gly) (pas de
liaisons hydrogène intra- caténaires !)
- Liaisons hydrogènes inter-chaînes mettant en jeu les OH des hydroxy-
prolines
- Liaisons hydrophobes
- Ponts disulfures aux extrémités non torsadées
1. Synthétisée en intracellulaire sous forme de procollagène : 2 chaînes alpha se
dimérisent via un domaine particulier, puis arrivée de la 3 ème chaîne ; mise en place de
ponts disulfures dans 2 domaines du trimère puis association des 3 hélices
procollagène
2. Clivage des extrémités Nter et Cter tropocollagène
3. Sécrétion
4. Tropocollagène assemblé en extracellulaire en fibres de collagène tissu- spécifiques
Les 3 hélices peuvent être identiques ou différentes ; 1 gène ancestral donne naissance aux 3
chaînes ; 28 combinaisons possibles de chaînes
1 exon du gène code en moyenne pour 6 motifs (Gly- X- Y) et 50 exons en moyenne par gène
Page | 14
CPCM- UE1B- FC13.2.2- Les structures secondaires régulières de type feuillet
Ce sont des feuillets vrillés, flexibles et inextensibles, pouvant s’organiser en cylindre.
Feuillet antiparallèle Feuillet parallèle
Association de brins par liaisons hydrogènes entre les NH et les C=O des liaisons
Formation et stabilisation peptidiques orientés vers l’intérieur du feuillet. Les séquences d’acides aminés formant
les brins sont parfois très éloignées dans une même chaîne polypeptidique
Connexion entre deux brins
Coude le plus souvent Boucles
Conformation des liaisons
Trans
peptidiques
Distance entre deux résidus 3,3Å
Acides aminés interdits proline et glycine
Les liaisons hydrogène sont bien
Les liaisons hydrogène ne sont pas
Particularité des liaisons perpendiculaires au plan du feuillet,
parfaitement perpendiculaires au plan du
hydrogène - Conséquence ce qui confère une plus grande
feuillet
stabilité
De par sa grande stabilité, est
fréquemment hydrophile ou
Caractère hydrophile / Généralement hydrophobes, enfouis dans la
amphiphile (alternance de résidus
hydrophobe / amphiphile structure protéique
hydrophobes / hydrophiles) donc en
surface des protéines
Les liaisons peptidiques non engagées dans les liaisons hydrogène stabilisatrices du feuillet, ainsi que les restes R des
acides aminés sont orientés vers l’extérieur du feuillet (au dessus ou au dessous du plan) et peuvent réaliser des
contacts avec l’eau ou avec d’autres structures protéiques
Feuillet antiparallèle et parallèle – Existence de feuillet mixte
3.2.3- Les structures secondaires irrégulières de type coudes et boucles
Coude
Nombre d’acides aminés 3 à 4 acides aminés
Une liaison hydrogène :
- entre le C=O de l’acide aminé n° i et le NH de l’acide
Stabilisation aminé n° i + 3 : coude
- entre le C=O de l’acide aminé n° i et le NH de l’acide
aminé n° i + 2 : coude (moins fréquent)
Proline et Glycine
Acides aminés fondamentaux
La proline est fréquemment en position i + 1, jamais n°1,
Page | 15
CPCM- UE1B- FC1jamais n°4
La glycine est fréquemment en position i + 3
Entre n°1 et n°4 : Asn, Ser, Asp fréquemment
Permet un changement d’orientation d’une chaîne
Intérêt
polypeptidique. Ex : connexion de brins
Enfoui dans la structure protéique ou en surface au contact
Localisation
du solvant
Les boucles sont des structures de 4 à 12 acides aminés, fréquemment placées en surface des
protéines, au contact du solvant. Elles peuvent intervenir dans des phénomènes de reconnaissance,
de catalyse (fixation d’un substrat) ; ex : 1- anti- trypsine (1AT vue avec Barouki)
Le coude
3.3- Les structures super-secondaires (ou motifs structuraux) des protéines
Les structures super-secondaires sont des combinaisons simples de structures secondaires
fréquemment retrouvées dans les protéines, caractérisées par une architecture spécifique et
parfois par une fonction particulière (voir le tableau page 12).
4) La structure tertiaire
Il s’agit de la description de l’association des acides aminés éloignés en séquence
primaire et rapprochés par des boucles et des coudes. Il s’agit de la structure spatiale de la
protéine (disposition globale dans l’espace de l’ensemble des structures secondaires quelle
que soit leur nature). Toutes les protéines possèdent une structure tertiaire.
Les boucles ont un rôle de jonction entre segments ; elles sont très flexibles et peuvent
être impliquées dans des changements de conformation.
L’hydrophobicité des radicaux d’AA conditionne le reploiement : les AA hydrophobes
définissent ainsi des domaines transmembranaires ou enfouis au cœur de la protéine.
Selon leur structure tertiaire, les protéines peuvent être classées en deux catégories :
- les protéines globulaires : elles présentent généralement une activité enzymatique.
- les protéines fibreuses : ce sont généralement des protéines de structure (collagènes,
kératines, élastine…).
Page | 16
CPCM- UE1B- FC14.1- La stabilisation de la structure tertiaire
Liaison forte (covalente) : Liaisons faibles
Eventuellement pont(s)
disulfure(s) intra-chaîne ou inter-
chaînes polypeptidiques. On
rappelle qu’un pont disulfure ne
peut s’établir qu’entre deux
cystéines
Liaison de Van der Waals (3,5Å, 4kJ.mol-1) :
- Type dipôle permanent / dipôle permanent
- Type dipôle permanent / dipôle induit
- Type dipôle induit / dipôle induit
Les acides aminés neutres sont principalement engagés dans des interactions hydrogènes ; les
acides aminés chargés dans des interactions ioniques ; les acides aminés hydrophobes dans
des interactions hydrophobes.
– (motif en épingle à cheveux)
Rencontré dans de très nombreuses protéines. Peut se répéter
n fois ( – )n. Les hélices sont connectées par des coudes
de 2 à 4 acides aminés caractérisés par une glycine en
position n°2.
Les hélices sont disposées à 20° et établissent entre elles des
contacts hydrophobes
Hélice – tour – Hélice
Fréquemment rencontré dans les protéines facteurs de
transcription (liaison avec l’ADN). Dans ce cas les hélices
sont disposées à environ 120°
Page | 17
CPCM- UE1B- FC1Hélice –boucle – Hélice
Rencontré dans de nombreuses protéines. Ici, ce motif
permet la fixation de Ca2+ dans la troponine C (intervention
d’acides aminés acides Asp, Glu). Notez que Ca2+ se fixe sur
la boucle et pas sur les hélices
– (motif en épingle à cheveux)
Rencontré dans de très nombreuses protéines. Peut se répéter
n fois ( – )n Nombreuses combinaisons possibles
––
Deux brins parallèles (formant un feuillet) connectés par
une hélice pouvant se placer au dessus ou au dessous du
plan du feuillet. Ce motif peut se répéter ( – – )n
––
Deux brins antiparallèles (formant un feuillet) connectés à
une hélice .
Exemple : la structure doigt de zinc, rencontrée dans de
nombreuses protéines facteurs de transcription. Présence
caractéristique de cystéines et d’histidine pour la chélation
du zinc (Zn2+)
Les principales structures super-secondaires
4.2- Structure tertiaire et domaines
4.2.1. Définition d’un domaine
Un domaine correspond à une zone d’association compacte de structures secondaires et /
ou de superstructures secondaires. Il constitue une unité fondamentale structurale et
souvent fonctionnelle d’une structure tertiaire. Une structure tertiaire peut donc être
constituée de plusieurs domaines de structures et de fonctions différentes, reliés entre eux par
des structures irrégulières de chaîne polypeptidique. On admettra qu’il n’existe pas de
différence entre domaine et sous unité.
Une région de la protéine pouvant être fonctionnellement individualisée est un domaine
(domaine de fixation d’un ligand ; domaine de dimérisation ; domaine catalytique ; etc…)
Un domaine peut aussi être défini comme une région de la protéine présentant une homologie
de séquence avec une autre molécule.
Un domaine enfin peut être une unité de reploiement (domaine en feuillet ou domaine en
hélice).
Page | 18
CPCM- UE1B- FC14.2.2. Différents types de domaines – Exemples fondamentaux d’architecture
Type de domaine Exemple Architecture
Hormone de croissance Fagot d’hélices (4 hélices)
Domaines : hélices associées par contact Cytochrome b-562, Fagot d’hélices (4 hélices) pour de
hydrophobe (cœur hydrophobe), fréquemment Myohémiérythrine, nombreuses protéines à noyau hème
antiparallèle connectées par des boucles et Ferritine (fixation du fer)
disposées à 20°, 50°, 90°, ce qui facilite 8 hélices connectées par des boucles.
l’empaquetage de la structure Myoglobine
Fixe le fer par un noyau hème
Histones 4 hélices
Tonneau en méandres. Les résidus
Rétinol binding protein hydrophiles alternent avec les résidus
(RBP), hydrophobes, ce qui conduit à une face
Domaines : le plus souvent deux feuillets hydrophile et une face hydrophobe.
antiparallèles formant un sandwich ou un Porines (protéines
transmembranaires), Le tonneau est une alternative aux
feuillet replié en tonneau . On n’observe que hélices amphiphile pour réaliser des
Green fluorescent
des brins connectés par des coudes ou boucles. canaux transmembranaires
protein (GFP)
Notez que la distinction entre tonneau et
sandwich n’est pas claire, des systèmes
partiellement en tonneau pouvant se former dans -cristalline,
les structures sandwich (cas des Pré-albumine Tonneau à 2 motifs grecs
immunoglobulines) Super – oxyde dismutase Sandwich à motifs grecs. Les plans
(S.O.D), des feuillets se font face. Fixation de
Immunoglobuline ligand sur une boucle
Tonneau ( – (ou TIM) : cœur
hydrophobe formé d’un feuillet -
Domaines – : les deux motifs les plus Triose phosphate parallèle entouré d’hélices
fréquents sont : isomérase, nombreuses amphiphiles pour assurer le contact
- le tonneau – enzymes avec le solvant et avec le feuillet. Le
site actif est toujours en C-terminal et
- l’enroulement - (arrangement de
le substrat se fixe en partie sur les
Rossman)
boucles
Les feuillets sont parallèles ou mixtes (plus
rarement)
Enzymes fixant des Sandwich ouvert ( – – – )2 =
coenzymes arrangement de Rossman
nucléotidiques
(GADPH, flavodoxine
par exemple)
Domaines + : présence d’hélices et de Ubiquitine, hexokinase,
feuillets séparés. Architectures mélangées lysozyme
5) Structure 3D et chaperons
La structure 3D requière parfois l’intervention de chaperons pour se mettre en place. La
structure 3D est acquise progressivement pendant la traduction, et se mettent en place
séquentiellement :
1. les liaisons hydrophobes
2. les liaisons hydrogènes
3. les liaisons de Van Der Waals
4. les liaisons ioniques
5. les ponts disulfures
Page | 19
CPCM- UE1B- FC1On obtient ainsi la conformation dite native.
Il existe des chaperons de réparation de protéines endommagées ou mal reployées (ex :
Hsp60) et des chaperons permettant l’acquisition de la structure 3D native (ex : Hsp70), qui
empêchent les liaisons incorrectes.
Les Hsp70, liées aux ribosomes, sont ubiquitaires. Elles fixent la protéine hydrophobe en
étant phosphorylées (charge – qui interagit avec les charges + des acides aminés). L’hydrolyse
de l’ATP permet le changement de conformation de la protéine. Le chaperon est
déphosphorylé après reploiement de la protéine.
Hsp60, en forme de tonneau, lie la protéine abîmée et l’hydrolyse de l’ATP là- encore permet
le reploiement de la protéine. Si la réparation est impossible, la protéine sera dégradée. Hsp60
préviendra alors l’agrégation des protéines anormales.
Chez les Eucaryotes, il existe des chaperons spécifiques de protéine ou spécifiques de
compartiment (ex : calnexine et calréticuline du REG).
6) La structure quaternaire
5.1- Définition – Structure quaternaire – Monomère - Protomère
Une structure quaternaire correspond à l’association d’au moins deux chaînes
polypeptidiques identiques ou différentes par liaisons faibles. On dit aussi qu’une structure
quaternaire est composée de sous unités, chaque sous unité formant un monomère.
Généralement, les différentes sous unités ont tendance à adopter un arrangement
symétrique. Quand il existe des sous unités différentes, l’entité asymétrique à partir de
laquelle le complexe symétrique est construit est appelé « protomère ». Un exemple
classique est le protomère () de l’hémoglobine ()2. Le protomère correspond au plus
petit ensemble fonctionnel.
5.2- L’assemblage d’une structure quaternaire – Torsade d’hélices
Les sous unités s’assemblent par mise en contact optimisée de surface de liaisons
complémentaires. Par exemple une surface riche en acides aminés chargés négativement dans
la sous unité A pourra faire contact avec une surface riche en acides aminés chargés
positivement dans la sous unité B
Les structures en torsade d’hélices droites, dites encore super-hélices gauches ou
« coiled – coil », permettent l’assemblage de deux sous unités par complémentarité entre
surface d’interaction. Ce type de structure est toujours caractérisé par la répétition d’un motif
heptade, tel que l’on retrouve régulièrement le même acide aminé hydrophobe tous les 7
résidus. L’exemple type est la structure Leucine Zipper, rencontrée dans de nombreuses
protéines facteurs de transcription.
Page | 20
CPCM- UE1B- FC1Les caractéristiques de la structure en hélice « coiled – coil »
Nombre de résidus par tour d’hélice 3,5
Point d’émergence des chaînes latérales Tous les 2 tours donc tous les 7 acides aminés
Séquence caractéristique (Heptade)n = (a, b, c, d, e, f, g)n
Présence de leucine ou isoleucine tous les 7
acides aminés (position « d ») + un autre
Particularités de l’heptade acide amine hydrophobe en position « a ».
Ainsi, l’acide aminé « a » et l’acide aminé
« d » stabilisent la structure à tour de rôle
Exemple Leucine Zipper
Homo-dimérisation ou hétéro-dimérisation
Intérêt de la structure “leucine zipper » par association d’hélice de chaque
monomère
Exemples de protéines fibreuses à structure Myosine, tropomyosine, fibrine, kératine
« coiled coil » (grande résistance mécanique)
7) Pathologies de conformation des protéines
La maladie de Creutzfeld Jacob se caractérise par une dépression, puis une démence et la mort
de l’individu. Le cerveau présente une structure spongieuse avec des « vides » entre les
cellules. Il existe des formes familiales laissant penser à une anomalie génétique.
Une autre maladie, le Kuru, atteint dans une tribu de Nouvelle Guinée (peuple des Fore) les
femmes et les enfants, mais pas les hommes, ce qui laisse à penser à une transmission non
génétique. Il s’agit aussi d’une encéphalopathie spongiforme transmissible.
Dans cette tribu, les hommes mangent les muscles des défunts (rite mortuaire), tandis que
femmes et enfants consomment le cerveau.
L’agent infectieux a été dénommé « prion » et s’est avéré être une protéine mal conformée. La
protéine cellulaire saine, PrPc, contient 40% d’hélices alpha (3 hélices). La protéine
pathologique, PrPsc contient 30% d’hélices et 45% de feuillets : cette protéine mal conformée
induit un changement de conformation de la protéine saine lors de l’infection, c’est- à- dire se
comporte comme un chaperon.
Il existe donc des formes génétiques (mutation) et des formes transmises, acquises, pour
lesquelles la séquence protéique est normale mais la protéine présente une mauvaise
conformation.
Transmission : (barrières d’espèce) :
Ovin- humain : -
Ovin- bovin : +
Bovin- humain : +
Humain- hamster : +
Humain- rat : -
Page | 21
CPCM- UE1B- FC1Vous pouvez aussi lire

















































