Vers des pratiques de conservation durable : Évaluation de l'effet de l'ensemencement et de la remise à l'eau sur le succès reproducteur du saumon ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Vers des pratiques de conservation durable : Évaluation de l’effet de l’ensemencement et de la remise à l’eau sur le succès reproducteur du saumon atlantique (Salmo salar, L.) Mémoire Raphaël Bouchard Maîtrise en biologie - avec mémoire Maître ès sciences (M. Sc.) Québec, Canada © Raphaël Bouchard, 2021
Vers des pratiques de conservation durable : Évaluation de l’effet de l’ensemencement et de la remise à l’eau sur le succès reproducteur du saumon atlantique (Salmo salar, L.) Mémoire Raphaël Bouchard Sous la direction de : Louis Bernatchez, directeur ou directrice de recherche Julien April, codirecteur
Résumé L’ensemencement et la remise à l’eau sont parmi les outils de conservation les plus utilisés pour la gestion de la pêche récréative. Par contre, le rapport coût/bénéfice de ces deux pratiques n’est que très rarement évalué de façon exhaustive. Dans le cas du saumon atlantique, plusieurs d’études rapportent que les poissons ensemencés sont moins adaptés au milieu naturel, mais peu ont spécifiquement examinés quelles étaient les causes de la réduction de succès reproducteur ou ont comparé leur contribution à la diversité génétique à celle des tacons précoces. De plus, seulement une seule étude a documenté la capacité de reproduction des saumons remis à l’eau par les pêcheurs, alors que la fréquence des remises à l’eau est en constante augmentation en Atlantique Nord. Notamment, aucune étude n’a évalué si les saumons remis à l’eau peuvent se reproduire au-delà de températures de 20°C qui sont connues pour augmenter la mortalité chez les saumons graciés. Dans cette étude, nous documentons le patron de reproduction des poissons ensemencés ainsi que de ceux pêchés puis remis à l’eau en utilisant un pedigree moléculaire comprenant tous les parents d’une population sauvage de saumon atlantique (Salmo salar, L.). Nos résultats démontrent que les saumons ensemencés ont significativement moins de partenaires de reproduction que les saumons sauvages ce qui réduit leur succès reproducteur par rapport à leurs conspécifiques sauvages. La participation des saumons ensemencés à la reproduction a significativement augmenté la diversité génétique au sein des rejetons. Cependant, les tacons précoces du système l’on fait de façon plus importante et ont significativement augmenté le nombre d’allèles chez les rejetons. Bien que 83% des saumons remis à l’eau aient produit des rejetons, les femelles graciées avaient en moyenne 73% du succès reproducteur des saumons qui n’étaient pas pêchés. Les températures supérieures à 20°C n’étaient pas associées à une réduction de succès reproducteur chez les saumons remis à l’eau. Ceci suggère que la population étudiée est localement adaptée aux températures élevées ou que les saumons ont régulé leur température interne de façon comportementale en se déplaçant vers des refuges thermiques. Ces données permettront de raffiner les analyses de risques et de bénéfices associés à l’ensemencement et à la remise à l’eau et ainsi, d’optimiser les pratiques de conservation utilisées pour la gestion des populations de saumons atlantiques. ii
Abstract Captive-breeding programs as well as and catch-and-release are among the most commonly adopted conservation practices in recreational fisheries. However, risks and benefits associated with their joint implementation are rarely extensively evaluated. In the case of Atlantic Salmon, while previous studies revealed that captive-bred fish show reduced fitness compared to their wild counterparts in nature. Yet, few examined the extent and causes of their reduced reproductive success or directly compared their contribution to enhance genetic diversity to that of wild fish, including mature male parr. Furthermore, only one study specifically measured the reproductive success of caught and released Atlantic salmon in natural settings, and no study to date evaluated if released salmon are able to reproduce when released at temperature above 20°C which is known to increase post- release mortality. Here, we use high-throughput microsatellite sequencing of 38 loci to accurately assign 2500 offspring to a comprehensive set of possible parents from a supplemented Atlantic salmon population in Québec, Canada. The resolved molecular pedigree provided informative insight on the reproductive pattern of both captive-bred salmon and caught-and-released salmon. Captive-bred salmon had fewer partners and significantly reduced relative reproductive success compared to wild salmon. Supplementation of captive-bred salmon significantly contributed to increase genetic diversity but mature male parr did so to an even greater extent and significantly inflated the number of alleles found among offspring. Moreover, our results showed that that at least 83% of caught-and-released salmon did successfully reproduced although caught-and-released female salmon have a significantly reduced reproductive success, averaging 73% of the reproductive output of wild non-caught salmon. Reproductive success of released salmon was not influenced by water temperature over 20°C which suggests either that the studied population is locally adapted to warm waters or that they behaviorally regulated body temperature by accessing nearby thermal refugia. Our results should help refining managers’ ability to analyze the risks and benefits associated with captive- breeding and catch-and-release, and thus, optimize conservation practices used for the preservation of Atlantic salmon populations. iii
Table des matières Résumé ................................................................................................................................................................ii Abstract................................................................................................................................................................ iii Table des matières ..............................................................................................................................................iv Liste des figures...................................................................................................................................................vi Liste des tableaux .............................................................................................................................................. viii Liste des abréviations ..........................................................................................................................................ix Remerciements....................................................................................................................................................xi Avant-propos ..................................................................................................................................................... xiii Introduction .......................................................................................................................................................... 1 Exploitation et conservation de la ressource halieutique : une balance précaire ............................................ 1 La pêche récréative, ou le déclin invisible .................................................................................................. 1 Soutien anthropogénique des populations : l’ensemencement .................................................................. 2 La remise à l’eau comme outil de gestion et de conservation .................................................................... 4 L’ensemencement et la remise à l’eau en pratique : le cas du saumon atlantique (Salmo salar, L.) .............. 5 Présentation de l’espèce : L’Odyssée du saumon atlantique, roi des rivières ............................................ 5 Déclin des populations de saumon atlantique. ........................................................................................... 7 Conservation du saumon atlantique par l’ensemencement ........................................................................ 7 Efficacité de la remise à l’eau pour la gestion de la pêche au saumon ...................................................... 9 Gestion de la pêche au saumon dans un contexte de réchauffement climatique..................................... 12 Contexte du projet de recherche ................................................................................................................... 13 Chapitre 1: Effect of captive-breeding and catch-and-release on the reproductive success of Atlantic salmon (Salmo salar L.) ................................................................................................................................................. 15 1.1 Résumé ................................................................................................................................................... 15 1.2 Abstract ................................................................................................................................................... 16 1.3 Introduction.............................................................................................................................................. 17 1.4 Materials and Methods ............................................................................................................................ 20 1.4.1 Study area........................................................................................................................................ 20 1.4.2 Captive-breeding program of Rimouski River .................................................................................. 20 1.4.3 Atlantic Salmon angling fishery in the Rimouski River ..................................................................... 21 1.4.4 Sampling .......................................................................................................................................... 21 1.4.5 Molecular analyses .......................................................................................................................... 22 1.4.6 Parental allocation of fry .................................................................................................................. 23 iv
1.4.7 Detection of first generation (F0) migrant ......................................................................................... 23 1.4.8 Analysis of reproductive success of hatchery and C&R fish ............................................................ 24 1.4.9 Effect of captive-breeding on components of reproductive success ................................................ 25 1.4.10 Genetic estimates of the effect of captive-bred fish on the effective number of breeders (Nb) ..... 26 1.4.11 Estimate of the effect of mature male parr and captive-bred salmon on offspring’s genetic diversity .................................................................................................................................................................. 26 1.4.12 Effect of temperature and air exposure time on reproductive success of C&R salmon ................. 27 1.5 Results .................................................................................................................................................... 27 1.5.1 Adult population of Rimouski River .................................................................................................. 27 1.5.2 Caught-and-released salmon during 2018 angling season ............................................................. 28 1.5.3 Parental allocation ........................................................................................................................... 28 1.5.4 Description of the mating system ..................................................................................................... 29 1.5.5 Relative reproductive success (RRS) of captive-bred versus wild and C&R versus non-caught Atlantic salmon.......................................................................................................................................... 29 1.5.6 Effect of origin and male reproductive tactics on components of reproductive success .................. 31 1.5.8 Estimation of the effect of captive-bred anadromous salmon and of mature male parr on genetic diversity of offspring .................................................................................................................................. 36 1.5.9 Impact of C&R conditions on reproductive success ......................................................................... 37 1.6 Discussion ............................................................................................................................................... 38 1.6.1 Atlantic Salmon mating system in the Rimouski River ..................................................................... 39 1.6.2 Reduced fitness of captive-bred Atlantic Salmon after one generation of captivity ......................... 41 1.6.3 Contribution of captive-bred salmon and mature male parr to the effective number of breeders (Nb) and their influence on genetic diversity ..................................................................................................... 43 1.6.4 Effect of catch-and-release on the reproductive success of Atlantic Salmon .................................. 44 1.6.5 Evolutionary and practical consequences of captive-breeding and catch & release on wild populations................................................................................................................................................ 46 1.7 Acknowledgments ................................................................................................................................... 47 Conclusion ......................................................................................................................................................... 48 Conséquence de l’ensemencement sur le succès reproducteur : génétique et/ou épigénétique? ............... 48 Ensemencement, tacons précoces et diversité génétique ............................................................................ 49 La remise à l’eau comme outil de gestion ..................................................................................................... 50 Le futur de la pêche au saumon dans un contexte de réchauffement climatique ......................................... 51 Bibliographie ...................................................................................................................................................... 53 v
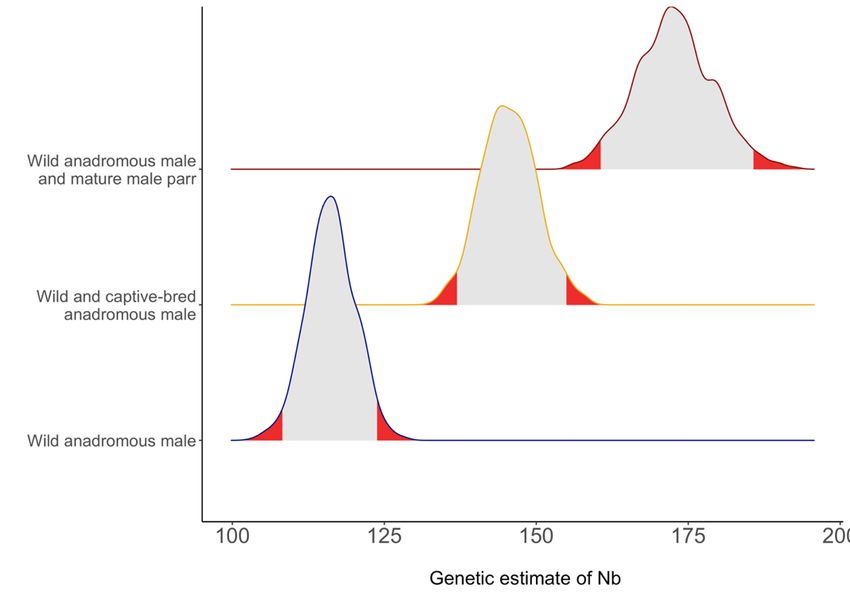
Liste des figures Figure 1: Maximum-likelihood estimates of relative reproductive success (RRS) and their associated 95% confidence intervals for captive-bred vs wild fish and caught-&-released vs non-caught Atlantic salmon (Salmo salar). If captive and wild or caught-&-released and non- caught salmon had equal fitness, then RRS would be equal to 1 (dashed red line). Filled and empty circles represent RRS that were significantly different at the 95% confidence interval given the permutation test. ........................................................................................................ 31 Figure 2: A) Relationship between the number of offspring (No. offspring) and mating success (No. mates) for female Atlantic salmon for three alternative reproductive tactics (Mature parr, 1SW and MSW). Coloured lines represent ZINB model prediction for captive- bred (blue) and wild (yellow) females, grey areas and hatched lines represent 95% confidence intervals (CI) obtained by bootstrap. Circles show individual data points. B) Relationship between mating success (No. mates) and length of female Atlantic salmon for three alternative reproductive tactics (Mature parr, 1SW and MSW). Coloured lines represent ZINB model prediction for captive-bred (blue) and wild (yellow) females, grey areas and hatched lines represent 95% confidence intervals (CI) obtained by bootstrap. Circles show individual data points........................................................................................................ 33 Figure 3: A) Relationship between number of offspring (No. offspring) and mating success (No. mates) for 1SW and MSW male Atlantic salmon. Lines represent ZINB model prediction for captive-bred (blue) and wild (yellow) 1SW and MSW males, grey areas represent 95% confidence intervals (CI) obtained by bootstrap. B) Relationship between mating success (No. mates) and winters at sea (Sea winters)x in male Atlantic salmon. Large circles with error bars represent the model prediction ± 95% confidence interval (CI) obtained by bootstrap while small circles show individual data points. Captive-bred males had significantly less mates than wild males (p < 0.038). ....................................................................... 35 Figure 4: Distribution of genetic (LDNe) estimates of effective number of breeders (Nb) considering either only wild anadromous salmon, both wild and captive-bred anadromous salmon or both wild anadromous salmon and mature male parr. The distribution of Nb calculated for wild anadromous salmon only was obtained directly from NeEstimator 2 (Do et al. 2014) whereas the distribution for wild and captive-bred anadromous salmon or wild anadromous salmon and mature male parr was obtained by subsampling 916 fry from the parent-fry assignment of wild and hatchery breeders 1000 times and calculating Nb on each subsampling step. The 2.5% and 97.5% tails of the distribution are indicated in red. .......... 36 Figure 5: Loess regression of the mean value of the number of alleles over all genotyped microsatellite loci calculated for 50 to 1500 fry fathered by wild anadromous male, wild and captive-bred anadromous male combined and all wild anadromous male and mature male parr. Each estimate was bootstraped 1000 times. Five to 95% interval distribution of the data were given around the mean value. ...................................................................................................... 37 vi
Figure 6: Reproductive success (estimated by number of fry assigned) for the group of caught-and-released Atlantic Salmon (n = 18) with increasing water temperature. .............. 38 vii
Liste des tableaux Table 1: Details of the genotyped adult and juveniles Atlantic salmon. The samples include all the returning adults caught at the Rimouski River dam from summer 2018 and the juveniles caught on the Rimouski River spawning grounds during spring 2019. .................... 28 Table 2: Relative reproductive success (RRS) of naturally spawning F1 parent. (nF1 is the sample size for naturally spawning captive-bred (C-B), wild (W), caught-and-released (C&R) and non-caught (N-C) parent; nF2 is the number of offspring assigned to each group of parent; RS is the reproductive success measured as the mean number of offspring assigned per parent; Variance is the average of the squared differences from the mean reproductive success; RRS is calculated as the RS of captive-bred fish over the RS of wild-origin fish and RS of C&R fish over the RS of non-caught fish, associated P-values are based on one-tailed permutation tests; Statistical power is the RRS value that would be significant with 80% and 95% probability). .......................................................................................................................................... 30 Table 3: Summaries of ZINB model testing the effect of number of mates, length and origin (wild/captive-bred) and the effect of length and origin on female’s reproductive success and mating success respectively. The “zero inflation” term accounts for the large number of adults with zero reproductive success and mating success in our sample. ............................. 32 Table 4: Summaries of ZINB model testing the effect of number of mates, sea age, and origin (wild/captive-bred) and the effect of sea age and origin on reproductive success and mating success respectively. The “zero inflation” term accounts for the large number of adults with zero reproductive success and mating success in our sample. ............................. 34 viii
Liste des abréviations RS: reproductive success RRS: relative reproductive success 1SW: one-sea winter salmon MSW: multi-sea winter salmon C&R: catch-and-release ix
À mon père x
Remerciements Quel univers que celui du saumon atlantique ! En acceptant le projet, je n’avais vraiment aucune idée du contexte sociopolitique dont fait l’objet ce nomade des eaux. Après seulement deux mois à travailler sur ma maîtrise, je passais déjà à Radio-Canada en tant qu’« expert » du saumon, ce que j’étais loin d’être à l’époque. Évidemment, le syndrome de l’imposteur cogne souvent à la porte dans ces moments, mais cela m’a amené à parfaire mes connaissances et à pousser mes capacités toujours un peu plus loin. Après près de trois ans de travail, je dois avouer être fier de l’expertise que j’ai acquise et des répercussions que ce projet aura sur la gestion du roi des rivières. Tout d’abord, merci Louis de m’avoir confié ce projet d’envergure. Je dois avouer avoir été honoré quand tu m’as accepté dans ton équipe, j’ai toujours admiré la qualité de la science qui sortait de ton laboratoire et je suis très content d’en faire partie aujourd’hui. Merci également de m’avoir fait grandir comme scientifique, c’est grâce à toi que j’ai compris que : la science, ça se fait en équipe. Finalement, merci de m’avoir offert ce poste au doctorat, je n’aurais pas pu rêver mieux. On s’embarque pour une grosse aventure, mais ces dernières années passées dans ton laboratoire m’ont bien préparé à relever tous les défis ! Mille mercis à toi Julien ! J’ai beaucoup apprécié ton encadrement tout au long de mon parcours et ta confiance en mes capacités. La réalisation du projet n’aurait jamais été possible sans ta planification exceptionnelle de l’échantillonnage. Bien que ça ne se soit pas toujours passé comme souhaité, je crois qu’on est arrivé à un produit fini très solide. Je te remercie également pour toutes les opportunités que tu m’as offertes pendant ma maîtrise : entrevues, conférences, revue de littérature sur la fraie du saumon, élaboration d’un protocole d’échantillonnage standardisé des nids de saumons, vulgarisation scientifique écrite… Tous ces accomplissements m’ont clairement donné une occasion de perfectionner mes connaissances et mes capacités à communiquer, tant à l’oral qu’à l’écrit. Merci à tous ces professeurs du département qui ont su partager leur passion et leur dévouement pour la biologie. Je pense notamment à Juan Carlos Villarreal avec qui j’ai pu publier mon premier article scientifique. À Julie Turgeon qui m’a confié la tâche difficile d’enseigner la théorie de l’évolution et la biologie moléculaire aux jeunes scientifiques du premier cycle pendant presque deux ans. À Christian Landry de m’avoir laissé travailler sur la levure pendant près de deux sessions et par la même occasion de parfaire mes connaissances en bioinformatique qui me serviront fort probablement pendant mon doctorat. Merci aux équipes du laboratoire Bernatchez et Moore qui ont contribué de près ou de loin au projet : Cécilia, Alysse, Éric, Bérénice, Anne-Laure, Claire, Maëva, Gabriella, Hugo, Martin, Clément, Florent, Isabeau, Jean- xi
Sébastien, Sara, Xavier… Deux équipes très motivantes qui unissent leurs forces pour préserver la faune aquatique ! Merci également aux colloques Sann et Louis-Philippe. Un an de pandémie, beaucoup trop de mois confinés, mais tout de même beaucoup de beaux moments partagés ! Maman, merci pour ton soutien moral et financier pendant toutes ces années. Éloïse Pelletier, merci d’avoir su rendre cette dernière année légère et remplie de bonheur malgré cette ambiance de fin du monde. xii
Avant-propos L’article intitulé « Effect of captive-breeding and catch-and-release on the reproductive success of Atlantic salmon (Salmo salar L.) » présenté dans ce mémoire sera soumis à la revue Ecological Applications. Les références de ce document sont présentées sous le format de l’American Psychological Association (APA), conformément aux consignes pour les auteurs de Evolutionary Applications. Je suis le premier auteur de l’article inséré au Chapitre 1 : j’ai participé au design de l’étude et j’ai réalisé l’acquisition, l’analyse et l’interprétation des données. Mes co-auteurs sont Kyle Wellband (aide à l’analyse des résultats), Laurie Leconpte (aide au laboratoire), ainsi que mon directeur Louis Bernatchez et mon co-directeur Julien April. Tous les co-auteurs ont contribué à la révision du manuscrit. xiii
Introduction Exploitation et conservation de la ressource halieutique : une balance précaire La pêche récréative, ou le déclin invisible Depuis le paléolithique (Yellen et al. 2015; Pringle, 1997), la pêche s’est inscrite comme pierre angulaire au sein des sociétés. Son ubiquité découle du fait qu’elle génère une ressource alimentaire et économique, tout en répondant à certains besoins sociaux n’étant pas reliés à la consommation (Arlinghaus, 2007). De ces besoins, on pense notamment au développement des techniques et outils de pêche, mais l’activité de pêche dite récréative favorise également la cohésion sociale et la transmission du savoir traditionnel (Acheson, 1981). En raison de son importance mondiale, l’exploitation des ressources halieutiques a connu une forte expansion lors du dernier siècle afin de répondre à une demande croissante (McIntyre et al. 2016). C’est cependant au cours des dernières décennies que son exploitation s’est intensifiée, avec une croissance annuelle moyenne se chiffrant à 3,1 % de 1961 à 2017, un taux deux fois plus élevé que l’augmentation démographique mondiale (1.6%) (FAO, 2020). Sans surprise, l’intensification de l’activité de pêche à des fins commerciales s’est accompagnée d’une pression croissante sur les populations exploitées (Pauly et al. 2002). En 2017, c’est 95% des pêcheries mondiales qui sont classées comme étant pleinement exploitées ou surexploitées (FAO, 2020). Les stocks de poissons exploités par la pêche récréative n’échappent pas à cette tendance (Post et al. 2002). Bien que les captures commerciales soient huit fois plus élevées que les captures récréatives en termes de biomasse, la pêche récréative égale et même supplante l’activité des pêcheries commerciales dans certaines localités (Cooke & Cowx, 2004). Ce phénomène s’observe notamment dans les pays développés des régions tempérées où une personne sur dix pêche pour le plaisir, cumulant ainsi 220 millions de pêcheurs récréatifs à l’échelle mondiale ce qui est cinq fois plus que le nombre de pêcheurs commerciaux (Arlinghaus et al. 2019). Par conséquent, bien que la pêche commerciale soit généralement identifiée comme l’acteur principal ayant mené au déclin des stocks de poissons (Pauly et al. 2002), la pêche récréative y participe également (Lewin et al. 2006). Présentement, deux mesures de gestion et de conservation existent afin de mitiger les impacts des activités de pêche récréative sur les espèces exploitées, soit l’ensemencement et la remise à l’eau. 1
Soutien anthropogénique des populations : l’ensemencement L’ensemencement de poissons est une mesure fréquemment utilisée en gestion et conservation afin d’assurer la pérennité des espèces aquatiques (Fraser, 2008). Typiquement, cette pratique consiste à introduire en milieu naturel des juvéniles produits en écloseries à partir de reproducteurs sauvages capturés en nature (Fraser, 2008). L’application de cette mesure ne date pas d’hier, en fait les premiers ensemencements auraient eu lieu au début du XVIIIe siècle et servaient uniquement à augmenter les opportunités de captures pour la pêche sportive (Pister, 2001). En ensemençant, nos ancêtres faisaient donc l’hypothèse intuitive qu’apporter de nouveaux poissons dans une rivière ou une étendue d’eau permettrait d’augmenter l’abondance d’une population. Cependant, les recherches scientifiques des dernières décennies ont démontré que la question est bien moins intuitive qu’il n’y parait (Araki et al. 2010). En fait, l’ensemencement permet parfois d’augmenter l’abondance d’une population, mais cela ne vient pas sans conséquence pour les individus sauvages. Mais pourquoi donc l’ensemencement ne fonctionne-t-il pas à tous les coups ? Reisenbichler & McIntyre furent parmi les premiers à se poser en 1977. Brièvement, l’équipe a généré des lignées des truites arc-en-ciel (Oncorhynchus mykiss) en élevage pendant quelques générations et ont comparé leur taux de survie à des truites sauvages lorsqu’introduites en milieu naturel. Leurs résultats étaient pour le moins frappants, en répétant cette expérience de jardin commun sur plusieurs tronçons de rivières, ils ont démontré que les truites ensemencées avaient en moyenne un taux de survie de 10% inférieur à leurs conspécifiques sauvages. Cela s’explique par le fait que le milieu de l’écloserie est bien souvent très différent du milieu naturel : la densité de poissons est plus élevée (Stingwell et al. 2014), la prédation et la compétition pour les ressources alimentaires sont inexistantes (Stingwell et al. 2014) et la physicochimie de l’eau diffère de celle de la rivière ensemencée (Lavoie et al. 2018) pour ne citer que quelques exemples. Bien que cela permette de réduire la mortalité et donc d’obtenir un nombre considérable de juvéniles pour l’ensemencement, ces différences contribuent à générer une progéniture qui ne développe pas une physiologie optimale pour le milieu naturel. En conséquence, les poissons ensemencés font état d’une réduction de valeur sélective et ont donc moins de chance de produire des descendants qui survivront jusqu’à maturité, et qui, à leur tour, se reproduiront transmettant ainsi leurs gènes aux générations futures (Christie et al. 2014). Les premières preuves empiriques de Reisenbichler & McIntyre (1977) ont donc lancé un débat actif dans 2
la communauté scientifique en concluant que « la sélection (ou l’absence de sélection) en écloserie génère des poissons qui sont mal adaptés au milieu sauvage ce qui pourrait avoir des conséquences écologiques et génétiques à long terme sur les populations ciblées ». Plus de 40 ans de recherche plus tard, les preuves démontrant que les poissons ensemencés ont une capacité de reproduction inférieure aux poissons sauvages s’accumulent. Sur 51 comparaisons entre poissons sauvages et ensemencés de quatre espèces de Salmonidés, 46 démontrent que les poissons ensemencés ont un succès reproducteur inférieur à leurs conspécifiques sauvages (Christie et al. 2014). Bien qu’utiliser des géniteurs provenant de lignées sauvages permette de réduire cette différence (Araki et al. 2007a), le succès reproducteur est toujours plus faible chez les poissons ensemencés ce qui démontre que la sélection de domestication en milieu d’élevage est très rapide (Christie et al. 2012). En effet, cette force sélective favorise les individus adaptés au milieu captif ; cependant, cela se produit au détriment de l’adaptation au milieu sauvage (Frankham, 2008 ; Christie et al. 2012). Comme l’article de Reisenbichler & McIntyre (1977) l’avait prédit, cette diminution de succès reproducteur chez les poissons ensemencés s’accompagne également de répercussions écologiques sur les populations ciblées (Ryman & Laikre, 1991; Hindar et al. 1991; Waples, 1991). En analysant 94 populations de trois espèces de saumons du Pacifique ciblées par l’ensemencement, Chilcote et al. (2011) ont démontré que la productivité d’une population (moyenne par adulte de rejetons qui contribuent à la reproduction) diminue avec une augmentation de la proportion d’adultes ensemencés. Pour ne citer qu’un cas explicite, les années où l’ensemencement a permis de doubler le nombre d’adultes, la productivité a été réduite de moitié. Ainsi, le bénéfice démographique immédiat de l’ensemencement peut-être complètement annulé par la perte de productivité. Face à ces constats, il est légitime de se demander pourquoi cette pratique est toujours utilisée. Dans certains cas extrêmes où la survie d’une population ou d’une espèce est en danger, l’ensemencement est une des solutions qui nous permette de prévenir l’extinction (Fraser et al. 2008). En effet, les populations caractérisées par un effectif réel très faible ou en déclin ont, la plupart du temps, un taux de consanguinité qui est conséquemment élevé (Frankham, 2005). La consanguinité aura pour effet de réduire la valeur sélective de la population par le biais d’une dépression de consanguinité (Tallmon et al. 2004). La population est alors entraînée dans un vortex d’extinction qui est causé par l’augmentation en fréquence de l’état homozygote d’allèles récessives délétères (Tallmon et al. 2004). Dans 3
ces petites populations, la taille efficace (effectif d’une population idéale qui aurait le même taux de perte d’hétérozygotie que l’effectif réel de la population) est également d’un ordre de grandeur plus petit que la taille réelle de la population (Frankham, 1995). Cette réduction de la taille efficace contribue notamment à la perte de diversité génétique par dérive génétique au sein de la population ce qui a des répercussions négatives sur son potentiel évolutif (Frankham, 2005). En effet, la diversité génétique est en fait le substrat sur lequel la sélection naturelle agit et permet aux populations de répondre adaptativement face aux changements de l’environnement (Toro & Caballero, 2005 ; Layton et al. 2021). Ainsi, le maintien de la diversité génétique est important, non seulement pour la conservation, mais aussi pour la gestion des pêcheries puisque la perte de diversité génétique peut altérer le maintien et la résilience d’une espèce et même causer son épuisement (Frankham, 2005). Dans ces situations, l’ensemencement a donc pour objectif de réduire la consanguinité et de préserver la diversité génétique en apportant de nouveaux individus dans la population (Fraser, 2008). La remise à l’eau comme outil de gestion et de conservation Offrant une alternative à l’ensemencement, la remise à l’eau des poissons capturés est une approche de gestion en plein essor qui vise principalement à réduire l’impact de la pêche récréative sur les populations de poissons (Policansky, 2002). L’implémentation de cette mesure permet notamment de maintenir l’activité socio-économique que génère ce secteur et sensibilise les pêcheurs à la conservation des espèces qu’ils exploitent (Arlinghaus et al. 2007). La remise à l’eau en tant qu’outil de gestion peut être mise en place selon différents cas de figures. Par exemple, lorsqu’un poisson est considéré de trop grande taille pour être conservé ou lorsqu’un pêcheur continue l’activité de pêche alors que son quota a déjà été atteint. Alors qu’elle est obligatoire dans certains cas, bien des pêcheurs remettent aujourd’hui leur poisson à l’eau sur une base volontaire, afin de répondre à des principes éthiques (Arlinghaus et al. 2007). À l’échelle mondiale, les pêcheurs remettent 60% de leurs prises à l’eau alors qu’une proportion considérable aurait pu être conservée légalement (Cook & Cowx, 2004). Néanmoins, l'efficacité de la remise à l'eau comme outil de gestion et de conservation repose sur la prémisse voulant que le poisson capturé puis relâché survive à cet événement et se reproduise aussi bien qu'un poisson non pêché (Arlinghaus et al. 2007). Auparavant, les gestionnaires de la ressource estimaient que la mortalité des poissons graciés était 4
relativement négligeable par rapport à la mortalité des poissons capturés qui est, quant à elle, inévitable (Muoneke & Childress, 1994 ; Bartholomew & Bohnsack 2005). Toutefois, les suivis télémétriques des poissons remis à l’eau ont permis de démontrer que le taux de mortalité est hautement variable, allant de 0 à 95% selon l’espèce étudiée et les conditions de remise à l’eau (Bartholomew & Bohnsack, 2005). Ainsi, l’application de cette pratique en tant qu’outil de gestion nécessite conjointement d’estimer le taux de mortalité des poissons graciés puisque ce paramètre peut affecter directement la démographie des populations exploitées. La capacité de reproduction des poissons remis à l’eau, bien qu’elle soit théoriquement d’importance égale à leur survie, n’est que rarement examinée (Cooke et al. 2002a). En effet, l’analyse du succès reproducteur des poissons est coûteuse et laborieuse ce qui rend son application difficile, voire impossible chez certaines espèces (Cooke et al. 2002a). Par mesure de précaution, l’activité de pêche est généralement limitée ou interdite pendant la période de reproduction afin de minimiser ses impacts potentiels sur le succès reproducteur d’une espèce donnée (Arlinghaus et al. 2007). L’ensemencement et la remise à l’eau en pratique : le cas du saumon atlantique (Salmo salar, L.) Présentation de l’espèce : L’Odyssée du saumon atlantique, roi des rivières Le saumon atlantique (Salmo salar L., salmonidae) est natif des régions tempérées et subarctiques de l’océan Atlantique Nord (Aas et al. 2010). Comme plusieurs espèces du clade des salmonidés, il est anadrome et se reproduit en eau douce et migre en eau salée pour rejoindre ses aires d’alimentation. Une partie de la population peut compléter l’entièreté de son cycle de vie en milieu dulcicole (résident) alors que d’autres entreprennent une migration en mer (anadrome). Cette migration nécessite une succession complexe d’acclimatations physiologiques, comportementales et morphologiques chez le saumon anadrome (McCormick, 2009); ce processus les préparant principalement à la vie en eau salée (smoltification). Suivant la smoltification, les saumoneaux entreprennent une migration printanière vers la mer, la dévalaison, qui est partiellement déclenchée par une augmentation de la température de l’eau (Byrne et al. 2003). Après une période de croissance rapide en mer, les adultes anadromes retournent vers leur rivière d’origine pour se reproduire après avoir séjourné typiquement 1 à 4 hivers en mer à s’alimenter (Webb et al. 2007 ; Fleming, 1996; Klemetsen et al. 2003). 5
Lors de la saison de reproduction, les femelles creusent le gravier et déposent leurs œufs dans un ou plusieurs nids qui forment une structure composite appelée frayère (Klemetsen et al. 2003). Pendant la fraie, les mâles anadromes établissent une hiérarchie basée sur la dominance pour avoir accès aux femelles (Fleming, 1996). Trois tactiques alternatives de reproduction existent chez les mâles, et celles-ci impliquent différents compromis de traits d’histoire de vie (Dodson et al. 2013). Les rédibermarins, qui séjournent plus d’un hiver en mer, sont les plus dominants sur les sites de fraies puisque les années passées en mer à s’alimenter leur permettent d’atteindre de plus grandes tailles que les madeleineaux (>63 cm vs < 63 cm) qui ne passent qu’un seul hiver en mer (Garant et al. 2003). Afin d’accéder aux femelles, les rédibermarins s’engagent dans une suite de combats où se sont généralement les plus imposants qui sortent victorieux (Fleming, 1996). Les madeleineaux sont, quant à eux, subordonnés sur les sites de fraies et occupent des positions satellites; ce faisant, leur succès reproducteur est généralement plus faible que celui des rédibermarins (Garant et al. 2003). Au sein d’une population donnée, certains mâles n’entreprennent pas de migration en mer et matures sexuellement en rivière. Ces mâles, appelés tacons précoces, ne combattent pas les mâles anadromes pour l’accès aux femelles, mais fertilisent plutôt les œufs de manière opportuniste en se faufilant au sein des nids (Fleming, 1996; Aubin-Horth & Dodson, 2004). Ils ont un succès reproducteur moins élevé que les mâles anadromes, mais ont de meilleures chances de survivre jusqu’à la reproduction (Taggart et al. 2001; Weir et al. 2010; Richard et al. 2012). Lorsque les montaisons de mâles anadromes sont petites, la participation des tacons précoces à la reproduction permet notamment de limiter la diminution de l’effectif efficace, la perte de diversité génétique et l’augmentation de la consanguinité chez les rejetons (Johnstone et al. 2013 ; Perrier et al. 2014). Après la fertilisation, les œufs éclosent au printemps et les alevins nouvellement formés utilisent les réserves contenues dans leur sac vitellin pendant 3 à 8 semaines avant leur émergence du gravier (Klemetsen et al. 2003). À la suite de l’émergence, dépendamment de la latitude, des conditions environnementales et de la génétique des individus, les tacons demeurent en eau douce pour une période variant de 1 à 7 ans (Thorstad et al. 2007), avant d’entreprendre leur migration en mer, si migration il y a. 6
Déclin des populations de saumon atlantique. Le saumon atlantique est pêché depuis la colonisation des côtes Européenne de l’Atlantique Nord comme en témoignent les ossements datés de 40 000 ans retrouvés en Espagne (Consuegra et al. 2002) et les peintures rupestres en Norvège et en Suède datant d’au moins 6000 ans (Webb et al. 2007). Aujourd’hui, les adultes anadromes sont d’une importance considérable du point de vue économique et culturel pour les toutes les nations établies en Atlantique Nord (ASF 2011). Cependant, l’espèce est en sérieux péril, et ce, sur toute son aire de répartition. Le conseil international de l’exploration de la mer (ICES) a documenté un déclin drastique des populations de saumon atlantique au cours dernières décennies (Chaput, 2012; ICES 2019). De nombreux facteurs ont synergétiquement contribué à la réduction de l’abondance des populations, notamment une surexploitation des stocks par la pêche commerciale et la construction d’ouvrages hydro-électriques (Limburg & Waldman, 2009; Otero et al. 2011). Ces impacts anthropiques ont réduit l’aire de répartition de l’espèce aux États-Unis (Spidle et al. 2001) et réduit l’abondance des populations au sud du Canada (COSEWIC, 2011). Depuis approximativement 1990, le déclin semble résulter principalement d’une augmentation de la mortalité en mer, et ce, malgré le fait que la pression de pêche en mer n’ait jamais été aussi faible depuis 1960 (Friedland et al. 2009; Chaput, 2012; ICES, 2019). Néanmoins, l’espèce demeure fortement prisée par l’industrie de la pêche sportive qui cible les adultes revenant en rivière pour se reproduire. De fait, la pêche au saumon génère annuellement plus de 102,1 millions de dollars en retombées économiques directes dans l’est du Canada (POC, 2008; ASF, 2011). Ainsi, afin d’assurer la durabilité de ce secteur au Québec, le Ministère de la Forêt, de la Faune et des Parcs (MFFP) a conjointement implanté l’ensemencement et la remise à l’eau comme pratiques de gestion et de conservation (MFFP, 2016). Conservation du saumon atlantique par l’ensemencement Si l’ensemencement était auparavant utilisé afin d’augmenter l’opportunité de pêche, la situation aujourd’hui est bien différente. Au Québec, son application est maintenant restreinte aux populations dont l’abondance est critique ou préoccupante et qui nécessitent une intervention afin d’éviter leur disparition (MFFP, 2016). L’encadrement de l’ensemencement se fait donc sous des conditions strictes, seulement cinq rivières sont en ce moment ciblées par un programme d’ensemencement multi annuel (MFFP, 2020). Par contre, le rapport coût/bénéfice de ces ensemencements n’est que très rarement évalué de 7
façon exhaustive, autant ici qu’ailleurs. Il est cependant crucial de connaître ses conséquences potentielles afin de pouvoir mitiger ses impacts sur les populations ciblées. L’environnement d’élevage induit de nombreuses modifications phénotypiques chez les saumons ensemencés. Par exemple, les saumons élevés en captivité sont plus agressifs que les saumons sauvages (Blanchet et al. 2008) ce qui réduit la capacité des saumons captifs à obtenir un partenaire pour la reproduction (Fleming et al. 1997). Les saumons nés en pisciculture démontrent également souvent des différences morphologiques par rapport à leurs congénères sauvages, soit des nageoires érodées et asymétriques, un corps et une tête moins fuselés et une nageoire caudale moins épaisse (Blanchet et al., 2008 ; Stingwell et al. 2014). Ces différences phénotypiques contribuent notamment à augmenter la mortalité des saumons ensemencés qui est jusqu’à cinq fois plus élevée que celle des saumons sauvages (Piggins & Mills, 1995). Les différences phénotypiques entre saumons sauvages et ensemencés ne peuvent pas toujours s’expliquer par une différentiation génétique (Christie et al. 2016; Le Luyer et al. 2017 ; Gavery et al. 2018 ; Rodriguez Barreto et al. 2019) bien que leurs profils d’expression de gènes soient foncièrement différents (Christie et al. 2016). Ainsi, les poissons ensemencés démontrent plus souvent des différences épigénétiques par rapport à leurs conspécifiques sauvages (Le Luyer et al. 2017 ; Gavery et al. 2018 ; Rodriguez Barreto et al. 2019). Contrairement aux différences génétiques, l’épigénétique réfère à des changements de régulation de gènes n’étant pas attribuables à des changements dans la séquence de l’ADN (Youngson & Whitelaw, 2008). Par exemple, Le Luyer et al (2017) ont démontré que, chez le saumon coho (Oncorhynchus kitsutch), c’est une centaine de régions dans le génome qui sont différentiellement méthylées entre les saumons ensemencés et sauvages. Plusieurs des locus hyperméthylés chez les saumons ensemencés, marque épigénétique typiquement associée à l’inactivation des gènes, étaient impliqués dans certaines fonctions essentielles telles que la locomotion et l’homéostasie. Ces résultats suggèrent donc que les marques épigénétiques induites par l’environnement d’élevage sous-tendent potentiellement la réduction de valeur sélective des saumons ensemencés. Comme chez d’autres espèces de saumons du Pacifique (Chilcote et al. 2011), augmenter la proportion de saumons ensemencés dans une rivière donnée contribue à diminuer la productivité d’une population ce qui est notamment du à un succès reproducteur inférieur à 8
celui de leurs conspécifiques sauvages (O’Sullivan et al. 2020). Bien que ces résultats démontrent que la capacité de reproduction des saumons ensemencés soit une composante essentielle du succès des programmes d’ensemencement, elle n’a été évaluée qu’une seule fois au Québec (Milot et al. 2013). L’étude de Milot et al. (2013) a été menée sur la population de la rivière Malbaie dans Charlevoix de 2002 à 2004. À cette époque, deux types d’ensemencement étaient pratiqués: l’ensemencement de tacons et l’ensemencement de saumoneaux. Leurs résultats démontraient pour la première fois que les saumons ensemencés au stade de tacon et de saumoneau avaient respectivement 70% et 40% du succès reproducteur des saumons sauvages. La réduction du succès reproducteur était donc plus accentuée chez les saumons ensemencés au stade de saumoneau qui passent plus de temps en captivité que les tacons. Cette étude mettait en évidence deux points majeurs: une seule génération en captivité peut réduire significativement le succès reproducteur des saumons ensemencés et que cet effet était d’autant plus important lorsque l’on augmentait le temps passé en captivité. Face à ces résultats, la recommandation de Milot et al. (2012) était d’ensemencer au stade d’alevin afin de limiter le temps passé en captivité. Aujourd’hui, plusieurs mesures sont mises en place par le MFFP afin de minimiser les impacts de l’ensemencement sur les populations ciblées (Bernatchez, 2009). Pour ne citer que quelques exemples, les géniteurs proviennent de la même rivière où les ensemencements ont lieu afin de maintenir l’intégrité génétique des populations (Araki et al. 2007b), le nombre de géniteurs est maintenu à 40 (20 mâles, 20 femelles) pour réduire la perte de diversité génétique en captivité (Frankham et al. 2002) et l’ensemencement est limité à 60 000 rejetons par année afin de minimiser les impacts sur la productivité de la population ciblée (Ryman & Laikre, 1991). À la suite de l’étude sur la rivière Malbaie, le MFFP a également décidé d’ensemencer le saumon au stade d’alevin afin de minimiser le temps passé en captivité. Cependant, aucune étude n’a tenté d’évaluer le succès reproducteur des saumons ensemencés à ce stade. Efficacité de la remise à l’eau pour la gestion de la pêche au saumon Contrairement à l’ensemencement, la pratique de remise à l’eau a grandi en popularité en tant qu’outil de gestion, non seulement au Québec où plus de 55% des saumons atlantiques capturés sont remis à l’eau (MFFP, 2016), mais dans tous les pays qui exploitent cette ressource (ICES, 2019). Seulement en 2019, c’est près de 166 000 saumons qui sont remis 9
Vous pouvez aussi lire

















































