VIII-1. ELECTROPHYSIOLOGIE VETERINAIRE
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Œil et Physiologie de la Vision - VIII-1
VIII-1. ELECTROPHYSIOLOGIE VETERINAIRE
Serge Georges Rosolen
Pour citer ce document
Serge Georges Rosolen, «VIII-1 : ELECTROPHYSIOLOGIE VETERINAIRE», Oeil et
physiologie de la vision [En ligne], VIII-La vision et son exploration chez l'animal, mis à
jour le 18/06/2013, URL :
http://lodel.irevues.inist.fr/oeiletphysiologiedelavision/index.php?id=213,
doi:10.4267/oeiletphysiologiedelavision.213
Plan
Introduction
Intérêt de l’ERG en médecine vétérinaire
But du chapitre
Physiologie animale comparée
La rétine
Généralités
Quelques particularités des rétines animales
Les voies visuelles
Description anatomofonctionnelle
Exploration rétinienne par ERG
Les conditions de genèse de l’ERG flash
Le choix du stimulateur et de la stimulation
Les conditions de stimulation
Les conditions de recueil et de traitement du signal
Les protocoles
Tester la réponse du système photopique
Tester la réponse du système scotopique
Tester la réponse des deux systèmes
Tester selon les espèces
Les facteurs affectant l’ERG flash
Les composantes de l’ERG et leurs origines
Indications de l’ERG flash chez l’animal
Exemples de résultats normaux
Situations cliniques
Le fond d’œil n’est pas visible
Les affections rétiniennes hérédodégénératives
Les anomalies génétiques des affections rétiniennes
Les troubles du comportement
Autres tests électrophysiologiques
Les autres ERGs
Les PEV
Contraintes anatomiques
1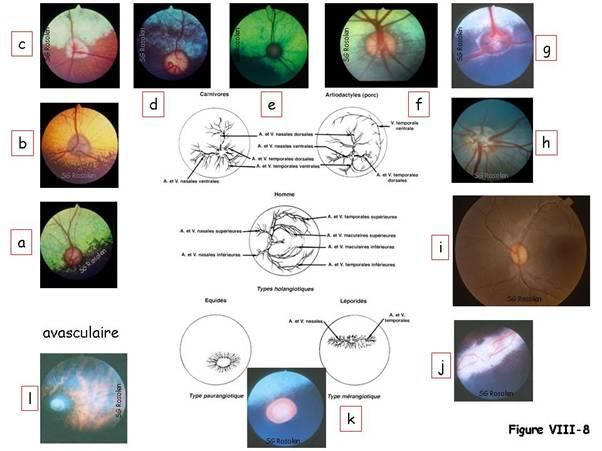
Œil et Physiologie de la Vision - VIII-1
Contraintes techniques
Conclusion
Texte intégral
Remerciements au Professeur Jean Sautet pour sa contribution aux illustrations d'anatomie
Introduction
L’appréciation de la fonction visuelle chez l’animal se fait sur des critères subjectifs
complétés par des tests subjectifs et objectifs.
Les critères subjectifs sont variés. On peut observer par exemple l’isolement d’un
individu au sein d’un troupeau de bovins ou d’ovins, l’apparition soudaine d’une
agressivité à l’encontre d’autres individus, une diminution notable de l’activité de jeu ou
de travail comme pour le chien de chasse ou encore une perte de poids chez un individu
au sein d’une colonie en élevage comme pour les rats, les souris ou les singes…
Ces signes déclenchent la mise en œuvre de tests subjectifs individuels d’évaluation de la
fonction visuelle comme la recherche du réflexe de clignement à la menace ou le test de
la boule de papier en vision binoculaire. Ces tests subjectifs individuels peuvent être
entrepris chez des espèces comme le chien ou le chat mais pas chez les espèces « mal
voyantes » comme les rongeurs (rat, souris, …). Dans le cas de ces espèces, importantes
pour la recherche biomédicale, il est nécessaire d’évaluer le fonctionnement de leur
système visuel par des tests objectifs électrophysiologiques.
Certains de ces tests électrophysiologiques sont utilisés en ophtalmologie vétérinaire,
c'est-à-dire pour les animaux de compagnie (chien, chat, cochons d’inde, lapins, etc.), de
rente (moutons, bovins, etc.) et chez certaines espèces sauvages maintenues en
captivité (oiseaux, primates, félins, etc.).
Ils permettent une évaluation objective essentiellement la fonction rétinienne de l’animal,
à l’aide de l’enregistrement de l’électrorétinogramme (ERG).
Intérêt de l’ERG en médecine vétérinaire
Les progrès de la médecine et de la chirurgie vétérinaires notamment en d’ophtalmologie
(chirurgie de la cataracte par incision étroite, mise en place d’implants intraoculaires,
etc) d’une part et la prise en compte, par la société, de l’animal en tant qu’être vivant
d’autre part, font que celui-ci est considéré comme un véritable patient, pouvant
bénéficier des mêmes approches diagnostiques et des mêmes stratégies thérapeutiques
que celles utilisées pour l’espèce humaine.
L’ERG est un examen complémentaire s’inscrivant dans cette démarche d’unicité de la
médecine [Zinsstag et al., 2005], [Cardiff et al., 2008] pour un bénéfice réciproque des
traitements de l’homme et de l’animal considéré comme un patient à part entière
[Enserink, 2007], [Rosolen SG, Picaud, 2010].
2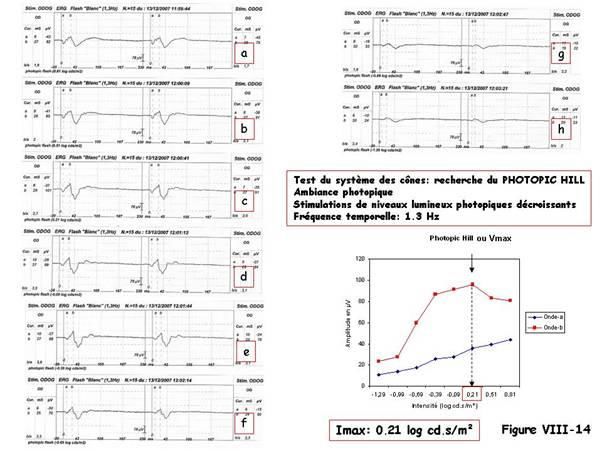
Œil et Physiologie de la Vision - VIII-1
But du chapitre
Ce chapitre a pour but de fournir les éléments essentiels à l’enregistrement et à
l’interprétation des examens électrorétinographiques effectués chez différentes espèces
d’animaux de compagnie atteints de maladies homologues ou non à des affections
rencontrées dans l’espèce humaine essentiellement des chiens et des chats.
L’enregistrement d’ERG effectués dans des conditions standards de stimulation, de
recueil et de traitement du signal est souvent indispensable pour le diagnostic et le
pronostic des affections concernées.
Au préalable, quelques caractéristiques anatomofonctionnelles visuelles animales sont
présentées, ainsi que les conditions spécifiques de mise en œuvre et de réalisation des
examens électrorétinographiques.
En fin de chapitre, quelques exemples de résultats sont commentés, enregistrés
essentiellement chez le chien.
Physiologie animale comparée
La rétine
Généralités
Bien que l’éventail des perceptions sensorielles des animaux soit très riche, c’est
néanmoins avec notre cerveau d’homme et non de poisson, d’oiseau, de chien ou de
cheval que nous analysons la façon dont les animaux se servent des informations reçues
et les interprètent.
Nous devons donc être très prudents dans l’interprétation des résultats des examens
pour l’évaluation de la fonction visuelle des espèces animales comme les vertébrés.
L’œil dans l’évolution
Au cours de l’évolution, l’œil et la fonction visuelle se sont avant tout adaptés au milieu
aquatique, terrestre ou aérien et au mode de vie nocturne ou diurne, de chaque espèce.
L’œil de type camérulaire
Chez tous les vertébrés, l’œil est de type camérulaire : c’est un objectif (le segment
antérieur avec ses lentilles et son diaphragme) et une chambre noire (le segment
postérieur) comportant un écran protecteur (absorbant l’excès de photons incidents) et
un capteur de photons (la rétine) qui transforme l’information lumineuse (énergie et
longueur d’onde) en un signal électrophysiologique (influx nerveux) interprétable par le
cerveau.
Les figures VIII-1, VIII-2, VIII-3 montrent les différents types d’yeux des vertébrés
aboutissants, chez les mammifères, au type camérulaire.
Figure VIII-1. L’œil de poisson (a) et (b) est gros et constitué d’une coque sclérale très
développée chez les espèces abyssales, permettant une résistance à la pression. Il
caractérisé par une cornée aplatie et un volumineux cristallin de forme sphérique non
déformable (a). Comme le cristallin n’est pas déformable, la mise au point
(accommodation) ne peut se faire que grâce à son déplacement d’avant en arrière à
l’aide d’un muscle (b).
3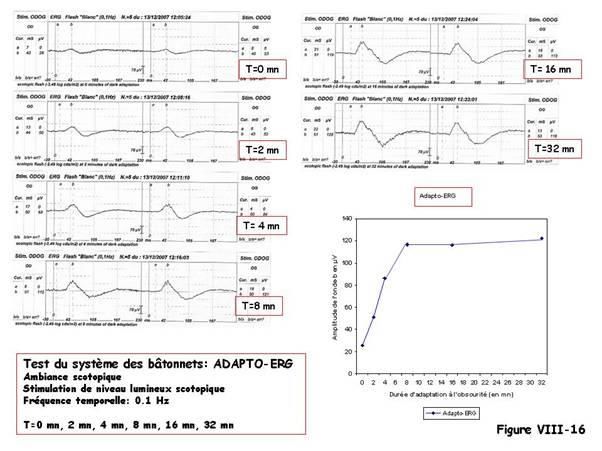
Œil et Physiologie de la Vision - VIII-1
Lors de la conquête du milieu aérien par les amphibiens (c) et les reptiles (d), les glandes
lacrymales et les paupières apparaissent. Elles permettent une hydratation de la cornée.
La cornée se bombe et cette nouvelle conformation (modification du rayon de courbure)
permet de s’adapter au changement d’indice de réfraction entre l’extérieur (air) et
l’intérieur (aqueux)… L’accommodation se fait par déplacement du cristallin, comme pour
les poissons.
Figure VIII-2. Chez les mammifères (e), le cristallin peut se déformer grâce à un système
complexe de muscles. Le mode de vie conditionne la forme et le volume du cristallin.
Chez les mammifères diurnes (f), le cristallin est lenticulaire alors que chez les espèces
nocturnes (g) (lémurien, rongeurs), le cristallin est sphérique et très volumineux (g) avec
une pupille en mydriase permanente qui laisse entrer une quantité maximale de lumière.
Figure VIII-3. Le système le plus élaboré se rencontre chez les oiseaux plongeur/pêcheur
comme le cormoran (h). L’œil est très volumineux, il occupe tout l’orbite. Lorsque
l’oiseau est en plongée, le muscle de Crampton permet l’aplatissement de la cornée
comme pour les poissons. Sous l’action du muscle de Brüch, le cristallin peut se déformer
jusqu’à former un lenticône. Le cormoran peut ainsi faire varier la puissance optique de
ses yeux d’environ 50 dioptries.
Le tableau VIII-1 résume les différents éléments anatomiques de l’œil des différents
vertébrés expliquant leur adaptation au milieu et leur mode de vie.
L’œil mobile
La vision n’est efficace que s’il y a mouvement car les capteurs visuels ne réagissent
qu’aux changements de luminosité. Ce mouvement peut être celui de l’objet observé ou
celui de l’œil lorsque l’objet est immobile. Dans ce cas, il y a déplacement de l’image sur
la rétine par des saccades oculaires.
Quelques particularités des rétines animales
La rétine inversée des vertébrés
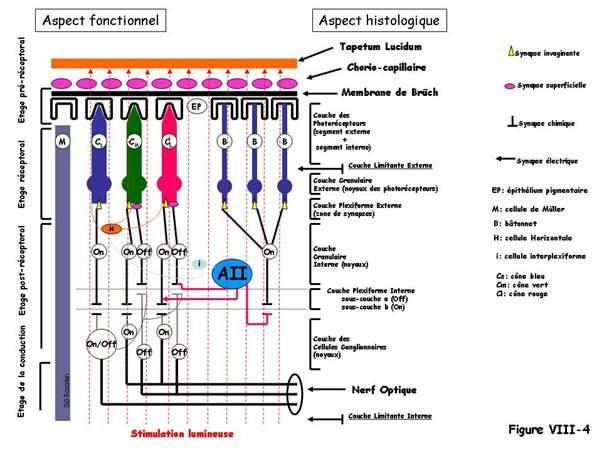
Chez tous les vertébrés, la rétine, présente à quelques variations près, le même schéma
fonctionnel (figure VIII-4) : a) un étage préréceptoral, b) un étage réceptoral composé
d’une très fine mosaïque de deux types de photorécepteurs : cônes et bâtonnets, c) un
étage postréceptoral avec cellules horizontales, bipolaires et amacrines et d) un étage de
conduction du message à partir des cellules ganglionnaires jusqu’au cortex visuel
primaire puis secondaire, très développé chez les mammifères.
La densité des deux types de photorécepteurs varie en fonction des espèces. Le tableau
VIII-2 présente ces différentes densités selon les différentes espèces et les auteurs
[Steinberg et al., 1973], [Wikler et al., 1990], [Zhang, Straznicky, 1991], [Jonas et al.,
1992], [Famiglietti, Sharpe, 1995], [Chandler et al., 1999], [Mowat et al., 2008].
Fovéa et area centralis
Animaux diurnes
La rétine des animaux diurnes (poissons, reptiles, oiseaux, carnivores, herbivores,
primates) est équipée de cônes nombreux, denses et diversifiés leur conférant la
perception des couleurs et une bonne acuité visuelle [Ebrey, Koutalos, 2001], [Wright,
Bowmaker, 2001] : poissons et oiseaux sont trichromates et la plupart des mammifères
sont dichromates (sauf l’homme et certains primates) [Packer et al., 1989], [Curcio et
al., 1990], [Jacobs, 1993], [Szel et al., 1996], [Ahnelt, Kolb, 2000].
4
Œil et Physiologie de la Vision - VIII-1
Une voire deux fovéas
Dans le monde diurne, le système rétinien le plus évolué pour discriminer les détails fins
d’un objet est constitué par une fovéa ; elle est présente chez les lézards, les primates.
On en trouve parfois deux chez certains oiseaux. L’aigle dispose d’un système optique
d’agrandissement et de deux fovéas : une fovéa monoculaire, centrale, qui observe et
une seconde fovéa binoculaire, temporale, qui permet un guidage très précis de
l’approche et de la capture de la proie entre les pattes.
Area centralis
Une autre particularité de la rétine des vertébrés est l’absence de macula, à l’exception
des primates non humains. Il existe cependant une zone appelée area centralis [Peichl,
1992] où la proportion de cônes est plus importante qu’ailleurs mais l’area centralis ne
présente pas de repères topographiques permettant de l’identifier par un examen
ophtalmoscopique.
La figure VIII-5 montre la régionalisation fonctionnelle de la neurorétine chez le chien.
Dans la partie ventrale, l’épithélium pigmentaire est pigmenté alors que dans la partie
dorsale, l’épithélium pigmentaire est dépigmenté, révélant la structure choroïdienne
appelé le Tapetum lucidum.
L’area centralis se situe dans la partie dorsolatérale à quelques millimètres au dessus de
la papille. Son diamètre est d’environ 0.2 mm chez le chien. Il n’y a pas de fovéa à
proprement parler car des bâtonnets y sont toujours présents. En cette zone centrale, les
cônes sont plus petits et plus minces qu’en rétine périphérique.
La densité des cônes y est d’environ 27 000/mm² ; elle décroît de façon importante en
périphérie pour atteindre une valeur d’environ 4 000/mm².
La densité des bâtonnets est sensiblement constante de l’ordre de 250 000/mm² sauf
autour de l’area centralis, où il existe une zone de densité maximale des bâtonnets au
environ de 450 000/mm².
Animaux nocturnes
Au contraire, la rétine des animaux nocturnes (poissons abyssaux, oiseaux, la plupart
des rongeurs de laboratoire) comporte essentiellement des bâtonnets qui favorisent leur
sensibilité à la lumière et aux mouvements, en ambiance scotopique [Szel, Rohlich,
1992], [Peichl, 2005], [Mustafi et al., 2009].
Une structure choroïdienne particulière : le Tapetum
Lucidum
Le fonctionnement de la rétine de tous les vertébrés est à peu près identique mais, d’un
point de vue ophtalmoscopique, les aspects des fonds d’yeux et de leur vascularisation
présentent de grandes variations. Elles sont en particulier liées à la présence d’une
structure choroïdienne spécifique : le Tapetum lucidum.
Structure choroïdienne, le Tapetum Lucidum est absent chez les rongeurs, le porc et les
primates et revêt de nombreuses variations chez les autres espèces.
Différents aspects du Tapetum Lucidum
Il représente plus des deux-tiers de la surface rétinienne chez le chat. Chez les
carnivores, il est de type cellulaire et sa composition est différente selon l’espèce. Chez le
chien, il est très riche en zinc-cystéïne [Lesiuk, Braekevelt, 1983], [Chijiiwa et al., 1990]
alors que chez le chat, il est essentiellement composé de riboflavines [Braekevelt, 1990].
Chez les ongulés, il est de type fibreux [Braekevelt, 1986].
5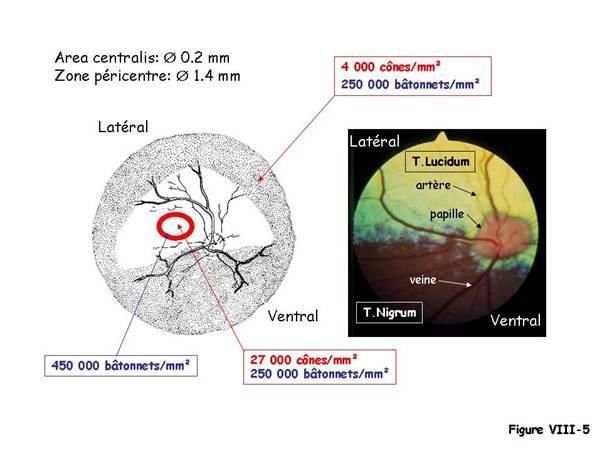
Œil et Physiologie de la Vision - VIII-1
La figure VIII-6 présente le fond d’œil de différentes espèces animales et ses variations
interspécifiques : un fond d’œil de cheval (a), de lapin (b), de mouton (c), de chat (d), de
porc (e), d’oiseau (f) et de primate (g), selon l’existence et la composition du Tapetum
lucidum.
Chez l’oiseau (f), il existe une structure choroïdienne très vascularisée : le pecten. Chez
les primates (g), on note aussi la présence d’une macula composée de la fovéa et la
fovéola.
Il existe des variations intraspécifiques comme chez le chien où les variations de couleur
du Tapetum lucidum dépendent de la couleur de la robe ; il est bleu-vert quand le poil
est foncé (h) et orangé quand la couleur du poil est clair (i).
Dans certains cas le Tapetum lucidum peut être absent ; l’épithélium pigmentaire est
pigmenté sur toute sa surface et on parle d’aspect tigroïde (j). Dans d’autres cas, il
existe à la fois une absence de Tapetum lucidum et une absence de pigmentation de
l’épithélium pigmentaire (k), c’est le cas de certains chiens qui présentent un phénotype
sub-albinos comme les chiens de race Sibérien Husky (k).
La figure VIII-7 présente des coupes de rétine réalisées dans une zone tapétale et dans
une zone non tapétale, sur la même rétine. En regard de la zone avec Tapetum lucidum
(TL), l’épithélium pigmentaire rétinien (EPR) est dépigmenté alors qu’en regard de la
zone dépourvue de Tapetum lucidum aussi appelée Tapetum nigrum (TN), l’EPR est
pigmenté. Le Tapetum lucidum aurait pour rôle d’augmenter le rendement des photons
incidents.
Différents types de vascularisation
Il est intéressant de noter que les variations de la vascularisation du fond d’œil n’ont
aucune relation avec le mode de vie de l’animal. La rétine est avasculaire chez le cobaye
(animal diurne avec plus de 15% de cônes), alors qu’elle est très vascularisée chez le rat
(animal nocturne avec moins de 1% de cônes).
Chez les mammifères, à l’exception des primates, il n’y a pas d’artère centrale de la
rétine ; la vascularisation rétinienne a pour origine l’artère carotide externe, sauf chez les
primates où elle provient de l’artère carotide interne.
On peut observer plusieurs types de vascularisation rétinienne chez les mammifères par
exemple de type holangiotique chez les carnivores.
Rappel. La vascularisation de type holangiotique correspond à un apport sanguin
direct de l’ensemble de la rétine. Dans le type mérangiotique, les vaisseaux se
localisent à une partie seulement de la rétine et dans le type paurangiotique, la
vascularisation rétinienne se localise à la région péripapillaire.
La figure VIII-8 montre différents types de vascularisation holangiotique du fond d’œil
gauche chez des mammifères : fonds d’yeux °de chiens de race Cocker (a), Boxer (b),
Akita Inu (c) et berger allemand (d), °de chats européens (e). On rencontre également
ce type de vascularisation chez les artiodactyles comme la chèvre (f), les bovins (g), les
porcins (h) et les primates (i). Le type de vascularisation mérangiotique se voit chez les
lagomorphes (lapin) (j) et le type paurangiotique chez le cheval (ongulé) (k). Il existe
également un fond d’oeil avasculaire chez le cobaye (l).
Des espèces pigmentées et des espèces albinos
Il existe de nombreuses espèces animales de phénotype albinos et dont l’épithelium
pigmentaire est dépigmenté.
6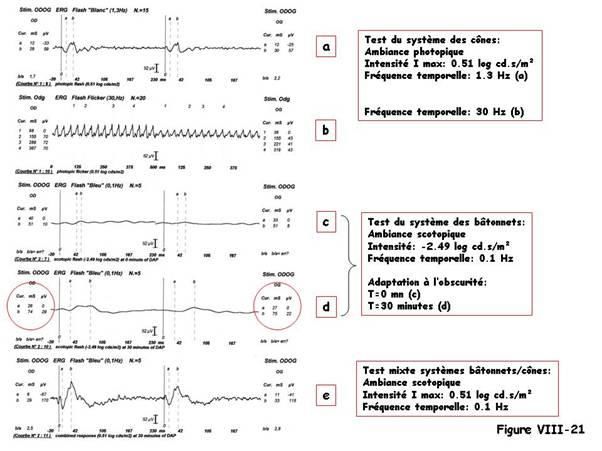
Œil et Physiologie de la Vision - VIII-1
La figure VIII-9 présente des exemples de fonds d’yeux d’animaux albinos chez des
espèces nocturnes comme le rat (a), la souris (b) et des espèces vivant en milieu
mésopique comme le cobaye (c) ou le lapin (d et e). Cependant on peut aussi rencontrer
des fonds d’yeux albinos chez certaines espèces diurnes comme le chat siamois (f et g)
ou le chien (h et i).
Cette absence de pigment doit être prise en considération lors de l’interprétation des
examens électrorétinographiques lors de la comparaison entre des espèces pigmentées
et des espèces ou des individus albinos [Wali, Leguire, 1992a], [Behn et al., 2003],
[Rosolen et al., 2005b] car les rétines des espèces pigmentées s’adaptent plus vite à
l’obscurité que celles des espèces albinos.
Les voies visuelles
Description anatomofonctionnelle
Chez les mammifères domestiques, la conformation anatomique des voies visuelles est
celle des primates ; seul le pourcentage de fibres décussant varie en fonction du degré
de latéralité des yeux.
Nerfs optique
L’ensemble des axones des cellules ganglionnaires se regroupe pour constituer le nerf
optique. Ces fibres optiques croisent partiellement au niveau du chiasma optique.
Fibres directes
Les fibres directes sont les fibres temporales (couvrant la partie nasale du champ visuel
binoculaire) ; elles sont relativement plus nombreuses dans les espèces où les yeux sont
frontaux et le champ binoculaire étendu (50% chez les primates) que dans les espèces
dont les champs monoculaires sont très étendus et les yeux latéraux (30-35% chez les
carnivores ; 16% chez le cheval et 10% chez le lapin).
Fibres croisées
Quelques fibres croisées mettent en relation la rétine avec l’hypothalamus et le
mésencéphale et forment le faisceau para-optique ou système optique accessoire.
Tractus optique et voies rétinotectales
Le tractus optique rassemble les fibres des deux hémirétines homolatérales et se divise
en deux racines, l’une médiale et l’autre latérale. La racine médiale entièrement croisée
est empruntés par les voies optiques rétinotectales à conduction lente et la racine
latérale, partiellement croisée est suivie par les voies optiques rétinotectales à
conduction plus rapide.
Le système optique rétinotectal est le seul système existant chez les vertébrés sans
néocortex (poissons, amphibiens, reptiles et oiseaux). Il ne permet à ces espèces que de
détecter les mouvements.
Chez les mammifères, la racine médiale du tractus optique aboutit aux collicules rostraux
(voie rétinotectale) tandis que la racine latérale qui se poursuit vers les corps géniculés
latéraux, forme la voie rétinocorticale dont le fonctionnement est similaire chez tous les
mammifères (y compris chez l’homme).
7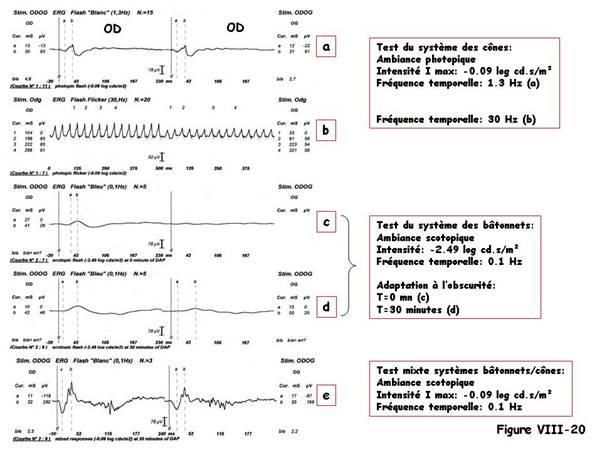
Œil et Physiologie de la Vision - VIII-1
Cortex visuel
Cependant la topographie du cortex visuel primaire peut varier en fonction de l’espèce et
même au sein d’une espèce donnée. C’est le cas de l’espèce canine qui, pour un même
génotype, présente plus de 350 phénotypes différents dont le poids varient de quelques
centaines de grammes (Chihuahua) à plusieurs dizaines de kilogrammes (Matin de
Naples) et dont la morphologie de la boîte crânienne varie du type longiligne (Lévriers,
Colleys) au type bréviligne (Boxer, Bulldog)…
Exploration rétinienne par ERG
L’ERG est l’examen fonctionnel de choix utilisé en médecine vétérinaire. Il est réalisable
dans des conditions cliniques. Il fournit des données quantifiables et comparables au
cours du temps.
De plus, les appareils disponibles sur le marché vétérinaire permettent d’effectuer des
enregistrements électrorétinographiques et un traitement du signal dans des conditions
de reproductibilité acceptables.
Cependant, cet examen de la fonction rétinienne reste un examen complémentaire qui
doit être replacé dans le contexte clinique pour lequel il a été demandé.
Les conditions de genèse de l’ERG flash
L’exploration fonctionnelle par ERG chez les animaux, sujets non verbal et non
coopératifs, doit se faire sous anesthésie générale.
Les conditions de stimulation et de recueil du signal doivent être effectuées dans des
conditions de reproductibilité afin de limiter les facteurs de variabilité et d’obtenir des
réponses interprétables.
L’éclairement rétinien doit être constant et uniforme pendant toute la durée des
différentes séquences de la procédure.
La gestion de ces contraintes techniques, la connaissance anatomophysiologique de
l’espèce examinée et l’application de protocoles raisonnés sont alors des éléments
essentiels pour l’interprétation correcte de l’ERG chez l’animal.
Le choix du stimulateur et de la stimulation
Le stimulateur
Pour des raisons de conformation anatomique de la tête (présence d’un museau allongé)
et de contrôle de la ventilation des animaux anesthésiés, par sonde endotrachéale,
l’utilisation d’une coupole unique ou « Ganzfeld » est difficile sauf pour des espèces de
petite taille.
La plupart des systèmes proposés chez l’animal pour réaliser des ERG sont des
photostimulateurs doubles équipés de lampes xénon ou de LED achromatiques. Ils sont
orientables pour que leur plan de stimulation soit parallèle au plan pupillaire (figure VIII-
10).
8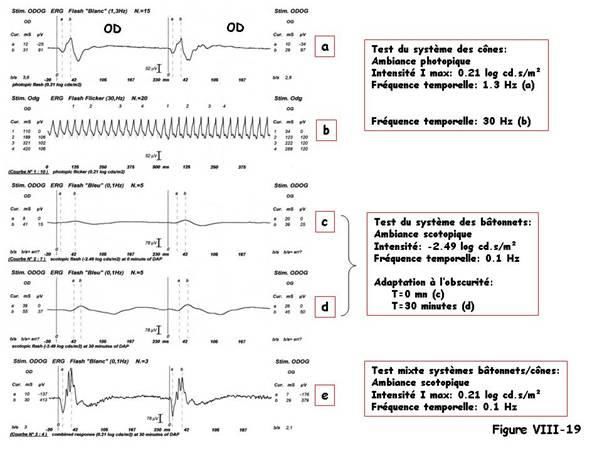
Œil et Physiologie de la Vision - VIII-1
Niveau lumineux de la stimulation
Les stimulations utilisées sont achromatiques, d’intensités modulables, croissantes à
partir d’une valeur minimale de 0,05 cd.s/m² (ou cd.m-².s) (stimulation de niveau
lumineux scotopique) jusqu’à une valeur maximale de 6,4 cd.s/m² (stimulation de niveau
lumineux photopique). Il est également possible de délivrer d’abord la stimulation la plus
intense puis les suivantes par ordre décroissant. Le pas entre chaque stimulation
successive est de 0,3 unité logarithmique.
Cela revient à dire, par exemple pour les stimulations croissantes, qu’on double la
suivante par rapport à la précédente. En effet si L2 = 2L1 : L2/L1 = 2 et log10 2 =
0,3. On peut exprimer les luminances des stimulations directement en log10 et
passer de la précédente à la suivante en ajoutant 0,3 unité log (log102). Voir
tableau VIII-3 pour les conversions.
Les conditions de stimulation
Adaptation rétinienne
Selon que l’examen s’applique à des espèces diurnes ou nocturnes, l’adaptation de la
rétine à une ambiance stable n’est pas la même.
Deux paramètres sont à considérer, d’une part, l’état d’adaptation de la rétine lié aux
cycles circadiens et, d’autre part, le fond adaptant de niveau lumineux constant et
uniforme qui permet d’obtenir un état d’adaptation rétinien stable sur lequel une
stimulation peut être délivrée.
Espèces diurnes
Pour les espèces diurnes, les sujets sont placés dans une pièce où règne une ambiance
de niveau lumineux photopique modéré stable. Ils y resteront au moins deux heures
[Marmor MF, Lurie M, 1979], [Marmor, 1991]. Cette ambiance photopique stable est
mise à profit pour réaliser l’anesthésie et la préparation du sujet. Ensuite les sujets sont
placés devant un fond adaptant de niveau lumineux constant et uniforme pendant au
moins 10 minutes [Marmor et al., 2009].
Espèces nocturnes
Pour les espèces nocturnes, les sujets sont mis dans une pièce obscure pendant plusieurs
heures. L’anesthésie et la préparation du sujet se font à l’obscurité.
Stimulations « plein champ »
La plupart des appareils qui permettent d’enregistrer des ERG chez l’animal de taille
moyenne ou de grande taille, sont constitués de deux photostimulateurs équipés de
lampes xénon ou de LED (figure VIII-10).
La surface stimulante de ces photostimulateurs doit être suffisamment large pour obtenir
des stimulations de type « plein champ ». Pour cela, le photostimulateur doit être placé
de telle sorte que l’angle sous lequel la surface stimulante est vue, soit le plus large
possible.
De plus, l’éclairement rétinien dépendant du carré de la distance qui sépare la rétine
stimulée de la surface stimulante, il faut toujours placer le stimulateur à la même
distance de la cornée afin de se placer dans des conditions reproductibles de stimulation.
La figure VIII-11 montre la taille de la surface stimulante (a), le positionnement de l’œil
et celui du photostimulateur (d) afin de réaliser des stimulations dans des conditions
« plein champ ».
9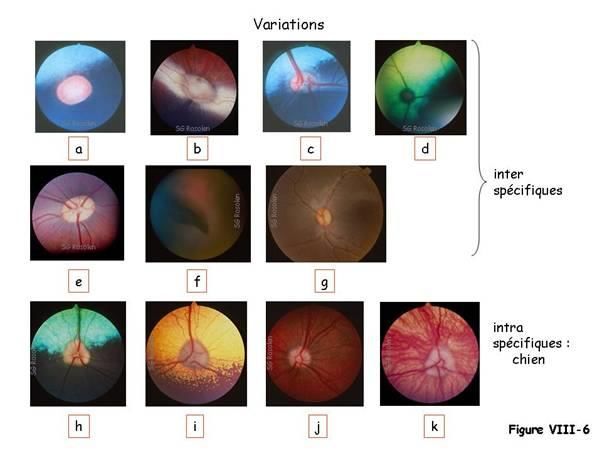
Œil et Physiologie de la Vision - VIII-1
Rappel. Avec le stimulateur utilisé, la surface stimulante (a) (source LED +
diffuseur) a un diamètre de 5,5 cm. Elle doit être placée à une distance inférieure à
1,5 cm afin que l’angle sous lequel est vue la stimulation soit supérieur à 130° :
c’est une stimulation « plein champ » (figure III-12).
Les conditions de recueil et de traitement du signal
La préparation du sujet
L’anesthésie
Une simple sédation ne suffit pas pour réaliser des ERG chez l’animal car l’utilisation de
flashs peut provoquer des mouvements de recul et des clignements de paupières.
Quelle que soit l’espèce, les examens se font tous sous anesthésie générale. Une voie
d’abord veineuse et une sonde endotrachéale sont utilisées afin d’assurer une anesthésie
dans des conditions de sécurité optimale.
Plusieurs types d’anesthésie peuvent être utilisés. Il convient donc de connaître
précisément les modifications induites par les différentes substances utilisées sur les
paramètres de l’ERG (diminution/augmentation des amplitudes, des temps de
culmination, disparition de certaines ondes) et d’en tenir compte lors de l’interprétation
des ERG [Morgan, Ward, 1970], [van Norren, Padmos, 1975], [van Norren, Padmos,
1977], [Stute A et al., 1978], [Raitta et al., 1982], [Murray, Borda, 1984], [Jarkman et
al., 1985], [Wasserschaff, Schmidt, 1986], [Malecki H et al., 1996], [Yanase, Ogawa,
1997], [Sloan, 1998], [Clarke, 1999], [Chaudhary et al., 2003], [Kommonen et al.,
2007], [Ropstad et al., 2007], [Norman et al., 2008], [Varela Lopez et al., 2010] (voir ci-
dessous).
Température corporelle
Pendant toute la durée de l’anesthésie, il convient également de maintenir la
température corporelle constante par des systèmes chauffants. Une baisse de
température de quelques degrés peut entraîner, par exemple, des diminutions
d’amplitudes des ondes-a et b de l’ERG [Mizota, Adachi-Usami, 2002], [Kong, Gouras,
2003].
La fixation du globe
Pendant l’anesthésie, le globe oculaire peut effectuer des mouvements de rotation
entraînant une modification de l’axe optique. Les conditions de stimulation et
d’enregistrement risquent ainsi d’être modifiées en cours d’enregistrement.
Il convient donc d’utiliser un système qui permet de fixer le globe pendant toute la durée
de l’examen. Un simple blépharostat permet de maintenir les paupières ouvertes mais
non pas d’assurer la fixité du globe.
La fixation peut être faite au moyen de fils de traction ou bien en utilisant les clips
servant à la fois de fixateur et d’électrode active (électrode clip, Siem-Biomédicale,
Nîmes, France)
Dilatation pupillaire
Une dilatation pupillaire pharmacologique (Tropicamide®) permet d’obtenir un diamètre
pupillaire constant pendant toute la durée de l’examen. Il est mesuré au début (figure
VIII-11-b et c) et à la fin des enregistrements grâce à un compas à strabisme pour
s’assurer que le diamètre pupillaire reste de taille constante durant toute la procédure.
Cette dilatation pupillaire est gage d’enregistrement des « meilleures » réponses c'est-à-
dire des réponses les plus amples.
10Œil et Physiologie de la Vision - VIII-1
Figure VIII-11 : la taille pupillaire mesurée chez ce chien atteint de cataracte après
dilatation est de 11 mm (c). Une fois ce diamètre pupillaire mesuré, le photostimulateur
(a) est rapproché (b) à une distance inférieure à 2,25 cm (flèche rouge) (d) puis toujours
replacé à la même distance lors d’examens ultérieurs.
Les conditions de stimulation plein champ, dilatation pupillaire maximale, constance de la
distance œil-surface stimulante sont des gages de reproductibilité des réponses pour tous
les examens ultérieurs.
Les différents types d’électrodes de recueil du signal
Electrodes actives
Plusieurs types d’électrodes actives peuvent être utilisés chez l’animal pour le recueil de
l’ERG [Steiss JE et al., 1992], [Esakowitz et al., 1993], [Hennessy, Vaegan, 1995],
[Bayer et al., 1999], [Rosolen et al., 2002], [Yin, Pardue, 2004], [Mentzer et al., 2005].
Les électrodes cornéennes permettent d’avoir un bon contact mais l’angle de courbure
cornéen varie en fonction de l’espèce animale et il est difficile d’utiliser les mêmes
électrodes que celles commercialisées pour usage humain.
La figure VIII-13 montre des réponses ERG obtenues chez le même animal dans les
mêmes conditions de stimulation « plein champ » œil par œil, en utilisant différents types
d’électrodes de recueil.
Figure VIII-13-a. L’électrode aiguille active est piquée en partie dans la paupière
inférieure en voie sous-cutanée. La réponse de l’œil gauche est contaminée par de
l’électromyogramme. Bien que l’œil de l’animal soit normal, les réponses enregistrées
pour chacun des deux yeux ne sont pas superposables.
Figure VIII-13-b. L’électrode aiguille active est placée dans le canthus externe, près du
pôle postérieur. Il n’est pas possible d’en contrôler la direction qui diffère nécessairement
entre l’œil gauche et l’œil droit. Cette variation de direction de recueil se traduit par des
réponses différentes pour chaque œil. Dans ce cas également, les réponses ne sont pas
superposables.
Figure VIII-13-c. L’électrode est de type cornéenne : c’est une électrode JET, constituée
d’un anneau d’or. L’absence de fixation du globe se traduit par une dissymétrie entre les
réponses recueillies sur l’œil droit et sur l’œil gauche.
Figure VIII-13-d. L’électrode clip est placée en région sous-conjonctivale, à 12h et à
moins d’un millimètre du limbe. Le clip assure à la fois le recueil du signal, la fixation du
globe et une légère traction vers l’avant qui évite tout prolapsus de la membrane
nictitante, très développée chez les animaux. Les réponses obtenues pour chaque œil
sont superposables. L’usage de cette électrode est atraumatique ; elle permet de
renouveler des enregistrements chez le même animal au cours de sessions différentes et
de recueillir des réponses reproductibles.
Remarque importante. Pour chaque enregistrement, il est indispensable de préciser
la méthode utilisée ainsi que le coefficient de variation des paramètres de l’ERG en
fonction de la technique utilisée, pour permettre la comparaison des résultats.
Le tableau VIII-4 montre les coefficients de variations des paramètres de l’ERG
enregistrés avec différents types d’électrodes, selon différents auteurs. Il apparaît que
quels que soient les auteurs et le type d’électrodes, le coefficient de variation des temps
de culmination est nettement inférieur à celui des amplitudes, quelle que soit l’onde
mesurée. Cependant lorsque le globe est fixé (clip), le coefficient de variation des
amplitudes mesurées est inférieure à celui obtenu avec une électrode par simple contact.
11Œil et Physiologie de la Vision - VIII-1
Il existe également des électrodes actives constituées d’un stimulateur LED intégré
[Kooijman, Damhof, 1980], [Kooijman, Damhof, 1981]. Les résultats présentés dans le
tableau VIII-4 par Maehara ont été obtenus avec ce type d’électrode de contact
contenant un stimulateur intégré.
Electrodes de références
Les électrodes de référence doivent être placées en sous-cutanée stricte afin d’éviter
l’enregistrement de l’électromyogramme. Ce sont, de préférence, des aiguilles à
acupuncture qui évitent tous risques d’hémorragies sous-cutanées.
Le traitement du signal
Il est identique à celui pratiqué chez l’homme décrit au chapitre V-1.
Les protocoles
Tester la réponse du système photopique
L’étude du système photopique se fait en ambiance photopique avec des stimulations de
niveaux lumineux photopiques. On utilise d’abord une stimulation flash de basse
fréquence temporelle (1 à 2 Hz) dite « stimulation flash » d’intensité croissante jusqu’à
obtenir la valeur maximale de l’amplitude de l’onde-b de la cone-response dite « Photopic
Hill ». Cette intensité maximale est ensuite utilisée comme stimulation à la fréquence
temporelle de 30 Hz, dite « stimulation flicker » pour obtenir la flicker-response.
Ces deux modes de stimulation permettent de recueillir les réponses du système
photopique provenant de structures d’origines prépondérantes différentes.
Cone-response d’amplitude maximale ou Photopic Hill
Dans une ambiance de niveau lumineux photopique suffisante pour que les bâtonnets
fonctionnent en mode saturé, l’augmentation (ou la diminution à partir d’une valeur
maximale) de l’intensité de la stimulation (par pas de 0,3 unité logarithmique) entraîne
une variation conjointe de l’amplitude de l’onde-a et de l’onde-b.
Augmentation linéaire de l’onde-a
L’amplitude de l’onde-a croît de façon linéaire en fonction du logarithme de l’intensité de
la stimulation (figure VIII-14).
Augmentation non-linéaire de l’onde-b
Par contre, l’amplitude de l’onde-b commence par croître, passe par un maximum puis,
alors que l’intensité de la stimulation continue à croître, diminue jusqu’à un plateau
(figure VIII-14. Cette amplitude maximale de l’onde-b est dite « Photopic Hill » ou Vmax
[Peachey et al., 1992], [Wali, Leguire, 1992b], [Lachapelle et al., 2001], [Rufiange et al.,
2003].
° Origine physiologique probable. Cette variation non linéaire de l’amplitude de l’onde-b
de la cone-response de l’ERG flash est probablement le résultat de l’inégale contribution
à sa constitution des réponses des cellules bipolaires de cônes ON et OFF. Jusqu’à
l’intensité Imax, l’amplitude de l’onde-b résulterait d’une participation des réponses des
bipolaires de cônes ON, prépondérante sur celle des bipolaires de cônes OFF (donc
davantage de dépolarisation) puis, l’intensité continuant à croître, l’amplitude de l’onde-b
résulterait d’une participation plus importante des réponses des bipolaires de cônes OFF
(donc davantage d’hyperpolarisation) ayant pour conséquence une décroissance de
l’amplitude de l’onde-b [Lachapelle, et al., 2001], [Rufiange, et al., 2003].
12Œil et Physiologie de la Vision - VIII-1
Courbe « Intensité-réponse »
Il est possible d’établir une courbe « intensité-réponse » pour chaque espèce testée et
chaque sujet testé. Pour se faire, on recueille en ambiance photopique (luminance
comprise entre 20 cd/m² et 30 cd/m²), les ERG flash évoqués par 10 à 15 stimulations
d’intensités successivement croissantes par pas de 0.3 unité logarithmique entre deux
intensités successives et de basse fréquence temporelle.
L’intensité peut avoir des valeurs décroissantes à partir d’une valeur maximale
comme sur la figure VIII-14 : réponses de (a) à (h), les réponses étant
rigoureusement superposables [Wali, Leguire, 1992b].
On porte en abscisse, le logarithme de l’intensité de la stimulation et, en ordonnée,
l’amplitude de l’onde-b de la cone-response, puis on détermine l’intensité dite Imax qui
correspond au Photopic Hill (ou Vmax). Dans l’exemple donné Imax = 0,21 log cd.s/m².
La figure VIII-14 montre comment rechercher le Photopic Hill chez un chien en utilisant
une fréquence temporelle de 1.3 Hz.
L’intensité la plus forte (a) correspond à 0.81 log cd.s/m², la plus faible (h) est de -
1.29 log cd .s/m². Chaque intensité intermédiaire (b-g) correspond à un pas de 0.3
log cd.s/m². Dans ce cas l’intensité pour laquelle on mesure une amplitude de
l’onde-b maximale est de 0.21 log cd.s/m². On notera une légère dissymétrie entre
œil droit (101 µV) et œil gauche (91 µV), de l’ordre de 10%. C’est une différence
interoculaire qui reste dans les limites de la normale chez le chien.
° Intérêt de déterminer et d’utiliser Imax. L’amplitude maximale est une caractéristique
du système photopique indépendante de l’intensité du fond adaptant [Lachapelle, et al.,
2001]. Figure VIII-15-a montre la cone-response d’un sujet évoquée avec Imax
précédemment déterminée (voir figure VIII-14-c).
Utiliser l’intensité Imax de chaque individu pour évoquer la réponse du système
photopique permet de se situer dans des conditions de stimulations optimales et
reproductibles. Cette intensité peut être différente selon les individus testés, mais sa
mise en œuvre pour l’enregistrement des réponses du système photopique de chacun,
limite significativement la variabilité interindividuelle des amplitudes des ondes-b.
Flicker-response avec des stimulations flash de fréquence
temporelle 30 Hz
La stimulation flash de niveau lumineux photopique et d’intensité correspondant à Imax,
est délivrée à la fréquence temporelle de 30 Hz pendant au moins 15 secondes sur le
même fond adaptant photopique que celui utilisé précédemment. Elle est dite
« stimulation flicker ». On recueille la flicker-response ou réponse flicker qui est le reflet
des structures photopiques essentiellement postréceptorales (figure VIII-15-b).
Tester la réponse du système scotopique
La « stimulation flash »
Le test du fonctionnement du système des bâtonnets s’effectue en ambiance scotopique
(obscurité). La stimulation est achromatique, de niveau lumineux scotopique de 2,5 à 3
unités logarithmiques inférieures à celui utilisé pour tester le système photopique.
Elle est délivrée à une fréquence temporelle de 0,1 Hz, répétée une, trois ou cinq fois à
différents temps d’adaptation à l’obscurité : juste à la mise à l’obscurité après une
adaptation à un fond adaptant de niveau lumineux photopique, au temps T=0 alors qu’il
n’y a pas encore eu le temps nécessaire pour que la rétine puisse s’adapter à l’obscurité,
puis au moins après 30 minutes d’adaptation à l’obscurité, lorsque la rétine est
totalement adaptée à l’obscurité (figure VIII-16 et figure VIII-17 T=0 mn et T=32 mn).
13Œil et Physiologie de la Vision - VIII-1
L’adapto-ERG
[Alfiéri R, Solé P, 1966]. Entre le moment de la mise à l’obscurité (t 0=0) et durant les 30
minutes que dure l’adaptation de la rétine à l’obscurité, il est possible d’enregistrer des
ERG flash à intervalles réguliers du temps d’adaptation ; on a choisi t1=2mn, t2=4mn,
t3=8mn, t4=16mn, t5=32mn après la mise à l’obscurité.
Pour évoquer l’ERG à chaque temps ti dans l’ambiance scotopique, on utilise de 3 à 5
stimulations flash, toujours d’un même niveau lumineux scotopique, délivrées à la
fréquence temporelle de 0,1 Hz.
L’amplitude de l’onde-b des ERG est ensuite mesurée ; elle peut être portée en fonction
du temps d’adaptation : c’est l’« adapto-ERG » ; elle montre que l’amplitude croît de
façon régulière de plus de 30%, associée à une augmentation du temps de culmination
de l’onde-b de plus de 2 écarts-type.
La figure VIII-16 montre les réponses du système scotopique à une stimulation de
niveau lumineux scotopique toujours identique, à différents temps d’adaptation à
l’obscurité. L’amplitude de l’onde-b croît. L’ «adapto-ERG » correspond à la
représentation de l’amplitude de l’onde-b en fonction du temps d’adaptation à
l’obscurité.
Bases physiologiques
Le potentiel de l’épithélium pigmentaire est stable quand l'adaptation de la rétine à une
ambiance photopique est d’au moins deux heures [Marmor MF, Lurie M, 1979], ou celle à
une ambiance scotopique d’au moins 40 minutes [Giessmann, Lutze, 1971] [Le Grand Y,
1972].
La différence de potentiel entre la cornée et la rétine dépend de la valeur du potentiel de
l'épithélium pigmentaire qui est variable selon l’état d’adaptation de la rétine à la lumière
ou à l’obscurité. En effet, lors du passage d'un état d'adaptation à l'autre, le potentiel de
l'épithélium pigmentaire passe par une variation maximale de la polarisation de sa
membrane basale, qui survient entre la 5ième et la 10ième minute après le changement
d'état d'adaptation.
Chez l’homme, cette variation maximale de polarisation de l’épithélium pigmentaire peut
être appréciée par l’enregistrement de l’EOG. Elle correspond à la survenue du Light Peak
en ambiance photopique ou du Dark Trough en ambiance scotopique.
Lorsque l’ERG est enregistré à intervalles réguliers au cours de l’adaptation à l’obscurité à
l'aide d'une stimulation flash de niveau lumineux scotopique (donc n’évoquant que la
seule réponse du système scotopique) juste après que la rétine ait été préalablement
adaptée à la lumière, l’amplitude de l’onde-b dépend étroitement d’un ensemble de
mécanismes intra-épithéliaux à l’origine de l’évolution de son potentiel.
Intérêt de l’adapto-ERG
Suivre la variation de l’amplitude de l’onde-b de l’ERG du système des bâtonnets au
cours de l’adaptation à l’obscurité permet d’apprécier la cinétique de l’adaptation de la
rétine liée au fonctionnement conjoint du système des bâtonnets et de l’épithélium
pigmentaire, donc d’apprécier, de façon indirecte, le fonctionnement de l'épithélium
pigmentaire.
Dans l’impossibilité d’enregistrer un EOG chez l’animal, l’adapto-ERG permet d’apprécier
de façon indirecte certains aspects du fonctionnement de l’épithélium pigmentaire sous la
dépendance du système des bâtonnets. Il exclut cependant le reflet du fonctionnement
de la membrane basale [Rigaudière F. et al., 2005].
14Œil et Physiologie de la Vision - VIII-1
Tester la réponse des deux systèmes
En fin d’adaptation à l’obscurité, toujours en ambiance scotopique, on peut délivrer une
stimulation unique, de niveau lumineux photopique et d’intensité correspondant à la
valeur du Imax. Elle évoque la réponse conjointe des deux systèmes des cônes et des
bâtonnets : c’est la mixed-response (figure VIII-17).
Tester selon les espèces
Les espèces diurnes
Pour les espèces diurnes, on s’intéresse essentiellement au fonctionnement du système
des cônes dans une moindre mesure celui du système des bâtonnets.
Les espèces nocturnes
Pour les espèces nocturnes dont la rétine est pauvre en cônes, on étudie principalement
le fonctionnement du système des bâtonnets.
Les facteurs affectant l’ERG flash
Le type d’anesthésique
Différents types d’anesthésiques sont utilisés selon les espèces, la durée et la profondeur
de l’anesthésie obtenue. Les molécules circulantes peuvent interférer avec les différents
neurotransmetteurs rétiniens. Dans certains cas, les molécules d’anesthésique injectées
peuvent bloquer la transmission du signal intrarétinien. Lors de la stimulation lumineuse,
une forte diminution d’amplitude d’une ou de plusieurs réponses selon les séquences
mises en œuvre, voire une absence d’ERG, sont directement liées à ce blocage des
processus intrarétiniens. Il ne faut pas alors en conclure que la rétine présente une
altération intrinsèque puisque la réponse recueillie n’est liée qu’à la modification
fonctionnelle conjoncturelle.
Chez l’animal, les anesthésiques les plus fréquemment utilisés sont soit injectés par voie
veineuse ou intramusculaire, soit inhalés. Les effets observés dépendent de la dose
utilisée mais également de la sensibilité de l’espèce animale.
Parmi les substances utilisées, chez le rat, le telazol -un anesthésique dissociatif- induit
une augmentation d’amplitude surtout de l’onde-b et une augmentation des temps de
culmination des réponses par rapport à celles obtenues avec du nembutal [Chaudhary, et
al., 2003].
Chez le chien, l’association kétamine-acépromazine induit une augmentation de
l’amplitude de l’onde-b au cours de l’adaptation à l’obscurité et une réduction du temps
d’obtention de la réponse maximale par rapport à l’utilisation de la kétamine seule
[Malecki H, et al., 1996].
Autre substance fréquemment utilisé chez les carnivores (chien et chat), un
alpha2agonist (hydrochloride de médétomidine) induit une diminution des amplitudes et
une augmentation des temps de culmination des ondes [Norman, et al., 2008].
L’utilisation d’anesthésiques volatils (halothane et sévoflurane) peut induire une
diminution de l’amplitude de l’onde-b dans des conditions scotopiques [Yanase, Ogawa,
1997] mais dans ce cas, c’est surtout l’hypercapnie qui est à l’origine de cette diminution
d’amplitude [Varela Lopez, et al., 2010].
Il faut donc soigneusement choisir l’anesthésique qui doit être bien adapté à l’animal
testé et qui doit présenter le meilleur compromis entre les différentes contraintes
physiologiques et techniques.
15Vous pouvez aussi lire

















































