Caractérisation moléculaire par microsatellites de trois races chevaline en Tunisie
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
DIVERSITÉ GÉNÉTIQUES DES RACES CHEVALINES TUNISIENNE 103
Caractérisation moléculaire par microsatellites de
trois races chevaline en Tunisie
H. OULED AHMED1, M.FATNASSI2*, H. FERCHICHI2, R. BACCOUCHE4, F. LASFAR3, M. ZAOUIA3,
A BEN YOUNES3, M.HADDAD3, A. TRIMECHE2
1
Laboratoire d’Analyses Génétiques Animales, Institut de la Recherche Vétérinaire de Tunisie 1006, Tunisie
2
Gestion de la santé et de la qualité des productions animales, Service de zootechnie, Ecole de médecine vétérinaire, Sidi-Thabet 2020, Tunisie.
3
Fondation Nationale d’Amélioration de la Race Chevaline, Sidi-Thabet 2020, Tunisie
4
Institut National Agronomique de Tunisie, 43, Avenue Charles Nicolle 1082 -Tunis- Mahrajène Tunisie.
*Auteur chargé de la correspondance : fatnassi_meriem@yahoo.com
RÉSUMÉ ABSTRACT
L’objectif de cette étude est l’évaluation de la diversité génétique de 3 races Molecular characterization of three horse breeds in Tunisia using
chevalines en Tunisie. Dix-sept marqueurs microsatellites ont été utilisés microsatellites markers
pour le génotypage de 300 chevaux appartenant aux 3 races (100 Barbe,
100 Arabe Barbe et 100 Pur-Sang Arabe). Les 17 microsatellites utilisés This study aimed to evaluate the genetic diversity of 3 horse breeds in Tunisia.
étaient fortement polymorphes avec un nombre d’allèles par locus variant A total of 17 microsatellite markers was used for the genotyping of 300 horses
entre 8 et 25. Le nombre moyen d’allèle est de 17,8 ; 18,5 et 13,8 pour la belonging to the 3 breeds (100Barb, 100 Arab-Barb and 100 Arabian Pur-
population barbe, Arabe-Barbe et Pur-Sang Arabe, respectivement. Le taux Sang). The number of alleles per locus varying from 8 to 25, with an average
de l’hétérozygotie observée est de 0,675 ; 0,657 et 0,666 pour la race Barbe, number of alleles of 17.8; 18.5 and 13.8 for Barb, Arab-Barb and Arabian
Arabe-Barbe et Pur-sang Arabe, respectivement. Ces valeurs sont inférieures Pur-Sang, respectively. The observed heterozygosity (Ho) were 0.675; 0.657
à celles de l’hétérozygotie attendue : 0,786 ; 0,870 et 0,872, respectivement. and 0.666for Barb, Arab-Barb, andArabian Pur-Sang, respectively. These
Nos résultats ont montré que 91,2% de la variabilité génétique totale est values were lower than those of the expected heterozygosity (He). Our results
expliquée par la variation intra-population et que 8,8 % de cette variabilité showed that 91.2% of the total genetic variation were within the population
est attribuée aux différences entre les 3 populations. Le calcul de la distance and the remaining (8.8%) originates from difference among populations.
génétique a permis de constater que l’Arabe-Barbe était génétiquement plus Arab-Barb breeds was more closely related to Barb than Arabian-Pur-Sang.
proche des Barbes que des Pur- sang Arabes. Cette constatation est confirmée This finding was confirmed by the factorial correspondence analysis (AFC).
par l’analyse factorielle des correspondances (AFC). En conclusion, cette In conclusion, this study contributes to the programs of conservation and
étude contribue aux programmes de conservation et de valorisation des upgrading of local horse breeds, and reinforce the need to improve the
races chevalines locales, et insiste aussi à l’amélioration des méthodes de methods of management of equine breeding in Tunisia.
gestion de l’élevage équin en Tunisie.
Keywords: Horses, microsatellites, genetic diversity,
Mots-clés : Chevaux, microsatellites, diversité génétique, Tunisia
Tunisie
Introduction être phénotypiques (caractères morpho-biométriques),
biochimiques ou immunogénétiques (polymorphisme des
Les chevaux occupent une place de choix dans l’histoire, la protéines), cytogénétiques (nombre, formes et anomalies
culture et l’économie des pays d’Afrique du nord. En Tunisie, chromosomiques) ou moléculaires (analyse des marqueurs
il existe cinq différentes races telles que la Barbe (BA), l’Arabe- dans l’ADN). Durant les dernières décennies et avec le
Barbe (AB), le Pur-Sang Arabe (AR), le Pur-Sang Anglais (PS) développement de nouvelles technologies de la biologie
et le Poney de Mogods (PM) [19]. La race BA est spécifique moléculaire, les polymorphismes de l’ADN sont devenus les
de l’Afrique du Nord, alors que la race AR est d’origine marqueurs de choix dans l’étude de la structure génétique
orientale ; elle est introduite en Tunisie avec la conquête et l’histoire évolutive des organismes. Un certain nombre
arabo-musulmane. Le croisement de ses deux races a donné la de marqueurs sont à présent utilisés, mais les marqueurs
race AB [14]. Selon la Fondation Nationale d’Amélioration de les plus couramment employés dans l’étude de la diversité
la Race Chevaline (FNARFC), l’effectif total des chevaux est génétique sont les microsatellites [31]. Par définition, les
estimé à 37 000 têtes, dont la grande majorité de ces chevaux microsatellites sont des séquences d’ADN constituées de
sont communément identifiés comme BA et AB (presque répétition en tandem d’un motif de 1 à 6 pb [9, 35]. Ils sont
80% de l’effectif total) [15]. En raison de l’importance des largement utilisés dans l’analyse de la diversité génétique
races chevalines Tunisienne, une information complète entre différentes races animales : chevaline [5, 19,32],
sur la diversité génétique et la structure de la population bovine [2, 26], ovine [10, 16, 20], cameline [29, 30]. Dans
présente une grande importance puisqu’elle constitue la basse ce contexte, cette étude vise (i) étudier la diversité génétique
de tous programmes de conservation et de gestion durable de trois races chevaline élevées en Tunisie (BA, AB, AR) par
[28]. D’une manière générale, la caractérisation des races l’étude des marqueurs microsatellites et (ii) déterminer leur
animales fait recours aux plusieurs méthodes qui peuvent relation génétique.
Revue Méd. Vét., 2018, 169, 4-6, 103-109104 OULED AHMED (H.) AND COLLABORATORS
Matériel et méthodes de dénaturation initiale à 95°C pendant 10 minutes, suivie
de 30 cycles comprenant chacun une dénaturation à 95°C
ECHANTILLONNAGE ET EXTRACTION D’ADN pendant 30 secondes, une étape d’hybridation à 60°C durant
30 secondes et une étape d’élongation de 1 minutes à 72°C.
Au total, 300 animaux de l’espèce chevaline composés de Après les 30 cycles d’amplification, une étape d’élongation
100 BA, 100 AB et 100 AR ont été utilisés dans ce travail. finale a été effectuée à 72°C pendant 1 heure.
Les échantillons du sang ont été prélevés dans des tubes
EDTA (5 ml) à partir de la veine jugulaire. Ils ont été choisis L’amplifiât a subi une dénaturation à 95°C pendant 3
au hasard parmi ceux présents dans la banque de sang du minutes en présence du formamide, le produit PCR est
Laboratoire d’Analyses Génétiques Animales (LAGA) détecté par électrophorèse capillaire en utilisant le séquenceur
de l’Institut de la Recherche Vétérinaire de Tunisie ABI PRISM 3130 (Applied Biosystem, USA). La visualisation
(IRVT). L’extraction d’ADN génomique à partir du sang de résultat a été effectuée à l’aide du logiciel Gene Mapper
total a été réalisée à l’aide du Kit d’extraction (PurelinkTM V.4, 0 (Applied Biosystems, Foster City, Californie, USA). Le
Genomic DNA Mini Kit, Invitrogen, Carlsbad, CA, USA). standard de taille (GENESCAN-LIZ 500 Applied Biosystems)
a été utilisé pour déterminer la taille des allèles.
AMPLIFICATION DE L’ADN PAR PCR
CHOIX DES MARQUEURS MICROSATELLITES
L’amplification de l’ADN a été réalisée par une PCR
multiplexe en utilisant le Kit (Stocks Marks for Horses, Equine Un jeu de 17 marqueurs microsatellites a été choisi pour
Genotyping kit Applied Biosystems, Foster City, Californie, le génotypage des races chevaline tunisienne (Tableau I).
USA). Les conditions d’amplifications comportent une étape Ces marqueurs font partie de la liste recommandée par le
Locus Les séquences des amorces (avant et arrière) Taille (pb) Références
AHT4 5’: AACCGCCTGAGCAAGGAAGT 144-164 [7]
3’: CCCAGAGAGTTTACCCT
AHT5 5’: ACGGACACATCCCTGCCTGC 126-144 [7]
3’: GCAGGCTAAGGAGGCTCAGC
ASB2 5’: CCACTAAGTGTCGTTTCAGAAGG 216-250 [8]
3’: CACAACTGAGTTCTCTGATAGG
ASB17 5’: ACCATTCAGGATCTCCACCG 87-129 [8]
3’: GAGGGCGGTACCTTTGTACC
ASB23 5’: GAGGGCAGCAGGTTGGGAAGG 175-211 [21
3’: ACATCCTGGTCAAATCACAGTCC
CA425 5’: AGCTGCCTCGTTAATTCA 226-246 [12]
3’: CTCATGTCCGCTTGTCTC
HMS1 5’: CATCACTCTTCATGTCTGCTTGG 170-186 [17]
3’:TTGACATAAATGCTTATCCTATGGC
HMS2 5’:CTTGCAGTCGAATGTGTATTAAATG 222-248 [17]
3’: ACGGTGGCAACTGCCAAGGAAG
HMS3 5: CCATCCTCACTTTTTCACTTTGTT 148-170 [17]
3’: CCAACTCTTTGTCACATAACAAGA
HMS6 5’: GAAGCTGCCAGTATTCAACCATTG 151-169 [17]
3’: CTCCATCTTGTGAAGTGTAACTCA
HMS7 5’:TGTTGTTGAAACATACCTTGACTGT 165-185 [17]
3’: CAGGAAACTCATGTTGATACCATC
HTG4 5’: CTATCTCAGTCTTGATTGCAGGAC 127-139 [13]
3’: CTCCCTCCCTCCCTCTGTTCTC
HTG6 5’: GTTCACTGAATGTCAAATTCTGCT 84-102 [13]
3’: CCTGCTTGGAGGCTGTGATAAGAT
HTG7 5’: CCTGAAGCAGAACATCCCTCCTTG 118-128 [25]
3’: ATAAAGTGTCTGGGCAGAGCTGCT
HTG10 5’:TTTTTATTCTGATCTGTCACATTT 95-115 [25]
3’: CAATTCCCGCCCCACCCCCGGCA
LEX3 5’: ACATCTAACCAGTGCTGAGACT 142-164 [11]
3’: GAAGGAAAAAAAGGAGGAAGAC
VHL20 5’: CAAGTCCTCTTACTTGAAGACTAG 87-105 [34]
3’: AACTCAGGGAGAATCTTCCTCAG
Tableau I : Caractéristiques de 17 marqueurs microsatellites utilisés dans la caractérisation moléculaire de 3 races (Barbe (BA), Arabe-Barbe (AB) et Pur-
Sang Arabe (AR)) chevalines en Tunisie
Revue Méd. Vét., 2018, 169, 4-6, 103-109DIVERSITÉ GÉNÉTIQUES DES RACES CHEVALINES TUNISIENNE 105
Groupe Consultatif ISAG-FAO pour l’analyse de la diversité Résultats
génétique, l’identification et le contrôle de filiation des races
équines à l’échelle mondiale. VARIABILITÉ GÉNÉTIQUE INTRA-POPULATION
MÉTHODES D’ANALYSES STATISTIQUES Nombre total d’allèles et la richesse allélique
Le nombre total d’allèle (Na) calculé pour chaque race
Le nombre d’allèle par locus, la fréquence allélique,
est présenté dans le tableau II. Les 17 microsatellites utilisés
taux d’hétérozygotie observé (Ho) et attendu (He) sous
ont montré un polymorphisme génétique important avec
l’hypothèse de Hardy-Weinberg ont été calculés en utilisant
un nombre allélique variant entre 8 et 25. Dans notre étude,
le logiciel GENETIX version 4.05.2 [4] afin de déterminer la
le nombre total d’allèles est de l’ordre de 303, 315 et 234
diversité génétique intra-population.
respectivement pour les races BA, AB et AR.
La diversité génétique inter-population a été évaluée Fréquences alléliques et allèles privés
par le calcul des distances génétiques [27] et le coefficient
de différentiation génétique Gst de Nei (les indices de L’analyse de résultats montre une différence portant sur la
différenciation de Wright (ou Nei)) déterminés à l’aide du distribution allélique au niveau de chaque locus ainsi que sur
logiciel GENETIX 4.05.2 [4]. La représentation de la relation les fréquences des allèles dans chaque population. Au total,
génétique entre les populations testées a été réalisée par 115 allèles privés ont été identifiés dans le présent travail
l’approche AFC [22] en utilisant le même logiciel. pour les 3 races. Ces allèles sont répartis comme suit : 29
pour la race BA, 40 allèles privés pour la race AB et 46 allèles
privés pour la race AR, et (Tableau III). La plupart des allèles
BA AB AR
Locus
Na Na Na
VHL20 20,000 19,000 11,000
HTG4 15,000 15,000 9,000
AHT4 18,000 16,000 11,000
HMS7 20,000 19,000 15,000
HTG6 18,000 21,000 9,000
AHT5 11,000 17,000 12,000
HMS6 15,000 19,000 15,000
ASB23 20,000 17,000 19,000
ASB2 21,000 22,000 22,000
HTG10 21,000 23,000 14,000
HTG7 12,000 14,000 16,000
HMS3 19,000 19,000 16,000
HMS2 24,000 23,000 16,000
ASB17 25,000 25,000 12,000
LEX3 20,000 21,000 16,000
HMS1 11,000 13,000 8,000
CA425 13,000 12,000 13,000
Na 303 315 234
Nam 17,824 18,529 13,765
Na : Nombre total d’allèles, Nam : Nombre moyen d’allèles
Tableau II : Nombre total d’allèles et nombre d’allèles moyen par locus pour les trois races (Barbe (BA), Arabe-Barbe (AB) et Pur-Sang Arabe (AR))
chevalines en Tunisie
Race Nombre total d’allèles Nombre d’allèles privés % d’allèles privés
BA 303 29 9,57
AB 315 40 12,70
AR 234 46 19,66
Tableau III : Nombre et pourcentage d’allèles privés identifiés pour les 3 races (Barbe (BA), Arabe-Barbe (AB) et Pur-Sang Arabe (AR)) chevalines
Tunisienne étudiées.
Revue Méd. Vét., 2018, 169, 4-6, 103-109106 OULED AHMED (H.) AND COLLABORATORS
privés (104/115) ont caractérisés par une fréquence allélique génétique entre les populations. L’estimation de ces
inférieure à 2%. Par contre, 11 allèles spécifiques ont montré paramètres est représentée dans le tableau V.
une fréquence variant entre 2 à 12%, ce sont les allèles :
HTG4-138, ASB23-184, HMS-169 pour la race BA, ASB23- F statistiques
183, HTG7-130 pour la race AB et ASB23-200, ASB2-223,
Le tableau VI présente les valeurs de FST, FIS et FIT calculées
224 et 236, HTG10-118 et HTG7-122 pour la race AR.
entre les différentes races chevalines étudiées. La valeur
Taux d’hétérozygotie minimale de FST(0,029) est enregistrée pour la race AR. La
race BA présente la plus forte valeur de FST(0,041). Les trois
Les valeurs des taux d’hétérozygoties attendue He et populations présentent des indices de fixation (FIS) positifs
observée Ho sont présentées dans le tableau IV. D’après ce plus ou moins élevés compris entre 0, 1527 pour la race AR
tableau, aucune différence significative n’a été trouvée entre et 0,2465 pour la race AB, alors que la race Barbe prend une
les taux d’hétérozygoties (He et Ho). valeur intermédiaire de 0,2241.
VARIABILITÉ GÉNÉTIQUE INTER-POPULATIONS Distances génétiques
Paramètres de la diversité génétique de Nei (HT,HS, GST) Les distances génétiques calculées entre les 3 races
chevalines sont présentées dans le tableau VII. On constate
Les paramètres de la diversité génétique [27] ont été que les valeurs de la distance génétique entre les populations
déterminés afin d’étudier la structuration de la diversité BA et AB sont très faibles. Ceci indique que ces dernières
Population He Ho
BA 0,870 0,675
AB 0,872 0,657
AR 0,786 0,666
He : hétérozygotie attendue, Ho : hétérozygotie observée
Tableau IV : Taux d’hétérozygotie pour les 3 races (Barbe (BA), Arabe-Barbe (AB) et Pur-Sang Arabe (AR)) chevalines en Tunisie
Toutes les populations
Locus HS HT DST GST
AHT4 0,7945 0,8697 0,0751 0,0864
AHT5 0,8642 0,8994 0,0353 0,0392
ASB17 0,8568 0,8938 0,0371 0,0415
ASB2 0,8787 0,9223 0,0436 0,0473
ASB23 0,8545 0,9038 0,0493 0,0545
CA425 0,788 0,8323 0,0442 0,0531
HMS1 0,7735 0,8393 0,0658 0,0784
HMS2 0,8894 0,9266 0,0372 0,0401
HMS3 0,8877 0,9152 0,0275 0,0301
HMS6 0,8539 0,8955 0,0416 0,0464
HMS7 0,8936 0,9075 0,0140 0,0154
HTG10 0,3125 0,6563 0,3438 0,5238
HTG4 0,8071 0,8219 0,0148 0,018
HTG6 0,779 0,8545 0,0751 0,0879
HTG7 0,7191 0,7876 0,0685 0,087
LEX3 0,8787 0,9081 0,0294 0,0324
VHL20 0,4292 0,7127 0,2835 0,3978
Multilocus 0,7801 0,8557 0,0756 0,0884
HT : diversité totale ; HS : diversité intra-sous-population ;
DST : diversité entre sous-population ;
GST : coefficient de différenciation génétique entre les sous population
Tableau V : Indices de diversité génétiques de Nei M., 1978 [27] estimés pour les 17 microsatellites de trois races chevalines en Tunisie
Revue Méd. Vét., 2018, 169, 4-6, 103-109DIVERSITÉ GÉNÉTIQUES DES RACES CHEVALINES TUNISIENNE 107
sont très proches génétiquement, mais, elles sont éloignées AB se distingue par un nombre moyen d’allèles plus élevée.
à la population AR. Cette constatation est confirmée ensuite Ces différences de valeurs trouvées entres les races peuvent
par l’analyse factorielle des correspondances. être expliquées soit par le nombre des microsatellites utilisé,
le nombre d’individus analysé dans chaque étude, ou bien
Analyse factorielle des correspondances (AFC) par des particularités liées à la structuration des populations
équines étudiées.
Les relations génétiques entre les 3 populations étudiées
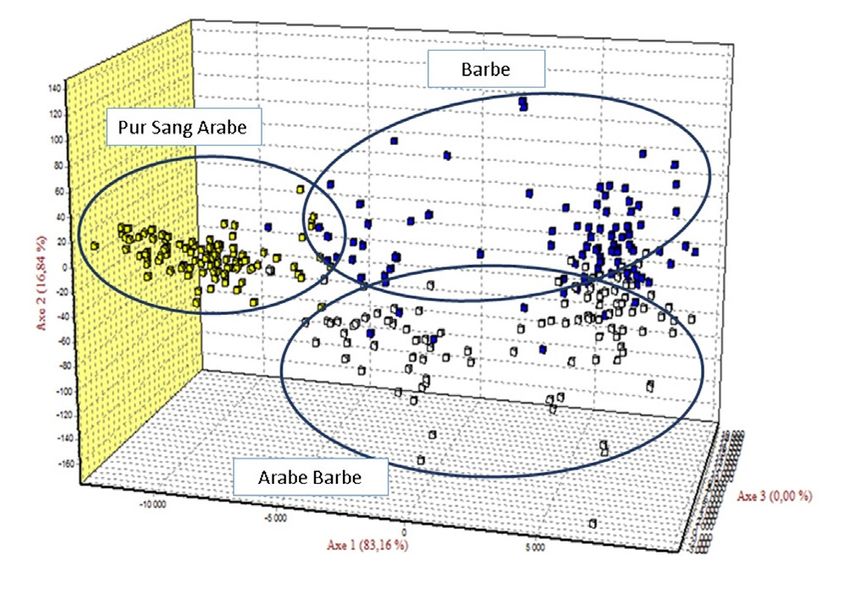
à partir d’AFC sont présentées dans la figure 1. On observe L’allèle privé est un indice d’originalité génétique est le
que le nuage de points représentant la population BA est plus nombre d’allèles présents dans une population à l’exclusion
proche des nuages de points de la population AB. de toute autre populations, ce qu’on appelle les allèles
« privés ». En utilisant ce paramètre on peut confirmer
l’appartenance ou non d’un individu d’origine inconnue à
une population [19]. Toute les races présentent des allèles
privés avec de pourcentages plus au moins variables. Les
taux d’hétérozygotie observés chez les races BA et AB sont
inférieurs aux taux rapportés par [19]. En utilisant ces mêmes
marqueurs microsatellites, ces derniers auteurs ont trouvé des
taux égales à 0,8 ± 0,03 et 0,8 ± 0,03 allèles, respectivement,
pour la race BA et la race AB. En comparaison avec cette
même étude, les taux d’hétérozygotie attendue obtenus chez
la population BA et AB sont comparables. Toutefois, des
taux relativement inferieurs ont été rapportés précédemment
dans une étude concernant la caractérisation moléculaire de
la race BA (0,046) et AB (0,039) moyennant 13 marqueurs
Figure 1 : Analyse Factorielle des Correspondances (AFC) des données microsatellites [18]. Malgré la richesse de la variabilité
moléculaires pour les 3 populations (Barbe (BA), Arabe-Barbe (AB) et
allélique, l’hétérozygotie attendue est supérieure à celle
Pur-Sang Arabe (AR)) chevalines en Tunisie
observée. Ceci montre un manque significatif d’hétérozygotes
qui pourrait être une conséquence du type de la gestion de
Discussion l’élevage.
Dans cet article nous avons réalisé une étude de La diversité génétique totale est la somme de la diversité
caractérisation génétique de 3 races de cheveux élevées en génétique intra-population et de la diversité génétique inter-
Tunisie par l’utilisation des microsatellites. Tous les loci sont population, la valeur Ht obtenue montre que 91,2% de la
polymorphes ce qui prouve l’efficience des microsatellites variabilité génétique totale est expliquée par la variation
utilisés dans l’analyse de la diversité génétique des populations intra-population et que 8,8 % de cette variabilité est attribuée
étudiées. Pour la race BA, le nombre moyen d’allèles de la race aux différences entre les 3 populations.
BA est proche à celui rapporté par [19], mais il est nettement
supérieur à celui observé sur une population de 26 chevaux de Les valeurs de Fst trouvées dans notre étude sont
la race Barbe tunisienne [18], de la race Barbe marocaine [1] inférieures à ceux rapporté dans une étude portant sur cinq
et également pour la race portugaise Sorraia [24]. Dans notre races de cheveux Indiens [3]. Cependant, elles sont très
étude et en comparaison avec les deux autres races, la race inferieur par rapport à ceux précédemment trouvées dans les
Population FST (3) FIS (1) FIT (2)
BA 0,041 0,2241 0,2559
AB 0,037 0,2465 0,2744
AR 0,029 0,1527 0,1773
FIS : différenciation des individus à l’intérieur des populations ;
FIT : différenciation des individus par rapport au total des populations ; FST : différenciation des populations par rapport au total
Tableau VI : Les indices de fixation des 3 races (Barbe (BA), Arabe-Barbe (AB) et Pur-Sang Arabe (AR)) chevalines tunisienne étudiées
BA AB AR
BA 0,000 0,090 0,775
AB 0,000 0,643
AR 0,000
Tableau VII : Distances génétiques de Nei (1978) entre les 3 populations (Barbe (BA), Arabe-Barbe (AB) et Pur-Sang Arabe (AR)) chevalines
Revue Méd. Vét., 2018, 169, 4-6, 103-109108 OULED AHMED (H.) AND COLLABORATORS
races polonaises ((FST = 10%, [37]), et les races brésiliennes 6. - BJORNSTAD G., GUNBY E., ROED K.H.: Genetic
(FST = 11,7%, [23]), ainsi que les races norvégiennes (FST structure of Norwegian horse breeds. J. Anim. Breed.
= 12%, [6]). Nos résultats indiquent, selon [36], une faible Genet., 2000, 117, 307–317.
différenciation génétique entre les 3 races chevalines étudiées 7. - BINNS M.M., HOLMES N.G., HOLLIMAN A., SCOTT
qui pourrait être dû à un régime de reproduction fermé ou à A.M.: The identification of polymorphic microsatellite
une sous structuration lors de l’échantillonnage. Les résultats loci in the horse and their use in thoroughbred parentage
d’analyse factorielle des correspondances trouvés dans testing. Br. Vet. J., 1995, 151, 9-15.
cette étude sont similaires à ceux rapporté dans des études 8. - BREEN M., LINDGREN G., BINNS M.M., NORMAN
précédentes [1, 18, 33]. J., IRVIN Z., BELL K., SANDBERG K., ELLEGREN
H.: Genetical and physical assignments of equine
En conclusion, il s’avère que l’utilisation d’un ensemble microsatellites-first integration of anchored markers in
de 17 marqueurs microsatellites est indispensable dans horse genome mapping. Mamm. Genome., 1997, 8 , 267-
l’étude de la diversité génétique. Les résultats de la variabilité 273.
génétique de 3 races chevalines tunisienne et les relations 9. - CHAMBERS G.K., MAC AVORY E.S.: Micro satellites,
entre eux fournissent des informations importantes sur leur consensus and controversy. Comparative Biochemistry
structuration génétique. Toutefois, l’évaluation de la relation and Physiology. Anim. Genet., 2000, 126, 455-476.
phylogénétique de ces 3 races révèle un taux d’homozygotie 10. - CIANI E., CIAMPOLINI R., D’ANDREA M.,
et une consanguinité élevée entre la race BA et AB. Donc, CASTELLANA E., CECCHI F., INCORONATO C.,
cette étude constitue une alerte pour établir des stratégies D’ANGELO F., ALBENZIO M., PILLA F., MATASSINO
de conservation et de gestion durable de races chevalines D., CIANCI D.: Analysis of genetic variability
tunisienne afin d’éviter la perte d’hétérozygotie. within and among Italian sheep breeds reveals
population stratification and suggests the presence of
Remerciements a phylogeographic gradient. Small. Rumin. Res., 2013,
112, 21-27.
Les auteurs remercient tous l’équipe du laboratoire 11. - COOGLE L., BAILEY E.: Equine dinucleotide repeat
«Gestion de la santé et de la qualité des productions animales, loci LEX064 through LEX070. Anim. Genet., 1998, 30,
référence LR14 Agr03» de l’Ecole National de Médecine 66-80
Vétérinaire Sidi Thabet, Tunisie et le Laboratoire d’Analyses 12. - EGGLESSTON-STOTT M.L., VALLE ADBAUTISTA
Génétiques Animales, Institut de la Recherche Vétérinaire de M., DILEANIS S., WICTUM E., BOWLING A.T.:
Tunisie qui ont financé la présente recherche, ainsi que tous Nine equine dinucleotide repeats at microsatellite loci
les personnels de Haras Sidi Thabet. UCDEQ136, UCDEQ405, UCDEQ412, UCDEQ425,
UCDEQ437, UCDEQ467, UCDEQ487, UCDEQ502
Références and UCDEQ505. Anim. Genet., 1997, 28, 370-383
13. - ELLEGREN H., JOHANSSON M., SANDBERG K.,
1. - AHMIDANI Y.: Le cheval barbe au Maghreb, analyses ANDERSSON L.: Cloning of highly polymorphic
génétiques et relations phylogénétiques, 137 pages, microsatellites in horse. Anim. Genet., 1992, 23, 133-142
Thèse Doc. Méd. Vét. Institut Agronomique et 14. - FERNANDEZ DE LA VEGA SEDANO J.: Analogies et
vétérinaire Hassan II, 2010. différences entre le barbe et l’espagnol. In. Tous les textes
2. - BACCOUCHE R., BEDHIAF S., HADDAD M., officiels sur le cheval barbe. Ed : Favre, Lausanne, Suisse,
JEMMALI B.: Population bovine locale en Tunisie : de 1989, pp.: 94-101.
la caractérisation morphologique à la caractérisation 15. - FNARC. : Fondation Nationale d’Amélioration de la
moléculaire. J. New. Sci., 2015, 14, 449-456. Race Chevaline, Rapport d’activité annuel, 2016.
3. - BEHL R., BEHL J., GUPTA N., GUPTA S.C.: Genetic 16. - GAOUAR S.B.S., KDIDI S., OURAGH L.: Estimating
relationships of five Indian horse breeds using population structure and genetic diversity of five
microsatellite markers. Animal., 2007, 1, 483-488. Moroccan sheep breeds by microsatellite markers. Small
4. - BELKHIR K., BORSA P., CHIKHI L., RAUFASTE Rumin. Res., 2016, 144, 23-27.
N., BONHOMME F.: GENETIX 4.05.2, Logiciel sous 17. - GUENIN G., BERTAND M., AMIGUES Y.:
Windows TM pour la génétique des populations. 2004., Characterization of seven new horse microsatellites:
Laboratoire Génome, Populations, Interactions, CNRS HMS1, HMS2, HMS3, HMS5, HMS6, HMS7, HMS8.
UMR 5000, Université de Montpellier II, Montpellier Anim. Genet., 1994, 25, 62
(France). 18. - HADDAD M.M., JEMMALI B., BEDHIAF A.,
5. - BERBER N., GAOUAR S., LEROY G., KDIDI S., BEDHIAF S., DJEMALI M.: Caractérisation moléculaire
TABET AOUEL N., SAIDI MEHTAR N.: Molecular des races chevalines autochtones en Tunisie. J. New. Sci.,
characterization and differentiation of five horse breeds 2014, 2, 11-20.
raised in Algeria using polymorphic microsatellite 19. - JEMMALI B., HADDAD M.M., OULED AHMED H.,
markers. J. Anim. Breed. Genet., 2014, 131, 387-394. LASFER F., BEN AOUN B., ESSAR S., KRIBI S., GTARI
Revue Méd. Vét., 2018, 169, 4-6, 103-109DIVERSITÉ GÉNÉTIQUES DES RACES CHEVALINES TUNISIENNE 109
S., EZZAOUIA M.H., REKIK B.: Investigation de la diversity of three Tunisian dromedary camel (Camelus
diversité génétique des races Barbe et Arabe Barbe en dromedarius) sub populations using microsatellite
Tunisie. J. New. Sci., 2015, 21, 948-956. . markers. Emir. J. Food Agric., 2015, 27, 362-366.
20. - KDIDI S., CALVO J.H., GONZALEZ-CALVO L., 30. - OULD AHMED M., BEN SALEM F., BEDHIAF S.,
BEN SASSI M., KHORCHANI T., YAHYAOUI M.H.: DJEMALI M.: Analyse moléculaire de la diversité
Genetic relationship and admixture in four Tunisian génétique des dromadaires (Camelus dromedarius) en
sheep breeds revealed by microsatellite markers. Small. Tunisie. Biotechnol. Agron. Soc. Environ., 2010, 14, 399-
Rumin. Res., 2015, 131, 64-69. 408.
21. - LEAR T.L., BRANDON R., BELL K.: Physical mapping 31. - ROGNON X., VERRIER E.: Caractérisation et gestion
of ten equine dinucleotide repeat microsatellites. Anim. des ressources génétiques. Les outils et méthodes de la
Genet., 1999, 30, 235 génétique pour la caractérisation, le suivi et la gestion
22. - LEBART L., MORINEAU A., WARWICK K.: de la variabilité génétique des populations animales.
Multivariate Descriptive Statistical Analysis. John UMR INRA/AgroParisTech « Génétique et Diversité
Wiley, New York, 1984, pp.: 231. Animales », 2007, Rabat.
23. - LIPPI A.S., MORTARI N.: Studies of blood groups 32. - RUKAVINA D., HASANBASIC D., DURMIC-PASIC
and protein polymorphisms in the Brazilian horse A., KALAMUJIC B., ZAHIROVIC A., RAMIC J.,
breeds Mangalarga Marchador and Mangalarga POJSKIC N.: Genetic Diversity of Arabian Horse
(Equuscaballus). Genet. Mol. Biol., 2003, 26, 431– 434. from Stud “Borike” (Bosnia and Herzegovina) Using
24. - LUIS C., JURAS R., OOM M.M., COTHRAN E.G.: Microsatellite Markers. R & R. J. Vet. Sci., 2016, 2, 21-25.
Genetic diversity and relationships of Portuguese and 33. - SEBTAOUI L., JEMMALI B., OULED AHMED H.,
other horse breeds based on protein and microsatellite SADFI A., HADDAD M.M., BEN ALAYA E., KAMOUN
loci variation. Anim. Genet., 2007, 38, 20-27. M., BEN GARA A., REKIK B.: Etude de la diversité
25. - MARKLUND S., ELLEGREN H., ERIKSSON S., génétique de races équine en Tunisie. Biotechnologie
SANDBERG K., ANDERSSON L.: Parentage testing Animale. 3ème Congrès International de Biotechnologie
and linkage analysis in the horse using a set of highly et Valorisation des Bio-Ressources (BVBR), 20 – 23
polymorphic microsatellites. Anim. Genet., 1994, 25, Mars 2015, Tabarka, Tunisia.
19-23. 34. - VAN HAERINGEN H., BOWLING A.T., STOTT
26. - MOAZAMI-GOUDARZI K., BELEMSAGA D.M.A., M.L., LENSTRA J.A., ZWAGSTRA K.A.: A highly
CERIOTTI G., LALOE D., FAGBOHOUN F., polymorphic horse microsatellite locus: VHL20. Anim.
KOUAGOU N.T., SIDIBE I., CODJIA V., CRIMELLA Genet., 1994, 25, 207.
M.C., GROSCLAUDE F., TOURE. S.M.: Caractérisation 35. - VIEIRA M.L.C., SANTINI L., MUNHOZ C.F.:
de la race bovine Somba à l’aide de marqueurs Microsatellite markers: what they mean and why they
moléculaires. Revue Élev. Méd. Vét. Pays trop., 2001, 54, are so useful. Genet. Mol. Biol., 2016, 39, 312-328.
129-138. 36. - WRIGHT S.: Evolution and the Genetics of populations.
27. - NEI M.: Estimation of average heterozygosity and Variability within and Among Natural Population,
genetic distance from a small number of individuals. University of Chicago Press, 1978.
Genetics+, 1978, 89, 583-590. 37. - ZABEK T., NOGAI A., RADKO A., NOGAI J., SLOTA
28. - NOTTER D.R.: The importance of genetic diversity in E.: Genetic variation of Polish endangered Bilgoraj
livestock populations of the future. J. Anim. Sci., 1999, horses and two common horse breeds in microsatellite
77, 61-69. loci. J. Appl. Genet., 2005, 46, 299–305
29. - NOUAIRIA G., KDIDI S., HAMMADI M.,
KHORCHANI T., YAHYAOUI M.H.: Assessing genetic
Revue Méd. Vét., 2018, 169, 4-6, 103-109Vous pouvez aussi lire

















































