Estimation de la contribution des différents stocks reproducteurs identifiés aux nourriceries chez le Flétan du Groenland Reinhardtius ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Estimation de la contribution des différents stocks
reproducteurs identifiés aux nourriceries chez le Flétan
du Groenland (Reinhardtius hippoglossoides)
Mémoire
Émilie Carrier
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Émilie Carrier, 2020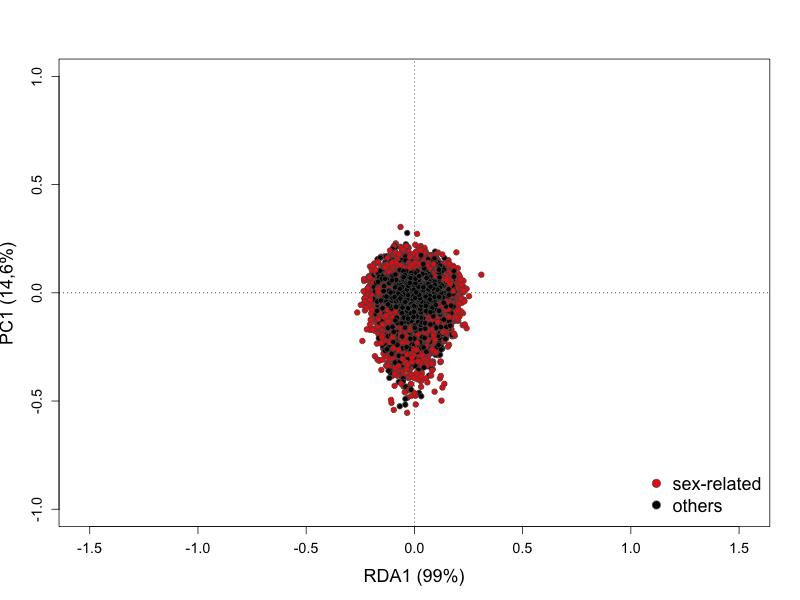
Estimation de la contribution des différents
stocks reproducteurs identifiés aux
nourriceries chez le Flétan du Groenland
(Reinhardtius hippoglossoides)
Mémoire
Emilie Carrier
Sous la direction de :
Louis Bernatchez, directeur de recherche
Pascal Sirois, codirecteur de rechercheRésumé
L'identification des stocks et la quantification de leur contribution relative au recrutement
sont des objectifs majeurs pour améliorer la gestion et la conservation des espèces
marines exploitées. Le séquençage de nouvelle génération permet d’analyser des milliers
de marqueurs génomiques et d’apporter la résolution nécessaire pour répondre à ces
questions chez les espèces marines qui sont faiblement différenciées. Le Flétan du
Groenland (Reinhardtius hippoglossoides) est un poisson plat largement exploité,
particulièrement dans le golfe du St-Laurent, au Canada. On y retrouve 2 nourriceries
connues, dont la contribution des juvéniles au renouvellement des différents stocks
adultes reste inconnue à ce jour. Les buts de ce projet sont de i) déterminer la structure
des populations de flétan du Groenland du Saint-Laurent et ii) d’estimer la contribution des
différents stocks reproducteurs identifiés aux différentes nourriceries. Pour ce faire, nous
avons échantillonné 100 juvéniles par nourricerie et 50 adultes de sites s’étendant du fjord
du Saguenay jusqu’au large de Terre-Neuve, avec quelques sites échantillonnés sur 2
années consécutives pour évaluer la stabilité temporelle de cette contribution. Nos
résultats montrent qu’après avoir retiré les marqueurs liés au sexe, la région de
l’Estuaire/Golfe du Saint-Laurent forme une population distincte de l’Atlantique près de
Terre-Neuve (Fst = 0.00146, p-value = 0.001). Les analyses d’assignation populationnelle
montrent que le recrutement dépend largement du stock du Saint-Laurent. Par ailleurs, on
retrouve une contribution variable du stock de Terre-Neuve, variant de 1% pour la
première année à 33% pour la deuxième, ce qui suggère un transport interannuel de
larves variable selon la force des courants profonds. Cette étude sert de modèle pour
l’identification des stocks pour les ressources halieutiques, dans un contexte où le milieu
marin offre peu de barrières à la dispersion, en plus de démontrer l’importance des
marqueurs liés au sexe et des répliques temporelles en génomique des populations.
iiAbstract
The identification of stocks and quantifying their relative contribution to recruitment are
major objectives toward improving the management and conservation of marine exploited
species. Next-generation sequencing allows to analyze thousands of genomic markers
which provide the resolution needed to address these questions in marine species with
weakly differentiated populations. Greenland Halibut (Reinhardtius hippoglossoides) is one
the most important exploited demersal species throughout the North Atlantic, and in
particular in the Gulf of St. Lawrence, Canada. There, two nurseries are known, the St.
Lawrence Estuary and the northern Anticosti Island, but their contribution to the renewal of
stocks remains unknown. The goals of this study were i) to document the genetic structure
and ii) to estimate the contribution of the different identified breeding stocks to nurseries.
We sampled 100 juveniles per nursery and 50 adults from seven sites ranging from
Saguenay fjord to offshore Newfoundland, with some sites sampled over two consecutive
years in order to evaluate the temporal stability of the contribution. Our results show that
after removing of sex-linked markers, the Estuary/Gulf of St. Lawrence represent a stock
which is genetically distinct from the Atlantic around Newfoundland (Fst = 0.00146, p-value
= 0.001). Population assignment showed that recruitment in both nurseries is largely
predominantly associated with the St. Lawrence stock. However, we found that the relative
contribution of both stocks to the nurseries is temporally variable with 1% contribution of
the Newfoundland stock one year but up to 33% for the second year, which may
hypothetically be caused by year-to-year variation in larval transport into the Gulf of St.
Lawrence. This study serves as a model for the identification of stocks for fisheries
resources in a context where few barriers to dispersal occurs, in addition to demonstrating
the importance of considering sex-linked markers and temporal replicates in studies of
population genomics.
iiiTable des matières
Résumé ................................................................................................................................... ii
Abstract .................................................................................................................................. iii
Table des matières ................................................................................................................. iv
Liste des figures ..................................................................................................................... vi
Liste des tableaux ................................................................................................................. vii
Remerciements .................................................................................................................... viii
Avant-propos ......................................................................................................................... ix
Introduction ............................................................................................................................ 1
Problématique générale ....................................................................................................... 1
La génomique des populations au service de la gestion des pêches ................................... 3
Structure des populations ................................................................................................... 3
Population et forces évolutives ........................................................................................... 4
Marqueurs moléculaires et méthode de séquençage .......................................................... 5
Méthodes de séquençage .................................................................................................... 8
Le Flétan du Groenland ...................................................................................................... 9
Biologie de l’espèce ............................................................................................................ 9
Pêche commerciale au Canada ........................................................................................ 10
Problématique ................................................................................................................... 11
Contexte du projet ............................................................................................................. 12
Objectifs ............................................................................................................................ 13
Chapter 1: ............................................................................................................................. 14
Résumé.............................................................................................................................. 15
Abstract ............................................................................................................................. 16
Introduction ....................................................................................................................... 17
Material and methods ........................................................................................................ 20
Sampling ........................................................................................................................... 20
DNA extraction ................................................................................................................. 22
Bioinformatics and sequencing ......................................................................................... 22
Identifying sex-linked and selected markers ..................................................................... 24
Population structure ......................................................................................................... 25
Assignment of juveniles ..................................................................................................... 26
Results ............................................................................................................................... 26
Genotype-By-Sequencing .................................................................................................. 26
ivSelection of SNPs .............................................................................................................. 27
F-statistics ......................................................................................................................... 27
Effect of sex-linked markers on population structure ....................................................... 28
Assignment of juveniles to identified stocks ...................................................................... 30
Discussion ......................................................................................................................... 33
Population structure: effect of sex-linked markers ........................................................... 33
Assignment of juveniles to identified stocks ...................................................................... 34
Comparison of SNPs and microhaplotypes ...................................................................... 35
Conclusion and perspective .............................................................................................. 36
Conclusion ............................................................................................................................ 38
Bibliographie ........................................................................................................................ 47
Annexes (supplementary materials) ..................................................................................... 58
vListe des figures
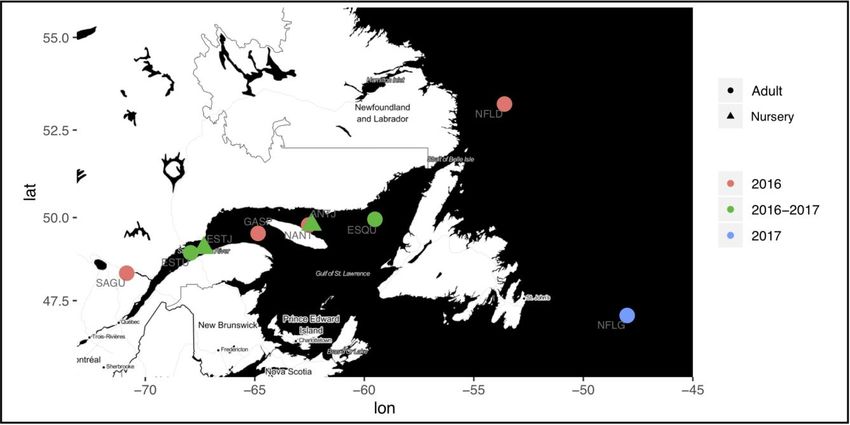
Figure 1. Map of sampling sites in Saguenay fjord (SAGU), St. Lawrence Estuary (ESTU,
ESTJ), Gaspé (GASP), North Anticosti Island (NANT, ANTJ), Esquiman (ESQU) and
Atlantic Ocean (NFLD, NFLG). The Estuary and northern Anticosti Island nurseries (ESTJ
and ANTJ) are defined by the triangles and adult populations of the St. Lawrence and
Atlantic (SAGU, NFLD, ESTU, NANT, GASP and ESQU) by circles. Sites that are sampled
for a single year are colored in pink for the first year of sampling and in blue for the
second. Those sampled for the two consecutive years are colored in
green……...……………………………….……………………………….………………………20
Fig 2. Population structure for Greenland Halibut from the Estuary/Gulf of St. Lawrence
(Saguenay fjord SAGU, St. Lawrence Estuary ESTU, St. Lawrence Estuary nursery ESTJ,
Gaspé GASP, North Anticosti Island NANT, North Anticosti Island nursery ANTJ and
Esquiman ESQU) and Newfoundland (NFLD and NFLG) using ADMIXTURE: a) for
dataset including sex-linked markers and b) for dataset without sex-linked markers.
Results are shown for K = 2. Sites are classified from west to east and individuals in
alphanumerical order. Sites sampled for two consecutive years are
combined…………….…………………………………………………………………………….28
Fig 3. Principal component analysis (PCA) with dataset including sex-linked markers.
Females are colored in pink and males in blue.………………………………………………29
Fig 4. Monte Carlo cross-validation test for SNPs dataset using different proportions of
training loci (0.1, 0.25, 0.5, 0.75 and 1) and individuals in training sets. Boxplots were
generated for overall sources populations (left) and for both populations separately (GSL
in center and NFL to the right). Selected SNPs were ranked in decreasing values of
differentiation index when using a different proportion then
1…………………………………………………………………………………………………….30
Fig 5. Individual assignment test using SNPs: a) for the first year and b) for the second
year, and for haplotypes: c) the first year of sampling and d) for the second year of
sampling to reference stocks of Gulf of St. Lawrence (GSL) and Newfoundland (NFL). All
loci and individuals were kept for the
test……………………………………..…………………………………………………….……..31
viListe des tableaux
Table 1. Number and percentage of remaining loci after every filtering step of the for: a)
SNPs and b) microhaplotype datasets…………………………………………………………23
Table 2. Pairwise differentiation index (Fst) of Greenland Halibut from 8 sampling sites
distributed from the St. Lawrence Estuary to offshore Newfoundland for 2 consecutive
years of sampling. Significant Fst (p-value < 0.05) are marked with an
asterisk……………………………………………………………………………….…………….27
viiRemerciements
J’aimerais tout d’abord remercier mon directeur de recherche, Louis Bernatchez, pour
m’avoir fait confiance dans la réalisation de ce projet et de m’avoir permis de développer
des compétences qui me seront utiles dans mon parcours professionnel, notamment en
communication scientifique. Je garde avec moi une expérience enrichissante du
laboratoire Bernatchez en raison de ses membres et de l’ambiance conviviale qui y règne.
Pour l’aide technique en laboratoire pour l’extraction et le séquençage de l’ADN, je
remercie particulièrement Alysse Perrault-Payette, Damien Boivin-Delisle et Louis
Deslauriers. Pour son support tout au long de ma maîtrise, tant pour résoudre des
problèmes techniques que l’interprétation des résultats, je remercie Anne-Laure Ferchaud.
Pour leur aide au niveau des analyses bio-informatiques, je remercie Éric Normandeau et
Yann Dorant. Je voudrais également remercier mon co-directeur, Pascal Sirois, pour
m’avoir apporté un retour à différentes étapes du projet.
Je remercie les professionnels et les pêcheurs volontaires sans qui le projet n’aurait pas
pu voir le jour, particulièrement Rénée Gagné et Céline Audet de l’Université du Québec à
Rimouski qui ont coordonné les opérations.
Je tiens à remercier Ressources Aquatiques Québec, qui est un regroupement de
chercheurs et d’étudiants interuniversitaire pour le développement durable de l’industrie
aquacole et des pêches commerciales et récréatives dans la région de Québec, pour
m’avoir donné l’opportunité de présenter lors des réunions annuelles, en plus de
m’accorder un soutien financier lors de voyages pour des communications scientifiques à
l’international.
Pour terminer, je voudrais remercier spécialement ma famille et mes amis, qui m’ont
toujours encouragée à poursuivre dans la voie professionnelle qui allait me rendre la plus
heureuse et accomplie, malgré toutes les épreuves pouvant survenir en cours de route.
viiiAvant-propos
Le projet qui a mené à la rédaction de ce mémoire a été financé par le Conseil de
recherches en sciences naturelles et en génie (CRSNG), le Fonds de recherche du
Québec – Nature et technologies (FRQNT) et par le regroupement interuniversitaire
Ressources Aquatiques Québec (RAQ).
Ce mémoire est essentiellement constitué d’un article « Estimating contribution of
reproductive stocks of Geenland Halibut (Reinhardtius hippoglossoides) to nurseries by
means of RADseq », qui a été soumis à la revue Evolutionnary Applications le 3 octobre
2019 et qui est en processus de révision par les pairs. Prochainement, ces connaissances
seront transférées dans la production d’un second article mettant en commun les récentes
avancés sur la migration des juvéniles, la connectivité transatlantique, ainsi que les effets
de la température et du sexe sur la croissance des juvéniles du Flétan du Groenland dans
tout l’est du Canada.
Je suis l’auteure principale de cet article. Il a été écrit en collaboration avec madame
Anne-Laure Ferchaud, monsieur Éric Normandeau, mon co-directeur Pascal Sirois et mon
directeur Louis Bernatchez. Pour le second article, je serai une des coauteurs.
Anne-Laure Ferchaud a participé aux analyses statistiques et à l’interprétation de celles-ci
et Éric Normandeau a effectué la préparation et la transformation des données bio-
informatique. L’échantillonnage a été réalisé par des équipes du ministère Pêches et
Océans Canada et des pêcheurs volontaires de la région, sous la coordination de Céline
Audet. J’ai réalisé les manipulations en laboratoire, analysé les données et interprété les
résultats avec l’assistance des différents coauteurs, ainsi que rédigé l’article en tant
qu’auteure principale. Tous les coauteurs ont participé à la révision du manuscrit.
ixIntroduction
Problématique générale
La pêche commerciale occupe une importance autant sur le plan économique, culturel et
politique pour divers peuples à l’échelle mondiale. En effet, de nombreuses communautés
côtières dépendent de cette industrie comme source principale de protéine et de revenus
(FAO, 2018). Les ressources halieutiques subissent plus que jamais des pressions
anthropiques, en raison de la surpêche, des captures accidentelles et des changements
climatiques.
Selon la définition du dictionnaire Larousse, une espèce est en surpêche lorsque le taux
d’exploitation est au-delà de son taux de renouvellement pour une période de temps
donné. On observe à ce jour plusieurs cas de pêcheries commerciales dont les
populations ont diminué drastiquement en quelques années, allant jusqu’au seuil de
l’extinction. Notamment, au Canada, on peut mentionner le cas de la morue de l’Atlantique
(Gadus morhua), espèce anadrome répartie dans tout l’est du Canada. La Morue est
composée de quatre stocks génétiquement distincts : les populations de l’Arctique, de
Terre-Neuve-et-Labrador, du Nord Laurentien et des Maritimes. Les scientifiques ont
observé une augmentation fulgurante du nombre d’effectifs dans tous les stocks (à
l’exception de celui de Terre-Neuve-et-Labrador) au début des années 80, ce qui a
influencé l’émission d’un nombre de permis et des quotas de pêche plus élevés dans les
zones exploitables (MPO, 2003). Par ailleurs, après 5 ans de pêche abondante, les stocks
ont chuté drastiquement : on note, par exemple, une diminution de 97% de l’abondance de
poissons (nombre d’individus matures) entre 1968 et 2001 pour le stock de Terre-Neuve-
et-Labrador. Ces observations imposent l’implantation d’un moratoire sur la pêche de la
Morue de l’Atlantique par les gestionnaires et l’interdiction d’utilisation d’engins mobiles
pour les poissons de fonds au début des années 90 (MPO, 2003). Cette diminution
importante du nombre d’effectifs a non seulement eu un impact au niveau de la taille des
différents stocks, mais également au niveau de leur composition phénotypique. On
observe un taux de croissance et un âge à la maturité moindres, ainsi qu’un taux de
mortalité supérieur après l’effondrement des populations (Smedbol et al. 2002).
1Tout comme la surpêche, les prises accidentelles causent des diminutions d’effectifs dans
les stocks. Celles-ci correspondent à l’ensemble des espèces non désirées (que ce soient
de mammifères marins, des oiseaux, des tortues marines, des poissons ou encore des
invertébrés), qu’elles soient d’intérêt commercial ou non, capturées lors des activités de
pêche (Hall, 1996). Bien que la plupart des méthodes de pêche aient le potentiel de
capturer des espèces non désirées (Northridge 1991), la pêche à la palangre et aux filets
maillants sont les deux techniques les plus dommageables selon une méta-analyse des
pêches commerciales situées sur la côte est américaine (Zollett 2009). L’ampleur des
effets néfastes à l’échelle écosystémique dépend de la quantité d’individus récoltés, du
statut de vulnérabilité (particulièrement pour les espèces longévives ayant un temps de
génération élevé) et du rôle dans la chaîne alimentaire des espèces accidentellement
visées (Lewison et al., 2011). À une échelle plus locale, les conséquences des prises
accidentelles pour les pêcheurs comprennent le bris d’équipement, la perte monétaire, en
plus de potentiellement nuire à leur sécurité (dans les cas où l’animal piégé peut être
dangereux). Pour les individus capturés, la pêche peut résulter en des blessures
irréversibles, voire mortelles.
Selon les plus récentes statistiques citées par le Groupe d’experts intergouvernemental
sur le climat (GIEC), si le développement se maintient à un niveau similaire, on prévoit
qu’à chaque cycle de 30 ans, la température de l’air augmentera entre 0,3 et 0,7 degré
Celsius (Kirtman et al. 2013). Les conséquences des changements climatiques sur les
milieux marins peuvent se classer en deux catégories : les changements physiques
directs, ainsi que les réponses biologiques et écologiques aux changements physiques
directs (FAO, 2016). On compte parmi les conséquences physiques directes
l’augmentation du dioxyde de carbone atmosphérique (qui contribue à l’acidification des
océans), l’augmentation de la température de l’eau (qui influence la modification de l’aire
de répartition d’espèces aquatiques, en plus de causer une hausse de la mortalité) et
l’augmentation de la fréquence d’évènements climatiques extrêmes (qui peuvent
contribuer à la destruction d’habitats côtiers et rendent plus risquées les pêches en haute
mer) (FAO, 2016). Pour ce qui est des impacts indirects des changements climatiques, il
est plus difficile d’en prédire les conséquences puisqu’il est plus difficile de les tester en
milieu contrôlé (FAO, 2016).
Tous ces facteurs risquent de modifier l’équilibre entre le taux de mortalité et le taux de
renouvellement des espèces exploitées, ce qui causerait un déclin des effectifs (FAO,
22012). Pour contrer cette problématique, il est primordial d’établir un plan de gestion
adapté aux caractéristiques biologiques de l’espèce en question, des mutations de son
milieu, ainsi que de son taux d’exploitation.
La génomique des populations au service de la gestion des
pêches
Structure des populations
La définition d’une population et la détermination de sa structure génétique revêtent d’une
importance particulière dans le cadre de mise en œuvre d’un plan de gestion. Cela permet
de comprendre les causes écologiques de la structuration lorsqu’on met en relation ces
informations avec des renseignements écologiques, d’en optimiser l’exploitation (dans le
cas des ressources naturelles) et de prédire les effets d’une perturbation sur l’espèce en
question. Une population correspond à un ensemble d’individus vivant à une proximité
suffisante pour que tous aient une probabilité de rencontre égale à des fins de
reproduction (Waples et Gaggiotti, 2006). Lorsque ce concept est appliqué au domaine de
la gestion des pêches, on emploie plutôt le terme de stocks. Tout comme une population,
un stock répond à des contraintes spatiales et reproductrices; les individus d’un même
stock doivent avoir des traits d’histoire de vie similaires et doivent présenter un certain
isolement reproducteur (Hilborn et Walters, 2013). Par contre, la distinction majeure entre
ces concepts réside dans leur application. On définit un stock dans le but de mettre en
place une politique de gestion pour préserver l’espèce exploitée à une échelle plus
appropriée que celle de l’espèce (Begg et al. 1999). Différentes approches permettent
d’identifier les stocks ou les populations d’une espèce. On peut les diviser en différentes
catégories, soit la capture-recapture, les données de débarquements, les traits d’histoire
de vie, la composition et de l’abondance des parasites, la chimie des otolithes, les traits
morphologiques et la génétique (Begg et al. 1999). Néanmoins, selon Elliott et
collaborateurs (1995) la combinaison de différentes approches permet d’augmenter la
fiabilité de l’inférence des unités de gestion.
3Population et forces évolutives
Il est possible d’étudier la structure des populations, c’est-à-dire la façon dont la variation
génétique est distribuée au sein d’un ensemble d’individus, en fonction des forces
évolutives qui agissent sur celle-ci, soit la dérive génétique, la mutation, la migration et la
sélection. Les forces évolutives, qu’elles soient neutres (dérive génétique, mutation et
migration) ou non (sélection), ont une action différente sur la diversité génétique. Parmi
toutes ces forces, la migration (ou la dispersion) a des conséquences importantes sur la
dynamique démographique des populations observées. On peut définir la dispersion
comme étant le mouvement d’individu(s) d’une population vers une autre de façon
permanente ou suffisamment longue pour contribuer à enrichir le patrimoine génétique de
la nouvelle population (Lowe et Allendorf 2010). Cette dispersion peut soit être passive
(provoquée par divers facteurs environnementaux) (Bohonak et Jenkins 2003) ou active
(mouvement volontaire d’individus) (Van Dyck et Baguette 2005). La dispersion est
bénéfique lorsque les coûts liés au déplacement et à l’acclimatation au nouvel habitat sont
inférieurs aux bénéfices projetés en termes d’augmentation de succès reproducteur (Cote
et al. 2010). Les populations d’une espèce sont panmictiques lorsque l’espèce est
composée d’une seule population génétique (ou métapopulation), c’est-à-dire qu’il y a
assez de migration entre les « sous-populations » pour observer la même structure
génétique chez chacune d’elles (Waples et Gaggiotti, 2006). Au contraire, lorsqu’il n’y a
pas de flux génétique entre les populations d’une même espèce (autrement dit, il n’y a pas
de dispersion), les populations sont isolées les unes des autres. Il existe une gamme de
variation de dépendance démographique entre ces 2 états en fonction du taux de
dispersion entre les sous-populations.
Par ailleurs, la mesure directe du taux de dispersion comporte des difficultés techniques et
logistiques. Il est particulièrement difficile de suivre les espèces de petite taille, les
espèces vivant dans des habitats ouverts et celles qui sont très mobiles (Dingle et Drake
2007). De plus, comme la durée de vie des batteries est limitée, il n’est parfois pas
possible de monitorer le mouvement d’individus jusqu’au moment de la dispersion
effective. C’est particulièrement le cas pour des espèces ayant une longévité importante et
où les mouvements vers une population différente se font occasionnellement (Moore et al.
2017). Les méthodes indirectes impliquent, quant à elles, l’utilisation de marqueurs
4moléculaires (Paetkau et al. 1995, Rannala et Mountain 1997, Hellberg et al. 2002,
Waples et Gaggiotti 2006). Les outils moléculaires offrent une possibilité de mieux
répondre à des questions écologiques, évolutives et de conservation, dans un contexte
d’évolution dynamique des populations naturelles (Manel et al. 2005). Ils permettent
notamment d’identifier les migrants, sans avoir besoin de suivre les déplacements
d’individus marqués. Autrement dit, il est possible de déterminer la proportion les migrants
présents dans les populations par leurs différences génétiques qui les distinguent de la
nouvelle population dans laquelle ils se trouvent (Paetkau et al. 1995, Waser et Strobeck
1998). L’ensemble des méthodes qui utilisent les informations génétiques pour déterminer
l’appartenance d’un individu à une population se nomme l’assignation populationnelle.
Celle-ci permet de répondre à plusieurs questions biologiques, telles que l’origine d’un
individu, la structure d’une population, le taux de dispersion et le taux d’hybridation (Waser
et Strobeck 1998, Manel et al. 2005). La puissance d’assignation est dépendante du
niveau de différenciation des sous-populations étudiées, du nombre de marqueurs utilisés
et de la taille d’échantillonnage (Cornuet et al. 1999). Plus les populations sont similaires,
plus le nombre de marqueurs est faible ou plus l’échantillon est petit, moins grande sera la
puissance du test statistique. Ainsi, il devient difficile de faire la distinction entre un faible
niveau de différenciation causé soit par une population de grande taille effective (dérive
génétique) et une grande dispersion, ou soit par une divergence récente (Allendorf et al.
2010). Ces outils d’assignation sont particulièrement utiles dans le domaine de la gestion
des ressources halieutiques. Comme il est difficile de suivre la dispersion des larves et
des œufs, l’assignation des juvéniles à un stock adulte est problématique (Begg et al.
1999). De plus, chez la plupart des espèces marines, des contraintes font en sorte que les
distributions des différentes unités de conservation sont continues. Elles incluent un
nombre réduit de barrières à la dispersion en milieu marin, la phase larvaire pélagique qui
provoque un flux génique important et des tailles de population élevées (Gagnaire et al.
2015, Riginos et al. 2016). Il devient alors difficile de faire une gestion individuelle de ces
entités (Waples et al. 2008).
Marqueurs moléculaires et méthode de séquençage
Le choix du type de marqueur utilisé pour les études de génétique des populations dépend
principalement de trois critères principaux : le niveau de polymorphisme, la facilité de
5générer les données, ainsi que le coût par individu. Les Polymorphismes de Nucléotide
Simple ou Single Nucleotide Polymorphism (SNP) sont des marqueurs bi-alléliques
répartis sur l’ensemble du génome (Collins et al. 1997) et correspondent à ce jour au
marqueur de choix en génomique des populations. Avec le développement du
séquençage de nouvelle génération, ces marqueurs sont de plus en plus utilisés dans
divers domaines de recherche en raison de leur polyvalence en génétique des populations
(Andrews et al. 2016, Ekblom et Galindo 2011, McCormack et al. 2013, Narum et al.
2013), ainsi que leur faible coût de développement (De Wit et al. 2015). On retrouve de
nombreux exemples dans la littérature de détection de structure de population chez des
espèces marines à l’aide de ce type de marqueur. Les espèces marines concernées
incluent le Homard d’Amérique (Homarus americanus) (Benestan et al. 2015), le
Holothurie rouge de Californie (Parastichopus californicus) (Xuereb et al. 2018) et la
Morue de l’Atlantique (Gadus morhua) (Barth et al. 2017). Par exemple, l’étude de
Benestan et collaborateurs (2015) a démontré la présence d’une différenciation
significative entre le nord et le sud de l’aire de répartition du Homard d’Amérique
(Homarus americanus). Bien que la puissance d’analyse soit suffisante pour de
nombreuses espèces aquatiques, les SNPs peuvent s’avérer impuissants pour résoudre la
structure de population d’espèces marines. En effet, les contraintes du cycle de vie, qui
sont répandues chez de nombreuses espèces, limitent la différenciation. On observe un
taux de dispersion élevé, en raison principalement de barrières physiques modérées, ainsi
que de grandes tailles de populations (Gagnaire et al., 2015). Par ailleurs, il est possible
de combiner plusieurs SNPs en déséquilibre de liaison, ce qui permet d’augmenter le
nombre de génotypes. Cette approche multi-SNPs se nomme micro-haplotypes (Kidd et
al. 2013, 2014). En effet, lorsqu’on analyse différents SNPs séparément, la probabilité
d’obtenir au hasard la même valeur de SNPs entre 2 individus (et ainsi inférer une même
unité populationnelle) est plus élevée que la combinaison des plusieurs SNPs. L’utilisation
des micro-haplotypes a comme avantage de ne nécessiter aucune méthode de
séquençage particulière, de demander peu de traitement bio-informatique pour faire la
transformation, en plus d’être compatible avec les logiciels d’assignation populationnelle
qui peuvent traiter les microsatellites. Plus particulièrement dans le domaine des pêches
commerciales, on retrouve deux exemples récents qui illustrent l’utilité de ces micro-
haplotypes pour répondre à une problématique de conservation. Dans leur étude publiée
en 2017, McKinney et collaborateurs ont comparé la puissance des marqueurs SNPs et
des micro-haplotypes pour déterminer la contribution relative des différents stocks chez le
6Saumon quinnat (Oncorhynchus tshawytscha) aux différentes populations de l’ouest de
l’Alaska. Pour les groupes les plus étroitement liés, les auteurs ont démontré que les
micro-haplotypes composés de plusieurs SNPs augmentent la précision des analyses de
stock mixtes de 7% à partir d’un total de 500 marqueurs. Celle-ci monte jusqu’à 17% pour
l’assignation d’individus inconnus à leur population d’origine. Il est également possible
d’utiliser les micro-haplotypes afin de performer une analyse l’assignation parentale de
larves d’origine inconnue à un couple de géniteurs potentiels. Ce type d’analyse peut être
employé notamment pour estimer le taux de dispersion des juvéniles des espèces qui sont
sédentaires à maturité (Christie 2010). De plus, cette méthode permet de détecter les
mouvements récents, plutôt que le flux génique historique, que fournit l’indice de
différenciation. C’est ce qu’a réalisé l’équipe de Baetscher et collaborateurs (2019) avec le
Sébaste du varech (Sebastes atrovirens), une espèce résidante des milieux côtiers de
Californie et dont une partie de l’aire de répartition est intégrée dans plusieurs aires
marines protégées. Sur les 6000 échantillons récoltés, ils ont pu identifier significativement
8 familles (couple et rejeton), en addition à 25 fratries à l’aide de 96 micro-haplotypes
hautement différenciés. Avec ces groupements, ils ont découvert qu’une partie de celles
qui naissent à l’intérieur d’une certaine zone protégée peut migrer en fonction des
courants marins pour s’établir soit dans une autre zone protégée à proximité, ou encore
dans des régions où la pêche est permise.
En addition au choix du type de marqueur, il est important de sélectionner les régions
génomiques à analyser. Particulièrement pour l’étude de populations d’espèces marines,
une étude de Benestan et collaborateur (2015) a démontré qu’une attention particulière
doit être mise sur les marqueurs associés au sexe. Ces marqueurs sont présents sur les
chromosomes sexuels (pour les espèces qui en possèdent) ou dans une région associée
à l’expression de gènes impliqués dans la différenciation sexuelle pour des traits
morphologiques, physiologiques et/ou comportementaux (Bell 1982), chez les espèces
ayant un déterminisme génétique sur la différenciation entre les sexes (Gamble et
Zarkower 2014). Ils peuvent à eux seuls amplifier, voire créer un signal de structuration
recherché ou le masquer si le ratio entre le nombre de mâles et de femelles n’est pas
balancé, surtout pour des espèces possédant un faible niveau de variation génétique. Par
exemple, chez le Homard d’Amérique (Homarus americanus), seulement 12 SNPs (sur un
total de 1717) peuvent créer une structuration significative (Fst moyen de 0,0800 pour les
marqueurs liés au sexe, comparativement à 0,0030 pour les autres) entre le milieu côtier
7et le milieu marin, en manipulant le nombre d’individus de chaque sexe dans ces deux
groupes (Benestan et al. 2015). À ce jour, peu d’études se sont intéressées à l’influence
des marqueurs associés au sexe pour répondre à des problématiques de biologie de la
conservation (Jackson et al., 2018, Messmer et al. 2018, Ogata et al., 2018).
Méthodes de séquençage
L’amélioration des méthodes de séquençage de nouvelle génération depuis le milieu des
années 2000 est un tournant dans la génétique des populations, en permettant de définir
des différenciations subtiles à des échelles locales fines (Metzker 2010). Bien qu’il soit
maintenant possible de séquencer l’entièreté du génome de toutes les espèces, il devient
fastidieux et dispendieux d’utiliser cette pratique pour génotyper des centaines ou des
milliers d’individus pour déterminer la structure de populations. Parmi toutes les approches
de séquençage, le « Génotype par Séquençage » (GBS) permet d’analyser des milliers de
SNPs chez plusieurs organismes à la fois (Davey et al. 2011). Le GBS a déjà été utilisé
pour de nombreux projets de génétique des populations, et, selon certains auteurs, cette
méthode a révolutionné les études dans le domaine par sa facilité d’utilisation et sa
polyvalence (Narum et al. 2013). Le protocole du GBS peut se diviser en trois grandes
étapes décrites dans le papier d’Elshire et al. (2011). La première étape est d’extraire
l’ADN et de distribuer les échantillons dans les puits d’une plaque. La deuxième étape
consiste à préparer des librairies de séquençage. Des enzymes de restriction sont
ajoutées afin de fragmenter le génome. Les endonucléases choisies sont non-spécifiques
pour créer des milliers de morceaux d’ADN génomique de toutes les longueurs. Des
séquences code-barres de quelques nucléotides qui sont spécifiques aux individus sont
ajoutées afin de les distinguer subséquemment. Les échantillons sont groupés et amplifiés
par PCR. La troisième étape au protocole est de séquencer les échantillons et identifier le
génotype. Il existe quelques variantes au protocole selon le nombre d’enzymes de
restriction utilisées et le moment auquel ils sont incorporés aux échantillons, la méthode
de sélection de fragments selon leur taille et le moment dans le protocole où sont
incorporés les adaptateurs utilisés pour se fixer à la matrice de séquençage (Li et Wang,
2017). Le principal avantage de cette méthode est de réduire la charge de matériel
génétique à séquencer, puisque l’étape de la PCR amplifie les fragments ayant une
longueur optimale. Cela permet de limiter le temps et les coûts des projets, d’autant plus
8qu’il n’est souvent pas nécessaire de séquencer l’ensemble du génome. Le GBS offre
également la possibilité de séquencer des espèces qui ne sont pas des modèles
biologiques, ce qui s’avère souvent le cas dans les études de génétique de populations
(Narum et al. 2013).
Le Flétan du Groenland
Biologie de l’espèce
Le flétan du Groenland (Reinhardtius hippoglossoides) est un poisson plat de la famille
des Pleuronectidae. Son aire de répartition est circumpolaire et l’espèce se retrouve à la
fois dans l’océan Atlantique et Pacifique. Dans l’océan Atlantique, l’espèce s’étend depuis
la mer de Barents à l’est, jusqu’à la terre de Baffin à l’ouest et à au sud jusqu’en Nouvelle-
Écosse (Bowering et Brodie 1995). On retrouve le Flétan du Groenland à des profondeurs
moyennes de 200 m jusqu’à 375 m dans le golfe du Saint-Laurent (MPO 2019) et ses
températures préférentielles varient de 1,3 à 2,7 °C (Peklova et al. 2012). Dans le golfe
du Saint-Laurent, la fraie a lieu l’hiver durant les mois de janvier jusqu’à mars. Pour les
poissons plats bentho-pélagiques tels que le flétan du Groenland, on observe un niveau
élevé de dispersion en raison de la phase larvaire pélagique prolongée, ce qui peut
conduire à une panmixie de la population (Bailey 1997, Hoarau et al. 2002). Les juvéniles
se retrouvent en majorité dans l’estuaire du Saint-Laurent et au nord de l’île d’Anticosti
regroupés dans deux nourriceries (MAPAQ 2014). Une nourricerie correspond à un
rassemblement de juvéniles (poissons ayant franchi le stade larvaire, mais dont les
organes génitaux ne sont pas encore développés), dans un lieu où la densité est
supérieure à la moyenne de l’aire de répartition de l’espèce. Cet habitat leur permet de s’y
nourrir et de se développer pour atteindre la forme adulte (Beck et al. 2001, Dahlgren et al.
2006). Hors du golfe du Saint-Laurent, seules deux autres nourriceries ont été identifiées
à ce jour. La première est située plus au nord, dans le détroit de Davis et aurait pour
origine les poissons adultes de la baie de Baffin et du nord-ouest du Groenland
(Jørgensen et Arboe 2013). La deuxième serait située dans la mer de Svalbald, au large
des côtes norvégiennes (Albert et Vollen 2014). Dans le golfe, cette espèce est sujette à
des modifications considérables de son environnement qui affectent l’état des stocks. En
raison des changements climatiques, les valeurs moyennes de température de l’eau sont
9à la hausse. En effet, on a observé une valeur moyenne inférieure à 6°C dans les régions
du centre du golfe et du chenal Esquiman en 2013 (Galbraith et al. 2017). Une des
conséquences entraînée par la hausse des températures de l’eau est la diminution de la
quantité d’oxygène dissous. Bien que cette espèce soit relativement résistante à l’anoxie
(entre 18% et 25% en moyenne, Gilbert et al. 2005), il a été démontré que l’hypoxie
affecte les juvéniles (Dupont-Prinet et al. 2013). Une équipe de Rimouski a démontré que
les adultes ont un seuil critique d’oxygène significativement supérieur à celui des juvéniles
à une température de 5°C, qui simule les valeurs actuelles approximatives (11% de
saturation chez les individus matures contre 15% chez les juvéniles). De plus, ces mêmes
auteurs ont étudié les effets de la rareté en oxygène sur le métabolisme. Ils ont observé
chez les juvéniles une baisse du métabolisme, qui a pour conséquence le ralentissement
de la digestion et une réduction approximative de 58% du métabolisme aérobie. Ces
résultats suggèrent que l’hypoxie dans le golfe pourrait affecter négativement le
recrutement et ainsi menacer à moyen ou à long la pêche commerciale (Dupont-Prinet et
al. 2013). Présentement, la gestion dans le Saint-Laurent est divisée en trois régions
distinctes : l’ouest du golfe (avec ou sans l’estuaire), le nord de l’île d’Anticosti et le chenal
Esquiman, qui sont soumises à des dynamiques environnementales ou des pratiques de
pêches différentes (MPO 2010).
Pêche commerciale au Canada
Depuis le déclin de la Morue de l’Atlantique dans les eaux canadiennes, en plus des
baisses du nombre d’effectifs chez plusieurs espèces de poissons plats (NAFO, 1994),
l’exploitation du flétan du Groenland a été le sujet de plusieurs questionnements,
particulièrement en raison de son impact économique. Les industries de la pêche et de la
transformation, particulièrement dans les régions du golfe du Saint-Laurent, de Terre-
Neuve et de l’Arctique, bénéficient considérablement de l’abondance du Flétan du
Groenland dans les eaux canadiennes. Les débarquements de flétan du Groenland
représentaient 39,8% du tonnage de poissons de fonds pour la province de Québec et
23,8% pour la province de Terre-Neuve et du Labrador en 2018. En raison du prix élevé
de sa chair et de son abondance, cette pêche représente plus de 50% de la valeur des
pêches commerciales de poissons de fond pour ces deux régions (DFO, 2019).
10Problématique
Malgré l’importance de cette espèce pour l’économie canadienne, on connaît peu sur son
écologie de même que sur sa structure des populations. Par ailleurs, plusieurs auteurs
affirment que la sous-population de flétan présente dans le Saint-Laurent serait isolée des
stocks de l’Atlantique. Les évidences proviennent d’études portant sur le polymorphisme
d’allozymes (Fairbairn, 1981) et sur l’abondance et la diversité des parasites (Khan et al.
1982, Arthur et Albert 1993). Fairbairn (1981) a estimé que le stock du Nord-Est de
l’Atlantique, représenté par un site près des côtes ouest islandaises et un dans la mer de
Barents, serait isolé des stocks du Nord-Ouest en raison de fréquences d’allèles uniques à
un locus pour les poissons provenant de la mer de Béring. Également, une différence
observée à deux allozymes entre les stocks du nord-ouest de l’Atlantique et le golfe du
Saint-Laurent suggère qu’ils sont partiellement isolés. Une autre équipe a obtenu des
résultats similaires en étudiant deux protozoaires vivant dans le sang des flétans (Khan,
1982). Selon les occurrences d’infections par les différentes espèces de parasites, ils ont
conclu qu’il existe trois stocks indépendants dans l’ouest : celui du Nord-Ouest, celui du
Sud-Ouest de Terre-Neuve dans la baie Fortune et celui du golfe du Saint-Laurent. Arthur
et collaborateurs (1993) se sont intéressés aux flétans du Groenland de cours d’eau à
proximité du golfe pour déterminer s’il y a présence d’une structure à une échelle plus
locale. La présence significativement plus importante de certains parasites dans les
flétans du Saint-Laurent contrairement à ceux du fjord du Saguenay et au stock de
l’Atlantique les distingueraient de ces derniers. Leurs résultats suggèrent donc que tout le
cycle vital des flétans du Saint-Laurent se produit à l’intérieur du golfe. Plus récemment
Roy et collaborateurs (2014) ont démontré à l’aide de marqueurs microsatellites que la
population du golfe serait plutôt panmictique et que cette population comprendrait aussi
les flétans retrouvés à l’Est du Labrador et près de Terre-Neuve. Ils proposent comme
explication que la différenciation locale observée provient de nouvelles recrues et de
colonisateurs issus du même fond génétique, ce qui est en accord avec les résultats
précédents qui montrent une différenciation à grande échelle. Bien que ces résultats
apportent une résolution plus locale de la connectivité, ils ne permettent pas de démontrer
avec certitude l’existence d’une structure des stocks de flétan du Groenland à l’intérieur du
Saint-Laurent. Cela permet donc de se questionner quant à la présence d’une telle
11Vous pouvez aussi lire

















































