Différenciation génétique et sélection chez le cerf de Virginie (Odocoileus virginianus) introduit à l'île d'Anticosti - Mémoire Jérémie Fuller ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Différenciation génétique et sélection chez le cerf de
Virginie (Odocoileus virginianus) introduit à l’île
d’Anticosti.
Mémoire
Jérémie Fuller
Maîtrise en biologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Jérémie Fuller, 2018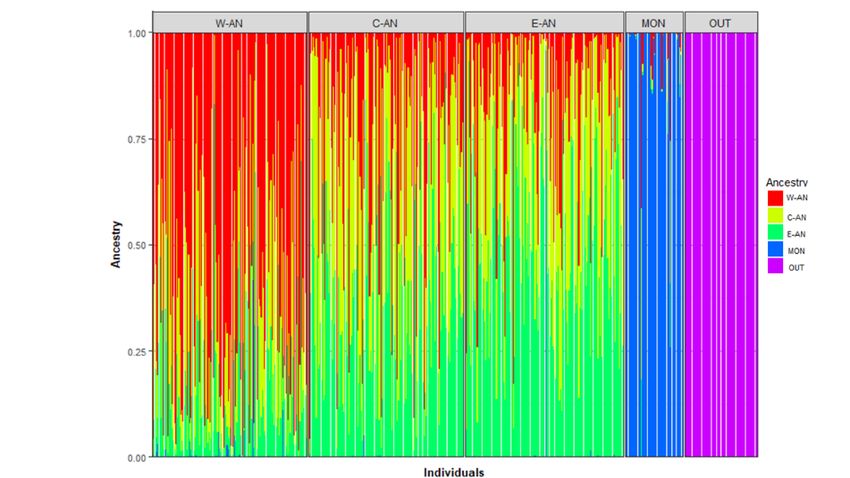
Différenciation génétique et sélection chez le cerf de
Virginie (Odocoileus virginianus) introduit à l’île
d’Anticosti.
Mémoire
Jérémie Fuller
Sous la direction de :
Steeve D. Côté, directeur de recherche
Louis Bernatchez, codirecteur de recherche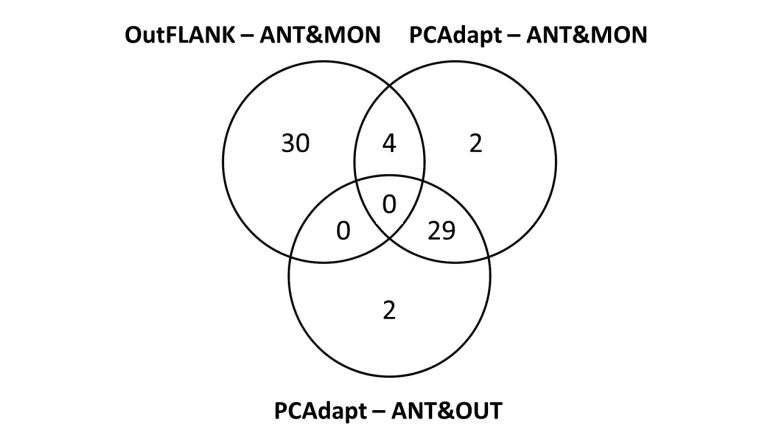
Résumé
L’introduction d’une espèce en milieu isolé est souvent associée à une perte de
la variation génétique, c’est l’effet fondateur. Les pressions de sélection peuvent
également différer du milieu d’origine, accélérant la divergence de la population
introduite. En 1896, environ 220 cerfs de Virginie (Odocoileus virginianus) provenant de
la rive sud du fleuve Saint-Laurent ont été introduits sur l’île d’Anticosti (Québec,
Canada) pour développer la chasse sportive. Cette population a rapidement atteint de
fortes densités (>20 cerfs/km2) et présente une taille corporelle inférieure à celle de la
population source. Ce projet avait comme objectifs d’évaluer le rôle de l’effet fondateur
et de la sélection sur la diversité et la structure génétique de la population de cerfs de
l’île d’Anticosti. Pour ce faire, la variabilité génétique de cette population a été comparée
à la population souche avec des techniques de séquençage à haut débit (GBS). Les
résultats montrent une faible différenciation génétique entre la population source et celle
de l’île d’Anticosti par rapport à l’ampleur de la différenciation génétique entre les rives
nord et sud du Saint-Laurent. L’absence de structuration à l’intérieur de l’île d’Anticosti
signifie que la dérive génétique n’a pas eu d’effet significatif lors de la colonisation de
l’ensemble de l’île. Ainsi, la grande diversité génétique de la population de cerfs
d’Anticosti et sa faible différenciation par rapport à celle du continent suggèrent un faible
effet fondateur. Le grand nombre d’individus introduits et la rapide croissance
démographique de la population ont sans aucun doute contribué à diminuer la dérive
génétique et expliquent le maintien d’une grande diversité génétique. De plus, des loci
potentiellement sous sélection ont été associés aux traits phénotypiques précédemment
identifiés comme différents entre les deux populations. Ces loci appuient une divergence
adaptative de la population insulaire en réponse aux conditions particulières de l’île
d’Anticosti. Les résultats de cette étude appuient la différenciation génétique de la
population de cerfs de Virginie de l’île d’Anticosti.
iiiAbstract
The introduction of a species is often associated with a loss of genetic variability,
referred as the founder effect. Different selection pressures in the colonized environment
may also accelerate the genetic divergence of two populations. In 1896, ca. 220 white-
tailed deer (Odocoileus virginianus) captured on the southern shore of the St. Lawrence
River were introduced on Anticosti Island (Québec, Canada) to develop sport hunting.
The absence of predator allowed the population to quickly reached high densities (>20
deer/km2) that affected forest composition because of browsing intensity. Deer body size
decreased by half compared to the source population because of their poor-quality diet.
Our objectives were to characterize the impacts of the founder effect and selection on
the diversity and genetic structure of the deer population on Anticosti Island. We
compared the genetic variability of this population to one of the strain populations
(Montmagny-L’Islet region) with high throughput sequencing techniques (GBS). The
results show a small, albeit significant genetic differentiation, between the source
population and Anticosti Island relative to the extent of genetic differentiation between
the north and south shores of the St. Lawrence. We found a lack of structuration within
the island meaning that the genetic drift did not have a significant magnitude during the
colonization of the whole island. The high genetic diversity of the Anticosti deer
population and its low differentiation from the mainland suggests a weak founder effect.
The large number of introduced individuals and the rapid population growth of the
population have undoubtedly helped to reduce genetic drift and explain the maintenance
of a great genetic diversity. Loci under putative selection were assigned to the
differentiated phenotypic traits supporting an adaptive divergence of the island
population in response to particular conditions. This study supports the genetic
differentiation of the white-tailed deer population of Anticosti Island.
ivTable des matières
Résumé.......................................................................................................................... iii
Abstract .......................................................................................................................... iv
Liste des tableaux ......................................................................................................... vii
Liste des figures ............................................................................................................viii
Remerciements .............................................................................................................. ix
Avant-propos ................................................................................................................. xii
Introduction ..................................................................................................................... 1
Les mécanismes évolutifs ............................................................................................ 1
Adaptation locale : sélection et plasticité phénotypique ............................................ 2
L’effet fondateur ....................................................................................................... 3
Les systèmes insulaires ............................................................................................... 5
Le cerf de Virginie ........................................................................................................ 7
Biologie .................................................................................................................... 7
Structure génétique des populations ........................................................................ 8
Le cas à l’étude : les cerfs introduits sur l’île d’Anticosti ............................................. 10
Objectifs et hypothèses ................................................................................................. 11
Approche méthodologique............................................................................................. 12
Chapitre 1 Weak founder effect and evidence for adaptive divergence in a recently
introduced insular population of white-tailed deer (Odocoileus virginianus) ................... 14
Résumé ..................................................................................................................... 15
Abstract ..................................................................................................................... 16
Introduction ................................................................................................................ 17
Material and Methods ................................................................................................ 19
Study area.............................................................................................................. 19
Sampling and phenotypic measurements ............................................................... 21
Genomic analyses .................................................................................................. 21
Results ...................................................................................................................... 27
SNP calling ............................................................................................................ 27
Clustering analysis ................................................................................................. 27
Genetic structure and population assignment ......................................................... 28
Outlier detection with genome scan approaches .................................................... 30
vPhenotype-genotype associations .......................................................................... 31
Genetic basis of phenotypic differentiation ............................................................. 32
Discussion ................................................................................................................. 34
Genetic differentiation and structure of Anticosti Island deer .................................. 34
Weak founder effect ............................................................................................... 36
Adaptation on the Island......................................................................................... 37
Limitations .............................................................................................................. 40
Management Implications ...................................................................................... 41
Acknowledgements.................................................................................................... 42
Conclusion générale...................................................................................................... 43
Différenciation génétique ........................................................................................... 43
Absence de structuration génétique ........................................................................... 44
Adaptation locale de la population ............................................................................. 45
Taille de population efficace et sélection sexuelle ...................................................... 46
Implications pour la gestion ....................................................................................... 48
Forces et limites de l’étude ........................................................................................ 50
Perspective de recherche .......................................................................................... 51
Références.................................................................................................................... 53
Annexe .......................................................................................................................... 66
viListe des tableaux
Table 1. Below main diagonal: Fixation index (FST) obtained with 8,518 SNPs between
populations of white-tailed deer from western Anticosti Island (W-AN), central Anticosti
Island (C-AN), eastern Anticosti Island (E-AN), Montmagny (MON) and Outaouais
(OUT); above main diagonal: 95% confidence intervals. ........................................................... 29
Table 2. Descriptive genetic statistics of white-tailed deer populations obtained with
8,518 SNPs: observed heterozygosity (Ho), expected heterozygosity (He), inbreeding
coefficients (Gis and F), nucleotidic richness (π), number of polymorphic loci (N),
effective population size (Ne), census population size (Nc) for Anticosti Island (ANT),
Montmagny (MON) and Outaouais (OUT) populations. ............................................................. 30
viiListe des figures
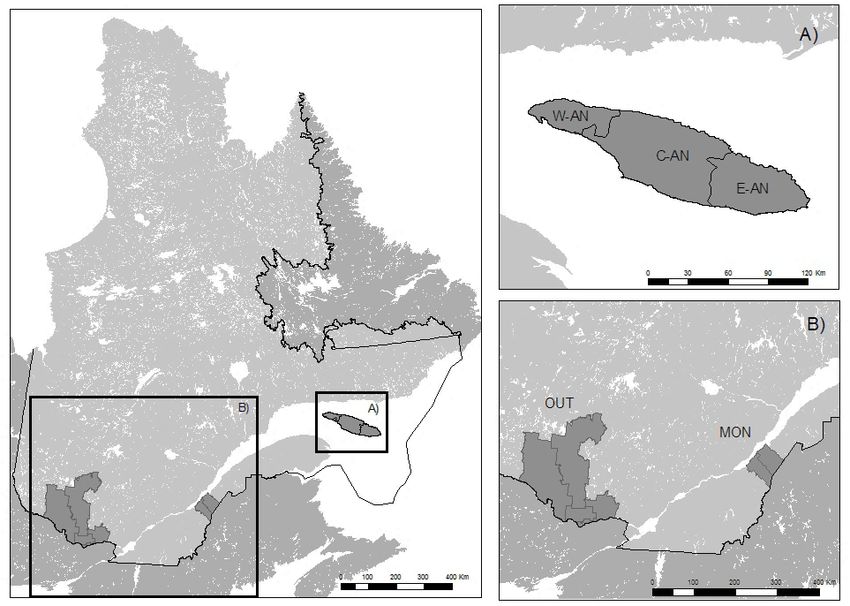
Figure 1. The three study areas (Québec, Canada). A) Anticosti Island divided into three
regions: western (W-AN), central (C-AN), and eastern (E-AN). B) Montmagny-L’Islet
(MON) and Outaouais (OUT).............................................................................................................. 20
Figure 2. Discriminant analysis of principal components (DAPC) of the genetic variation
of white-tailed deer without prior information on their group provenance: Western
Anticosti (W-AN), Central Anticosti (C-AN), Eastern Anticosti (E-AN), Montmagny (MON)
and Outaouais (OUT). ........................................................................................................................... 28
Figure 3. Venn diagrams of outliers detected by OutFLANK and pcadapt between white-
tailed deer from Anticosti Island and Montmagny (ANT&MON), and between Anticosti
Island and Outaouais (ANT&OUT). .................................................................................................. 30
Figure 4. Venn diagrams of loci, detected by LFMM, associated with rump fat thickness,
peroneus muscle mass, hind foot length and dressed body mass of male (A) and female
(B) white-tailed deer from Anticosti Island. ..................................................................................... 31
Figure 5. Correlations between the polygenic score and the corresponding phenotypic
trait (rump fat thickness, peroneus muscle mass, hind foot length, dressed body mass,
and antler spread) for male (Left) and female (Right) white-tailed deer from Anticosti
Island. All correlations were significant (p ˂ 0.001). .................................................................... 33
Figure 6. (A) Venn diagram of loci found in common by pcadapt analyses between
Anticosti Island and both mainland sites (Montmagny and Outaouais), compared to all
loci found by random forest and LFMM analyses. (B) Venn diagram of loci found by all
genome scan approaches (OutFLANK and pcadapt) among populations compared to all
loci found by random forest and LFMM analyses. ....................................................................... 34
viiiRemerciements
Lorsque j’ai entendu parler du projet sur les cerfs de l’île d’Anticosti, j’ai su qu’il était fait
pour moi. Depuis toujours les îles m’ont fasciné par leur caractère unique. Les aventures
d’Alfred Russel Wallace et de Charles Darwin m’ont particulièrement émerveillé durant
mes études. Ce fut pour moi un réel privilège de travailler sur ce projet longuement
attendu.
Je tiens tout d’abord à remercier mon codirecteur, Louis Bernatchez, d’avoir eu
confiance en moi et de m’avoir donné la chance de faire partie de son laboratoire. J’ai
eu aussi le plaisir de te connaître en Équateur et sur les îles Galapagos! Mythique! Nous
avons eu de grandes discussions, dont je garde un merveilleux souvenir. Je te remercie
pour toute l’aide que tu m’as apportée. Tu t’es également assuré que mon projet de
maîtrise me convenait en m’offrant la possibilité de travailler sur les mammifères. Je
remercie mon directeur, Steeve Côté, qui m’a accepté dans son laboratoire alors que je
n’étais qu’un inconnu. Je n’ai pas eu la chance d’assister à l’un de tes cours durant mon
baccalauréat, mais heureusement j’aurai pu bénéficier de ton enseignement à titre de
directeur. J’ai grandement apprécié ta rigueur, tes conseils et tes nombreuses histoires.
Merci pour tout. Je remercie également Julie Turgeon et Jean-Pierre Tremblay d’avoir
accepté de réviser mon mémoire.
Ce projet n’aurait pas eu la même envergure sans les professionnels de recherche que
j’ai côtoyés durant ces années. Pour le labo Côté-Tremblay, je tiens à remercier Sonia
de Bellefeuille, qui m’a grandement aidé avec les bases de données, Caroline Hins pour
tes précieux conseils et Julien Hénault-Richard pour la relecture de mes travaux. Pour le
labo Bernatchez, un immense merci à Cécilia Hernandez, Guillaume Côté et Damien
Boivin-Delisle de m’avoir aidé à (finalement) extraire de l’ADN de bonne qualité, et bien
sûr merci à Éric Normandeau pour son soutien en bioinformatique et nos conversations
en matière d’art martial.
Je tiens à souligner le soutien financier fortement apprécié de la Chaire de recherche
industrielle CRSNG en aménagement intégré des ressources de l’île d’Anticosti et de
ses partenaires. Merci également à la Fédération québécoise des chasseurs et
pêcheurs (FédéCP) et à la fondation Héritage faune de m’avoir décerné la bourse
Aurèle-Blais en aménagement de la faune.
Je remercie les gens de l’île d’Anticosti, sans qui mon terrain n’aurait pas été aussi
mémorable. Alex/Gaétan Laprise et Danièle Morin, un gros merci pour votre aide et pour
le séjour de chasse! Aimée Benoist-Chénier et Stefan Tremblay, merci beaucoup pour
votre accueil et votre soutien dans mes péripéties de terrain! Je remercie tous ceux et
celles qui ont, de près ou de loin, participé à la récolte des échantillons pour l’ensemble
du projet, notamment Alexandre Lottinville, technicien pour la chaire de recherche à
l’automne 2015; Michaël Bonin et ses assistant(e)s pour les échantillons de Montmagny;
Vicky Albert et l’équipe du MFFP du laboratoire d’expertise biolégale ainsi qu’André
ixDumont et la direction régionale de l’Outaouais du bureau de Gatineau pour les
échantillons d’Outaouais.
Une maîtrise, ce n’est pas uniquement du boulot, c’est aussi des rencontres
inoubliables! J’ai eu la chance de côtoyer non pas un laboratoire, mais deux! Et pas
parmi les plus petits.
Au labo Côté-Tremblay : Mael, Mathieu, Julien, Andréa, Karina, Brett, Amé, Pat, Ed,
Pascale, Max, Dalie, Clara, Sab, Émilie, Mic, Barb, Antoine, Myriam, Milène, Nico, Béa,
Flo, Laurent, Fred, Alexis (pour les pommes de 15 h), Linda, Delphine, Ève et Sophiane.
Merci gang pour votre aide et pour ses rires partagés, ces bières du jeudi, ces soirées
inoubliables, et cheers aux nombreux fans de Pérusse (#Fruitdemer). Vous êtes des
personnes extraordinaires et je vous souhaite bonne chance dans vos projets. Un merci
spécial à Marianne, ma partenaire généticienne en ce monde d’écologistes! Que de
beaux souvenirs en C.B.! Merci pour ton écoute et ta sensibilité. Je te souhaite le
meilleur, mon amie.
Au labo Bernatchez : J.-S., Anne-Laure, Martin, Jérémy, Ben, Hugo, Claire, Anaïs,
Justine, Noé, les nombreuses Anne, Damien, Clément, Laura, Yann, Quentin, Bérénice,
Lucie, Maëlle, Alysse, Simon, Charles, Émilie, Gratien, Olivier, Amanda, Madoka, Henrik
et Caro. Merci pour ces discussions enrichissantes, pour votre aide indispensable et
pour ces soirées qui n’en finissent plus (#workhardplayharder). Réaliser ma maîtrise
avec vous fut un privilège et une fierté. Je vous souhaite du bonheur et le succès que
vous méritez.
Merci également aux collègues du Vachon : Yannick, Mathilde, Guillaume, Fred, Dom,
Isa, Marc-André, Marianne et Audrey-jade ainsi qu’à mes collègues de l’IBIS : F-E (aka
Francisco de Padre!), F.-O., Laurence, Jeff, Bachar, Pier-Luc vous côtoyer au quotidien
fut un grand plaisir.
C’est également important d’être bien entouré à l’extérieur de l’université. Je tiens donc
à remercier mes ami(e)s, dont ma gang des Zozios pour votre soutien et nos soirées
mémorables. M. Brousseau! même si tu as été nommé dans la précédente section, tu
sais bien que tu pourrais avoir une section juste pour toi, bro! Apprendre à te connaître,
surtout à titre de coloc, fut un privilège. Merci Max d’être toujours là pour moi et de me
remonter le moral quand j’en ai besoin, t’es plus qu’un ami et j’espère que l’on gardera
contact malgré le temps. Ma maîtrise n’aurait certainement pas été la même sans toi.
Anne, toi aussi tu mérites d’avoir une section à part. Merci chère amie pour tes conseils
et ton énergie contagieuse. Je te souhaite bonne chance dans tes projets. Mélina, ma
grande amie, merci pour ton soutien au fil de toutes ses années. Bien sûr, Gab, la
meilleure annonce Kijiji! Thanks buddy de me changer les idées avec nos
journées/soirées jeux de société. Je remercie mon club de Tai Jitsu, où j’ai découvert
une seconde passion, et un merci spécial à Marie-Eve Castonguay, qui a accepté de
réviser mon mémoire.
À ma famille, qui a toujours cru en moi et m’a encouragé vers les études de cycles
supérieurs, je vous aime. Audrée, merci pour ton soutien et ta compréhension, mais
surtout merci de faire partie de ma vie. Un merci spécial à mes grands-parents, qui
xm’ont transmis leur passion pour la nature, me dirigeant de ce fait vers la biologie. Mom,
merci d’être toujours là pour moi.
Pour tous les chasseurs et chasseuses qui ont cru en l’existence du cerf Menier, voici
une histoire qui saura, je l’espère, alimenter vos discussions!
Bonne lecture.
‘’Nothing in biology makes sense except in the light of evolution’’
- Theodosius Dobzhansky
‘’Why do we fall? So that we can learn to pick ourselves up’’
- Thomas Wayne
xiAvant-propos
Ce projet de maîtrise porte sur la différenciation génétique du cerf de Virginie
introduit sur l’île d’Anticosti (Québec, Canada), une étude réalisée dans le cadre de la
Chaire de recherche industrielle CRSNG en aménagement intégré des ressources de
l’île d’Anticosti. Le mémoire débute par une introduction générale sur les mécanismes
évolutifs qui agissent sur la divergence des populations, particulièrement en situation
insulaire. Le cœur de ce mémoire consiste en un article scientifique (manuscrit) rédigé
en anglais et qui a été soumis au journal scientifique Molecular Ecology le 22 juin 2018.
Le mémoire se termine par une conclusion générale qui résume la contribution de mon
étude au savoir scientifique et où j’énonce quelques idées de recherche soulevées par
mes résultats.
En tant que premier auteur, mes responsabilités consistaient à sélectionner les
échantillons parmi la collection de la chaire de recherche, à participer à la collecte de
données sur le terrain à l’automne 2015, à réaliser les manipulations de laboratoire, à
effectuer les analyses et l’interprétation des résultats ainsi qu’à rédiger l’article.
Les coauteurs sont Anne-Laure Ferchaud, Martin Laporte, Jérémy Le Luyer et
Theodore B. Davis, mon directeur, Steeve D. Côté, et mon codirecteur, Louis
Bernatchez.
Anne-Laure Ferchaud, Martin Laporte et Jérémy Le Luyer, tous postdoctorants,
ont contribué aux analyses et à l’interprétation des résultats. Ils ont également participé
à la révision de l’article.
Theodore B. Davis nous a fourni le génome du cerf de Virginie avant sa
publication et a contribué à l’amélioration du manuscrit.
L’interprétation des résultats et la rédaction du manuscrit ont été supervisées par
Louis Bernatchez et Steeve D. Côté. Ce dernier a proposé et financé l’étude. Louis
Bernatchez a également contribué à une partie du financement.
xiiIntroduction
Les mécanismes évolutifs
La diversité biologique actuelle découle de l’évolution durant des milliers
d’années des premiers microorganismes et, plus tard, des organismes multicellulaires
complexes. Cette histoire a été forgée par quatre forces évolutives : la mutation, la
migration, la dérive génétique et la sélection (Allendorf & Luikart 2007). La mutation
induit de la diversité génétique sur une échelle de temps beaucoup plus longue que les
autres forces. En effet, les taux de mutation sont en général assez faibles, soit de l’ordre
de 10-4 allant jusqu’à 10-9 mutations/locus/génération selon les régions du génome
(Brumfield et al. 2003). À l’opposé, le déplacement d’individus entre populations, la
migration (dispersion au sens écologique), induit un flux génétique entre ces populations
et provoque une homogénéisation des fréquences alléliques entre elles (Lowe &
Allendorf 2010). Inversement, l’isolation d’une population favorise sa divergence. De son
côté, la dérive augmente la différenciation entre les groupes de manière stochastique
provenant de la pige aléatoire des gamètes lors des événements de reproduction
(Frankham 1995, Höglund 2009). Les allèles retenus ne représentent pas parfaitement
la fréquence allélique de la génération précédente, ce qui résulte ultimement en une
perte de diversité allélique (Charlesworth 2003). Ce phénomène est amplifié dans les
populations où la taille efficace est restreinte. Ces trois forces évolutives affectent
l’ensemble du génome de manière aléatoire et sont donc décrites comme « neutres »,
contrairement à la sélection qui agit sur certains loci en réponse aux pressions
environnementales. À noter qu’il existe différents types de sélection : directionnelle,
balancée, disruptive et sexuelle (voir la section Adaptation locale : sélection et plasticité
phénotypique).
Ces quatre forces évolutives figurent parmi les responsables des différences
génétiques, morphologiques et comportementales entre les populations d’une même
espèce. Parmi les nombreuses branches de la biologie, l’une tente de comprendre
l’influence de ces forces évolutives sur le vivant : la biologie évolutive. Plus active que
jamais, elle a bénéficié de l’essor des nouvelles technologies pour apporter plus de
précisions aux questions posées depuis plusieurs années.
1Adaptation locale : sélection et plasticité phénotypique
À l’inverse des trois forces neutres, la sélection agit directement sur le phénotype
des individus, qui est déterminé par l’interaction de certains loci avec l’environnement.
Ainsi, les allèles offrant un avantage sur la reproduction ou la survie d’un individu sont
favorisés et sont, en théorie, présents en plus grande proportion dans une population.
La fixation d’allèles bénéfiques et la disparition d’allèles délétères contribuent à la
différenciation entre deux groupes soumis à différentes pressions de sélection (Kawecki
& Ebert 2004, Allendorf & Luikart 2007, Nosil et al. 2009).
Une espèce fait face à différentes conditions environnementales dans son aire
de répartition, ce qui occasionne une hétérogénéité de pressions de sélection dans
l’espace. Les individus de différentes populations développent alors des adaptations
locales, soit des changements génétiques associés à leur environnement en réponse à
la sélection divergente (Le Corre & Kremer 2012, Savolainen et al. 2013). L’apparition
d’adaptations locales est facilitée en absence de flux de gènes et de dérive génétique,
car ces processus peuvent compromettre les génotypes favorisés par la sélection
(Kawecki & Ebert 2004, Blanquart et al. 2013). La définition classique soutient que la
survie des individus localement adaptés est inférieure s’ils se retrouvent dans un
nouveau milieu (Kawecki & Ebert 2004). D’autres types de sélection sont également
possibles. La sélection balancée, ou stabilisatrice, entraîne une augmentation de la
diversité génétique par l’action de différents mécanismes, dont l’avantage de
l’hétérozygote (Hansson & Westerberg 2002, Charlesworth 2006, Llaurens et al. 2017).
La sélection sexuelle dépend de la compétition survenant lors du choix des partenaires
et peut agir à l’encontre de la sélection naturelle. Elle se produit lorsqu’un individu
augmente son succès reproducteur grâce à un caractère sexuel secondaire, même si ce
caractère peur compromettre sa survie (Clutton-Brock 2017). Les changements
phénotypiques issus d’un changement génétique ne sont pas uniquement causés par la
sélection : la dérive peut également être responsable d’une différenciation phénotypique
(Jordan & Snell 2008, Kolbe et al. 2012, Spurgin et al. 2014). Les forces évolutives
neutres ont un impact aussi significatif que la sélection sur les populations et peuvent
ainsi être les précurseurs à la différenciation et ultimement à la spéciation.
2L’expression d’un phénotype différent n’implique pas nécessairement un
changement génétique, car il est également déterminé par l’environnement. La plasticité
phénotypique est définie comme l’expression de traits phénotypiques différents par des
organismes possédant un même génotype, mais situés dans des environnements
différents (Pigliucci et al. 2006). Par exemple, une population de lézard, l’anole brun
(Anolis sagrei), provenant de la Floride, fut introduite sur deux îles aux conditions
abiotiques fort différentes, et les individus de ces deux nouvelles populations ont
rapidement divergé en taille (Campbell & Echternacht 2003). La plasticité a un rôle à
jouer dans l’établissement d’une population dans un nouveau milieu, car elle détermine
la capacité d’acclimatation d’un individu (Price et al. 2003, Uller & Leimu 2011). Les
espèces qui font face à des conditions climatiques très variées, par exemple les
espèces migratrices ou nordiques, sont aussi avantagées par une capacité à exprimer
une plasticité de leurs traits.
On parlera d’adaptation lorsque le changement phénotypique se conserve à
l’échelle de la population. La plasticité phénotypique peut être un trait sélectionné, car
elle peut offrir un avantage à l’individu (Schmid & Guillaume 2017). L’adaptation à un
environnement peut toutefois être le résultat de la plasticité phénotypique combinée à la
différenciation génétique des traits adaptatifs (Sultan & Spencer 2002, Volis et al. 2015,
Schmid & Guillaume 2017). Par exemple, les printemps hâtifs causés par les
changements climatiques ont entraîné en 10 ans une date de mise bas précoce de 18
jours chez une population d’écureuil roux (Tamiasciurus hudsonicus) au Yukon (Réale et
al. 2003). Ce trait héritable est également connu pour varier selon des conditions
environnementales, dont l’abondance de nourriture (Réale et al. 2003).
L’effet fondateur
Une introduction d’espèce naturelle ou par l’intermédiaire de l’homme peut
occasionner une divergence génétique marquée par rapport à la population source
(Clegg et al. 2002, Kolbe et al. 2012). Ce phénomène, appelé effet fondateur, est un cas
particulier de la dérive génétique (Mayr 1954). C’est l’un des processus les plus forts,
avec les goulots d’étranglement (réduction drastique de la population), qui peut
rapidement engendrer une divergence génétique entre deux populations à la suite d’une
perte importante de la variation génétique et d’un brusque changement des fréquences
3alléliques (Nei et al. 1975, Allendorf 1986, Clegg et al. 2002). L’intensité de l’effet
fondateur varie selon le nombre d’événements d’introduction, le nombre d’individus
introduits, la diversité génétique de la population source ainsi que le temps nécessaire
pour que la population augmente en nombre (Clegg et al. 2002, Blanchong et al. 2013,
Murphy et al. 2015). Ainsi les espèces avec un taux de croissance intrinsèque plus faible
(ex. les grands mammifères) devraient perdre davantage de variation génétique que les
espèces avec des taux de croissance plus élevés (ex. les insectes et les petits
mammifères), car ces dernières nécessitent généralement moins de temps pour
augmenter en nombre (Allendorf & Luikart 2007). À la suite d’un effet fondateur ou d’un
goulot d’étranglement, la perte de variation génétique sera plus accentuée pour la
richesse allélique que l’hétérozygotie, car elle est plus sensible à la perte d’allèles rares
(Nei et al. 1975, Dlugosch & Parker 2008). La richesse allélique, soit le nombre d’allèles
à un locus, est plus importante pour le potentiel adaptatif à long terme d’une population,
car elle témoigne de la diversité sur laquelle la sélection peut opérer (Allendorf 1986,
Caballero & Garcia-Dorado 2013, Greenbaum et al. 2014).
La taille de la population, ou plus exactement le nombre réel d’individus qui
contribuent à la génération suivante, est un paramètre important pour la dérive
génétique. Cela fut conceptualisé par la taille de population efficace (Ne) qui est définie
comme l’effectif d’une population idéale, c’est-à-dire qui est soumise aux même taux de
dérive génétique et de consanguinité que la population étudiée (Wright 1931). Ainsi, il
s’agit d’un paramètre difficile à estimer, car plusieurs facteurs peuvent l’influencer, le
ratio des sexes, le succès reproducteur des individus et la démographie de la population
par exemple (Frankham 1995, Kalinowski & Waples 2002, Hedrick 2005, Palstra &
Ruzzante 2008). Les populations de grande taille efficace sont moins propices à la
dérive génétique, ce qui accentue le rôle de la sélection sur la fréquence allélique (Hartl
& Clark 1997, Blanquart et al. 2013).
Dans les petites populations, le choix du partenaire est réduit et le risque de
reproduction entre individus apparentés augmente. Ainsi, les taux de consanguinité sont
appelés à augmenter, ce qui diminue la variabilité génétique, indépendamment de la
dérive. Une diversité génétique faible est généralement associée à une moins bonne
valeur adaptative des individus (Reed & Frankham 2003, Wood et al. 2016). Cette
baisse de valeur sélective nommée dépression de consanguinité s’explique par
4l’augmentation de l’homozygotie au détriment de l’hétérozygotie sur l’ensemble du
génome (Keller & Waller 2002, Allendorf & Luikart 2007). C’est pourquoi des allèles
récessifs délétères, maintenus normalement à une fréquence faible par la sélection, sont
statistiquement plus sujets à se trouver en état d’homozygotie dans les populations
consanguines (Keller & Waller 2002, Allendorf & Luikart 2007). Ces allèles récessifs
seront alors exprimés et diminuent la valeur sélective de l’individu homozygote compte
tenu de leurs effets délétères. Par exemple, une population de cerfs rouges (Cervus
elaphus) isolée au nord de l’Allemagne présente aujourd’hui une forte tendance à la
brachygnathie, soit une réduction de la mâchoire, à la suite d’une dépression de
consanguinité (Zachos et al. 2007).
Un cas spécifique de la consanguinité peut survenir lors d’une introduction ou
d’une colonisation. Comme le nombre d’individus fondateurs est limité, un allèle délétère
porté par un individu aura statistiquement plus de possibilités de se trouver en forte
fréquence malgré sa valeur contre-sélective (Keller & Waller 2002, Casellas et al. 2009).
Cela augmente la probabilité de dépression consanguine dans la nouvelle population et
peut menacer la viabilité de celle-ci. C’est pourquoi les populations de taille plus
restreinte sont généralement sujettes à une attention et à des efforts de conservation
plus intensifs. Le maintien de certaines populations nécessite parfois un sauvetage
génétique qui consiste à ajouter des individus d’une autre population pour augmenter la
variation génétique (Johnson et al. 2010, Whiteley et al. 2015).
Les systèmes insulaires
Les îles ont toujours fasciné les biologistes, particulièrement pour la présence
d’organismes ayant une morphologie différente de leurs homologues continentaux. La
tendance morphologique généralement observée sur les îles est une augmentation de la
taille des petites espèces introduites (ex. lézards et rongeurs) et une réduction de la
taille des grandes espèces (ex. éléphants et cervidés) (Meiri et al. 2008). VanValen
(1973) a nommé ce phénomène « règle insulaire » et plusieurs méta-analyses ont tenté
d’en déterminer les causes par la phylogénie, les interactions biotiques (compétition et
prédation) ou les caractéristiques des îles (Lomolino 2005, Raia & Meiri 2006, Meiri et
al. 2008, McClain et al. 2013). Tout semble indiquer que ce changement de taille est
influencé par la combinaison de ces différents facteurs, dont l’importance varie
5largement selon le contexte (Durst & Roth 2012, Lomolino et al. 2012). Bien que certains
clades soient sur-représentées dans ces analyses en raison du grand nombre
d’introductions, il a été établi que les artiodactyles, les carnivores et les rongeurs sont
plus prédisposés évolutivement à répondre à cette règle pour des raisons encore
inconnues (Meiri et al. 2008, Raia & Meiri 2011). Par rapport aux carnivores, les
herbivores ont tendance à diminuer davantage en taille en absence de compétiteurs
hétérospécifiques et de prédateurs (Raia & Meiri 2006).
Les îles sont reconnues pour présenter des conditions environnementales
différentes des continents, en raison entre autres de la proximité de l’eau, qui exerce
des pressions de sélection divergentes sur les espèces (Weigelt et al. 2013). Les
espèces végétales sont les premières à répondre à ces changements
environnementaux, ce qui modifie la composition végétale et offre par conséquent des
ressources différentes aux consommateurs primaires (Agashe et al. 2011, Weigelt et al.
2013). Ainsi, les îles moins productives sont généralement occupées par des espèces
de petite taille (McClain et al. 2013). La compétition pour les ressources y est souvent
supérieure en raison de la superficie limitée, ce qui favorise la spécialisation pour des
niches trophiques peu utilisées (Warren et al. 2015). La faible présence de prédateurs
généralement observée dans les systèmes insulaires induit une faible pression de
sélection sur les espèces, ce qui leur permet d’adopter des comportements et
phénotypes qui seraient autrement contre-sélectionnés en présence d’une plus grande
densité et diversité de prédateurs (Runemark et al. 2014).
En général, les espèces insulaires montrent une variation génétique faible
comparativement aux espèces continentales qui leur sont apparentées, en raison de
leur isolement et de la plus forte fréquence de goulots d’étranglement (Frankham 1997).
La méta-analyse de Uller & Leimu (2011) a révélé que cette baisse de diversité est
particulièrement importante chez les insectes et les mammifères introduits dans un
nouvel environnement. On retrouve parfois quelques exceptions où la diversité
génétique de certaines populations introduites ne diffère pas significativement de la
population source, même en milieu isolé (voir Tableau S7, Annexe).
Ces laboratoires naturels que sont les îles suscitent l’intérêt pour la recherche en
biologie évolutive de par leur relative simplicité. Selon la biologie de l’espèce, la distance
6séparant les îles de la côte peut empêcher tout flux de gènes avec les populations
continentales laissant ainsi la dérive, la sélection et selon la période temporelle, la
mutation agir sur les populations (Pemberton et al. 1996, Blanquart et al. 2013).
Plusieurs introductions d’espèces sur des îles ont été réalisées par l’homme, ce qui
permet l’accès à des informations comme la date et le nombre d’individus introduits qui,
lorsque mises en lien avec la population source ou l’ancêtre continental, offrent
d’incroyables possibilités d’approfondir nos connaissances en biologie évolutive (Uller &
Leimu 2011).
Le cerf de Virginie
Biologie
Le cerf de Virginie (Odocoileus virginianus, Zimmermann 1780) est l’un des
cervidés les plus répandus d’Amérique Nord, son aire de répartition s’étendant du
Québec au Brésil (Seabury et al. 2011). Cette espèce qui a su s’adapter à différents
milieux possède une grande valeur économique en raison de la chasse sportive, ce qui
explique les nombreuses recherches à son sujet. En 2000, on estimait les revenus de la
chasse sportive à 78,2 M$ au Québec, dont 12 M$ en provenance d’Anticosti (Huot &
Lebel 2012). D’ailleurs, la chasse a provoqué une forte exploitation des cerfs dans
plusieurs régions de l’Amérique du Nord au début du 20e siècle (DeYoung et al. 2003,
Monteith et al. 2013). La chasse sportive influence la structure d’âge et la dynamique
des populations (Festa-Bianchet 2003, Milner et al. 2007), ce qui en fait un outil de
gestion efficace, lorsque bien contrôlé. La présence de chasseurs diminue le niveau
d’activité diurne, dont la quête de nourriture, ce qui affecte négativement la masse des
cerfs (Little et al. 2016, Strickland et al. 2017). Dans un contexte de surabondance, où la
compétition pour les ressources est déjà forte, une intense pression de chasse pourrait
affecter grandement la masse d’une population de cerfs et ultimement la survie des
individus. En ciblant des phénotypes particuliers, la chasse aux trophées peut
occasionner des changements dans les fréquences alléliques de la population si la
pression de chasse est très intense (Coltman et al. 2003a, Monteith et al. 2013,
Heffelfinger 2018).
7Le temps de génération, soit le temps nécessaire au remplacement d’une
cohorte, est d’approximativement quatre ans chez le cerf de Virginie (Kekkonen &
Brommer 2015). Il s’agit d’un mammifère polygame : les mâles peuvent se reproduire
avec plusieurs femelles pendant le rut, qui a lieu à la fin de l’automne. Durant cette
période, les mâles accordent moins de temps à l’alimentation et occupent la majorité de
leur temps à la recherche active de partenaires et à leur défense face à d’autres mâles
(Holzenbein & Schwede 1989, Mainguy & Côté 2008). La condition corporelle des mâles
après le rut en est affectée, et ce, au moment d’entrer dans la difficile période hivernale.
Durant cette rude saison, les cerfs continentaux se rassemblent généralement en des
lieux denses en conifères appelés « ravages » pour retourner à leur aire estivale au
printemps. C’est au printemps que les femelles mettent bas à un ou deux petits. La
condition corporelle, notamment le pourcentage de gras, des femelles est positivement
reliée à la taille de portée puisque le gras est essentiel à la production d’énergie requise
pour le développement des fœtus et à la thermorégulation de la mère durant l’hiver
(Cothran et al. 1987, Ayotte et al. non publié). De plus, les femelles avec une
hétérozygotie plus élevée produisent davantage de jumeaux (Cothran et al. 1987,
Leberg et al. 1992).
La plasticité du cerf de Virginie est l’une des explications de sa grande aire de
répartition. La qualité des sols, et donc du fourrage, et la date de naissance expliquent
davantage la morphologie des cerfs que leur génétique (Gray et al. 2002, Strickland &
Demarais 2008, Jones et al. 2010). L’influence de ces composantes environnementales
par rapport aux composantes génétiques a été mise en évidence dans des expériences
de jardin commun avec le cerf de Virginie (Michel et al. 2016). De manière générale,
l’effet de l’environnement sur la taille corporelle est largement démontré, bien que ce
trait soit héritable (Ozgul et al. 2009, Morrissey et al. 2012).
Structure génétique des populations
Plusieurs études sur la génétique du cerf de Virginie, notamment aux États-Unis,
ont été réalisées dans le passé. La majorité témoigne d’une diversité génétique élevée
(Smith et al. 2001), et ce, même en milieu perturbé par les activités anthropiques
(diversité des ADN microsatellites : H : 0.67–0.74, DeYoung et al. 2003, Blanchong et al.
2013). Le wapiti nord-américain (Cervus elaphus), qui fut lui aussi fortement exploité,
8affiche une diversité génétique nettement inférieure à celle du cerf (0.26-0.54, Polziehn
et al. 2000). Il en va de même pour l’orignal (Alces alces) (0.22-0.41, Broders et al.
1999). La capacité des cerfs à atteindre de fortes densités rapidement (DeYoung 2011)
et leur structure sociale peuvent expliquer le maintien de leur grande diversité génétique
(Blanchong et al. 2013). Les femelles ont généralement un comportement philopatrique
et tendent à demeurer à l’intérieur d’un rayon de 50 km de leur lieu de naissance une
fois l’âge adulte atteint (Purdue et al. 2000). Un comportement similaire explique en
grande partie la structuration génétique chez le cerf rouge (Nussey et al. 2005) et aussi
chez d’autres ongulés (Coltman et al. 2003b). À l’opposé, les mâles ont tendance à se
disperser, surtout en période de rut, sur des distances aussi grandes que 200 km
(Purdue et al. 2000). Ils maintiennent ainsi une variation génétique à l’intérieur des sous-
populations (Purdue et al. 2000). Les femelles vont effectuer leurs plus grands
mouvements presque exclusivement lors de la mise bas, cherchant à s’éloigner de leurs
congénères pour éviter la compétition (Lutz et al. 2015). Seules les grandes rivières
semblent constituer une barrière considérable pour l’espèce (Long et al. 2010, Locher et
al. 2015). Au Québec, le fleuve Saint-Laurent s’est révélé être la principale barrière aux
échanges entre les populations (Albert 2007). Ces déplacements entraînent une faible
structure génétique même à grande échelle (> 1000 km), comme observé entre des
sous-populations de cerfs de Virginie et de cerfs mulet (Odocoileus hemionus hemionus,
Rafinesque 1817) de l’Ouest canadien avec des FST globaux de 0.006 et 0.008
respectivement (Cullingham et al. 2011a, 2011b).
En tant que taxon privilégié pour la chasse sportive, les ongulés ont subi de
nombreuses translocations. Même en présence d’effet fondateur, les cerfs de Virginie
peuvent avoir une croissance démographique élevée lorsque les conditions
environnementales sont adéquates. Ces fortes croissances pourraient suffire à maintenir
une bonne diversité génétique malgré la perte d’allèles causée par un fort effet
fondateur. C’est ce qu’a illustré une étude récente portant sur la corrélation entre la
valeur sélective et l’hétérozygotie chez une population de cerfs de Virginie fondée par
quatre individus (Brommer et al. 2015). Ces quatre individus ont été introduits au sud de
la Finlande en 1934 et, de nos jours, les recensements témoignent de >50 000 individus
et une hétérozygotie moyenne de l’ADN microsatellite de 0.68 malgré une diminution de
la richesse allélique (Kekkonen et al. 2012, Brommer et al. 2015). Malgré leur isolement,
ces cerfs ne démontrent pas de différence morphologique par rapport à leur population
9source américaine (Kekkonen et al. 2016). Inversement, les cerfs des Keys (Odocoileus
virginianus clavium), appartenant à une sous-espèce du cerf de Virginie isolée sur
l’archipel des Keys au sud de la Floride, se sont fortement différenciés
morphologiquement et génétiquement des cerfs continentaux (FST = 0.202) à la suite de
leur isolement et de leur forte exploitation dans les années 1950 (Villanova et al. 2017).
En comparaison, les deux lignées évolutives d’Odocoileus hemionus, soit le cerf mulet
et le cerf à queue noire, ont une différenciation génétique plus modérée (FST = 0.123,
Powell et al. 2016).
Le cas à l’étude : les cerfs introduits sur l’île d’Anticosti
C’est pour la chasse sportive que le cerf de Virginie a été introduit dans la partie
ouest de l’île d’Anticosti (Québec, Canada) en 1896 et 1897 avec approximativement
220 individus (Newsome 1937). Ces derniers provenaient de la rive sud du fleuve Saint-
Laurent, principalement de la région de Montmagny-L’Islet (Martin-Zédé 1938,
McCormick 1982, Luc Jobin communication personnelle). À cette époque, l’île
d’Anticosti avait des ours noirs (Ursus americanus) comme seuls prédateurs (pour les
faons seulement), ce qui a néanmoins permis l’expansion rapide du cerf de Virginie
jusqu’à la disparition des ours en moins de 70 ans (Côté 2005). Dès les années 1930,
les effets négatifs sur la végétation causés par le broutement intensif des cerfs signalent
une grande population de cerfs (Marie-Victorin & Rolland-Germain 1969, Côté et al.
2008). Puis, vers la fin des années 1980, à l’aide de techniques d’inventaire modernes,
on a estimé leur nombre à 120 000 cerfs (Potvin et al. 1991) et, plus récemment en
2006, à >160 000 (Rochette & Gingras 2007). La grande pression de broutement
imposée à la végétation a même affecté la structure forestière de l’île. En effet, la
diminution des sapinières, ressources hivernales importantes pour le cerf (Lefort et al.
2007, Sauvé & Côté 2007), a entraîné l’augmentation des peuplements d’épinette
blanche, qui est une espèce de moins bonne qualité nutritive (Potvin et al. 2003).
La fluctuation de taille de population n’a pas été le seul changement observé sur
cette population. Des changements morphologiques ont en effet été mentionnés dans la
littérature, dont la petite taille des cerfs de l’île d’Anticosti (Lesage et al. (2001). Cette
même étude rapportait que les faons de l’île d’Anticosti avaient une plus grande
longueur des pattes, proportionnellement au corps, ainsi que des réserves de gras à
10Vous pouvez aussi lire

















































