Importance de l'imagerie spatiale à haute résolution pour la détection de l'effet de lisière en hiver - Mémoire Yann Baril-Chauvette Maîtrise en ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Importance de l'imagerie spatiale à haute résolution
pour la détection de l'effet de lisière en hiver
Mémoire
Yann Baril-Chauvette
Maîtrise en sciences forestières - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Yann Baril-Chauvette, 2022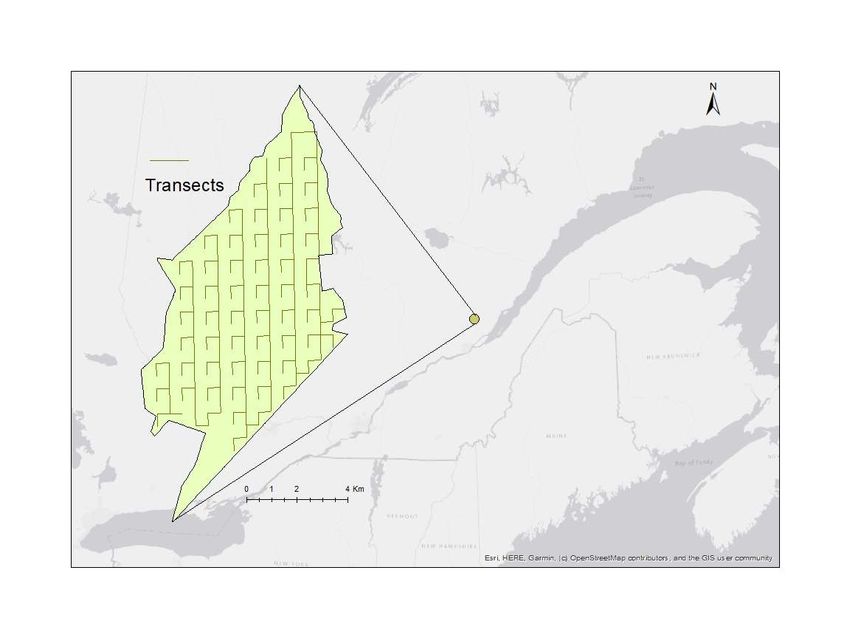
Importance de l’imagerie spatiale à haute résolution
pour la détection de l’effet de lisière en hiver
Mémoire
Yann Baril-Chauvette
Sous la direction de :
André Desrochers, directeur de recherche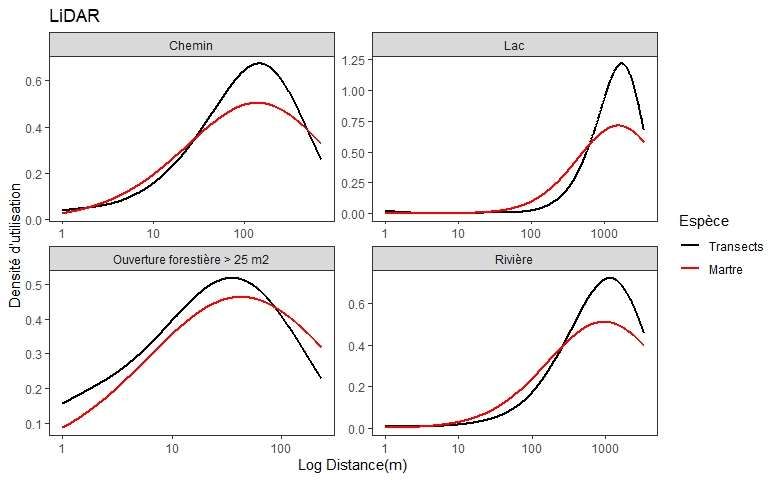
Résumé
En plus de leur taille, la configuration des coupes forestières influence la quantité de lisières dans un
paysage. La martre d’Amérique est considérée sensible aux opérations forestières puisqu’elle évite les
milieux ouverts comme les coupes forestières. Il n’est pas clair comment la martre réagit aux lisières
des milieux ouverts. Celles-ci peuvent offrir des proies mais également exposer les martres à leurs
prédateurs et aux intempéries. Nous avons analysé l’effet de ces lisières sur la répartition spatiale des
mammifères en hiver, avec une attention particulière à la martre d’Amérique. Nous avons également
comparé les résultats basés sur des images LiDAR de 2011 et 2016 et sur la couche thématique
écoforestière du gouvernement du Québec afin de déterminer l’effet de la résolution spatiale sur la
mesure des effets de lisière. Nous avons géolocalisé des pistes de martres et d’autres mammifères à
la forêt Montmorency, le long de transects répartis systématiquement, entre 2009 et 2018. Nous avons
comparé les distances des pistes et de points de référence disposés sur les transects aux lisières avec
des modèles linéaires généralisés. Les pistes de martres se retrouvaient davantage éloignées des
lisières d’ouvertures forestières que les points de référence, tout comme les espèces qu’elle
consomme. Avec les images LiDAR, la distance des pistes de martres aux lisières de chemins, de
rivières et de lacs ne se distinguait pas de celle des points de référence. Avec la couche thématique
écoforestière, nous n’avons trouvé aucune influence des lisières forestières, suggérant qu’une
délimitation précise des lisières est nécessaire pour en mesurer l’effet. L’évitement des lisières LiDAR
était obtenu en n’appliquant aucune taille minimale aux ouvertures utilisées pour leur délimitation. La
taille des coupes n’aurait donc pas d’influence sur l’évitement des lisières. Nous concluons qu’une
configuration spatiale réduisant la quantité de lisières augmenterait la superficie d’habitat de la martre
dans la sapinière à bouleau blanc de l’Est.
ii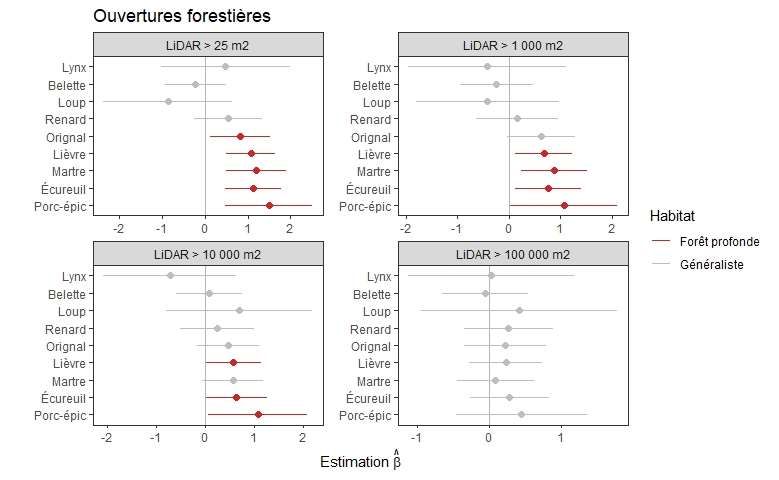
Abstract
Besides their size, spatial configuration of clear cuts determines the amount of edges in a landscape.
The American marten is considered sensitive to forest management since it avoids open areas such
as clear-cuts. It is unclear how marten respond to edges of open areas. Edges may represent foraging
opportunities, but also increased exposure to predators and severe weather. We analyzed the edge
effect on the spatial distribution of wintering mammals with a focus on marten. We compared the results
obtain by LiDAR images (2011 and 2016) and Quebec government’s photo interpretation data to
determine the impact of spatial resolution on measured responses to edges. We geolocated marten
and other mammal snow tracks in Forêt Montmorency, along systematically distributed transects,
between 2008 and 2018. We compared distances of snow tracks and reference points along transects
to edges with generalized linear models. Martens were found further from the edges of forest openings
than reference points, as were prey species. With LiDAR images, we found no association of martens
to roads, rivers and lakes edges. With the photo interpretation layer, we found no influence of forest
openings edges suggesting that a fine and precise delineation of edges is necessary to measure edge
effects. LiDAR edge avoidance was obtained by applying no minimum size to the openings used for
their delineation. Thus, the size of clear cuts would therefore not have an influence on edges avoidance.
We conclude that a spatial configuration with fewer edges would increase the area of suitable habitat
for martens in the eastern balsam fir-white birch forest.
iii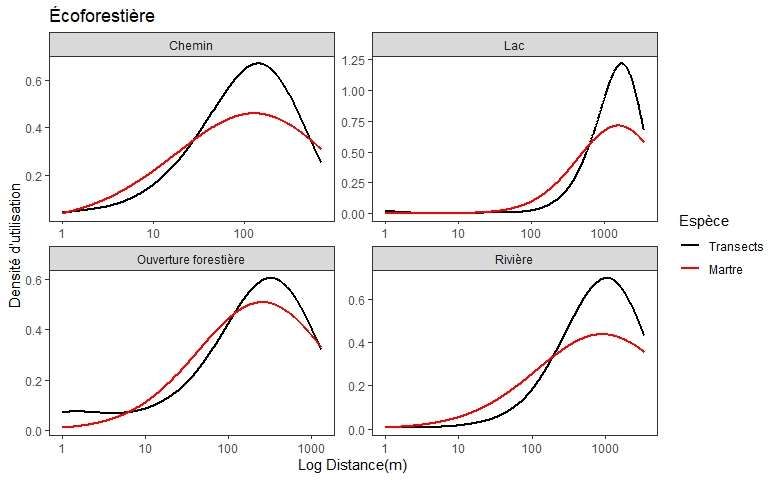
Table des matières
Résumé ................................................................................................................................................ ii
Abstract................................................................................................................................................ iii
Table des matières .............................................................................................................................. iv
Liste des figures ................................................................................................................................... vi
Remerciements ................................................................................................................................... vii
Introduction générale ........................................................................................................................... 1
Stratégie d’organisation de coupe en forêt boréale au Québec ....................................................... 1
Faune et lisières forestières ............................................................................................................. 3
Planification forestière et gestion intégrée des ressources .............................................................. 4
La martre d’Amérique ...................................................................................................................... 5
Habitat de la martre d’Amérique .................................................................................................. 5
Espèce d’intérêt socio-économique ............................................................................................. 6
Échelles et résolution spatiale.......................................................................................................... 7
Objectif et hypothèses ..................................................................................................................... 9
Chapitre 1. Importance de l’imagerie spatiale à haute résolution pour la détection de l’effet de lisière
en hiver .............................................................................................................................................. 10
Introduction .................................................................................................................................... 10
Méthodologie ................................................................................................................................. 13
Aire d’étude ................................................................................................................................ 13
Échantillonnage ......................................................................................................................... 14
Préparation des données ........................................................................................................... 15
Analyse des données ................................................................................................................. 17
Résultats ........................................................................................................................................ 18
Pistage ....................................................................................................................................... 18
Comparaison des tailles d’ouvertures forestières ...................................................................... 19
Comparaison des types de lisières ............................................................................................ 21
Comparaison des couches thématiques .................................................................................... 21
Discussion ..................................................................................................................................... 23
Conclusion générale .......................................................................................................................... 30
Implications pour l’aménagement forestier..................................................................................... 30
Capacité et limites du LiDAR ......................................................................................................... 32
Importance des variations régionales ............................................................................................ 32
Perspectives en conservation ........................................................................................................ 33
iv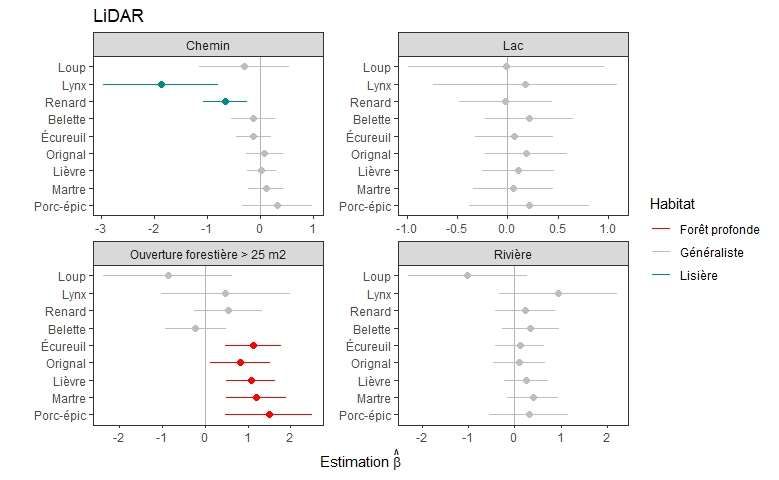
Bibliographie ...................................................................................................................................... 35
Annexes ............................................................................................................................................. 47
Annexe 1. Étendues en mètres des distances aux différents types de lisières selon le type de
couche thématique et les tailles minimales des ouvertures forestières LiDAR des points de
référence et des espèces échantillonnées. .................................................................................... 47
Annexe 2. Moyennes en mètres et écart avec les points de référence en pourcentage des
distances aux différents types de lisières (de lac, de rivière, de chemin ou forestière) selon le type
de couche thématique (écoforestière ou LiDAR) et les tailles minimales des ouvertures forestières
LiDAR (> 25 m2, > 1 000 m2, > 10 000 m2 et > 100 000 m2) des points de référence et des espèces
échantillonnées. Les moyennes de couleur verte indiquent un résultat significativement plus près
du type de lisière que les points de référence tandis que les moyennes de couleur rouge indiquent
un résultat significativement plus éloigné du type de lisière que les points de référence. Les
moyennes sans couleur indiquent des résultats non significatifs, ne se distinguant pas des points
de référence. .................................................................................................................................. 48
Annexe 3. Densité d’utilisation du territoire par la martre en fonction de la distance en log aux
types de lisières selon la couche thématique (LiDAR vs écoforestières) comparée aux points de
référence des transects ................................................................................................................. 49
Annexe 4. Script R pour générer les points de référence .............................................................. 50
Annexe 5. Script R de calcul des distances des pistes et points de référence aux lisières ............ 51
Annexe 6. Script R de calcul des coefficients du modèle linéaire généralisé ................................. 53
v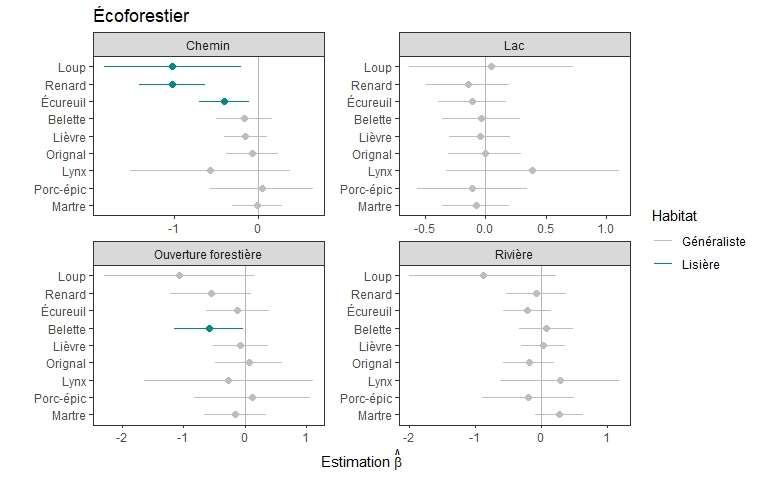
Liste des figures
Figure 1. Superficie des forêts de lisière et des forêts profondes selon le type d'organisation spatiale
des coupes forestières. ........................................................................................................................ 2
Figure 2. Localisation de la portion A de la Forêt Montmorency située à 80 km au nord de la ville de
Québec et des transects parcourus. .................................................................................................. 14
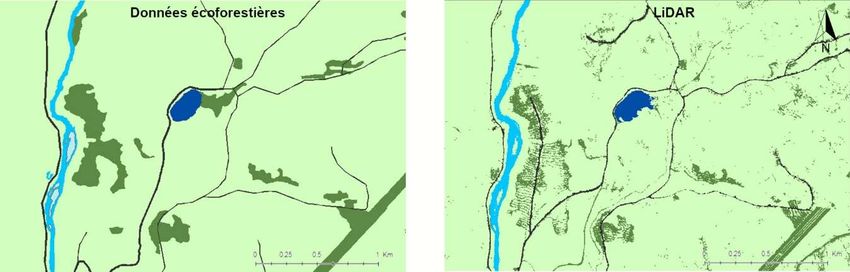
Figure 3.Comparaison de détection des ouvertures (forestières en vert foncé, lac en bleu foncé,
rivières en bleu pâle et chemins en noir) avec les données écoforestières (gauche) et les données
LiDAR (droite) pour la délimitation des lisières. ................................................................................. 16
Figure 4. Distances de pistes de mammifères aux lisières forestières selon la taille minimale des
ouvertures associées à ces lisières déterminées par l’imagerie LiDAR, comparées aux distances des
points de référence échantillonnées le long des transects. Les traits horizontaux représentent les
limites de confiance à 95 %. Les traits verticaux représentent l’hypothèse nulle (absence de
différence avec les points de référence). Les espèces sont classifiées de forêt profonde lorsque
l’estimation était significativement positive (rouge). Aucune estimation n’était significativement
négative. ............................................................................................................................................ 20
Figure 5. Distances des pistes de mammifères aux différentes lisières déterminées par l’imagerie
LiDAR, comparées aux distances des points de référence échantillonnées le long des transects. Les
traits horizontaux représentent les erreurs types. Les traits verticaux représentent l’hypothèse nulle
(absence de différence avec les points de référence). Les espèces sont classifiées de forêt profonde
lorsque l’estimation était significativement positive (rouge), et de lisière lorsque l’estimation était
significativement négative (vert)......................................................................................................... 22
Figure 6. Distances de pistes de mammifères aux différentes lisières déterminées par les données
écoforestières comparées aux distances des points de référence échantillonnées le long des
transects. Les traits horizontaux représentent les erreurs types. Les traits verticaux représentent
l’hypothèse nulle (absence de différence avec les points de référence). Les espèces sont classifiées
de lisière lorsque l’estimation était significativement négative (vert). Aucune estimation n’était
significativement positive. .................................................................................................................. 22
vi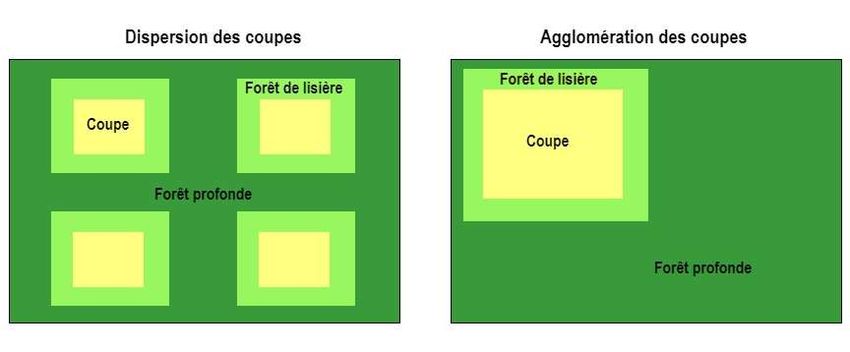
Remerciements
Ce mémoire a été réalisé avec la participation de beaucoup de gens et d’organismes. Je tiens tout
d’abord à remercier mon comité d’encadrement pour le soutien, les précieux conseils et discussions
qui ont permis autant à ce projet qu’à moi-même d’évoluer pour le mieux. Merci à André Desrochers,
mon directeur, spécialement pour sa disponibilité, son transfert de connaissances en analyse et
statistique et pour son esprit critique. Merci également à Pauline Suffice pour ses conseils constructifs
nombreux autant sur le plan académique que professionnel ainsi que pour son expertise des
mustélidés. Je remercie aussi la Fédération des Trappeurs Gestionnaires du Québec qui a initié le
projet et a permis sa réalisation avec le financement de la Fondation de faune et son programme de
gestion intégrée des ressources pour l’aménagement durable de la faune en milieu forestier. J’offre
également mes remerciements à mes évaluateurs externes, Marianne Cheveau et Jean-Pierre
Tremblay pour avoir pris de leur précieux temps afin de participer à l’amélioration de ce mémoire.
Finalement, je tiens à remercier Martine Lapointe et tous les bénévoles qui ont participé au programme
de suivi de pistage de mammifères de la forêt Montmorency.
viiIntroduction générale
Stratégie d’organisation de coupe en forêt boréale au Québec
La Stratégie d’aménagement durable des forêts produite par le Ministère de la Forêt, de la Faune et
des Parcs (ci-après nommé MFFP) en vertu de l’article 11 de la Loi sur l’aménagement durable du
territoire forestier repose sur l’aménagement écosystémique (MFFP, 2015). Plusieurs auteurs ont tenté
de décrire précisément cette approche qui consiste à intégrer les connaissances sur le fonctionnement
des écosystèmes lors de l’aménagement forestier. Il serait difficile de donner une définition universelle
de l’aménagement écosystémique selon Fyles (2008). Celui-ci la décrit comme une approche qui vise
à maintenir les écosystèmes jugés sains et résilients en misant sur une diminution des écarts entre les
paysages naturels et ceux qui sont aménagés. En diminuant ces écarts, on maintiendrait les fonctions
de l’écosystème et conserverait les bénéfices sociaux et économiques à long terme. La résilience ferait
référence à la capacité (Gunderson & Holling, 2003; Holling, 1973) et à la rapidité (Begon et al., 1996;
Perry & Amaranthus, 1997) d’un écosystème à se rétablir après une perturbation. Il est ainsi possible
de mesurer certains niveaux de résilience, mais toutefois difficile d’en attribuer une cible universelle.
Le problème est particulièrement évident en ce qui concerne le caractère sain d’un écosystème. Une
multitude de définitions et d’indicateurs ont émergé depuis quelques années afin de décrire ce concept
holistique. Les indicateurs sont variés puisqu’ils décrivent plusieurs angles de ce concept (Kruse,
2019). Le terme « sain » entraine donc une certaine confusion puisqu’il est interprété de nombreuses
manières. Néanmoins, la stratégie retenue par l'aménagement écosystémique pour maintenir les
écosystèmes sains et résilients, consiste en pratique, à imiter les perturbations naturelles lors de la
récolte forestière.
En forêt boréale, la stratégie de coupe en mosaïque permet d’imiter les épidémies d’insectes en
réduisant la dimension des aires des coupes totales et en les dispersant davantage dans le territoire
(Bélanger, 1992). On vise à répartir les coupes dans l’espace, mais également, dans le temps (MFFP,
2021b). Il en résulte une mosaïque de peuplements équiens de compositions et d’âges différents. Ce
type de coupe permettrait à la fois de maximiser la récolte, de maintenir une qualité visuelle, de
protéger les sources hydriques, mais aussi d’améliorer les habitats fauniques de plusieurs espèces
1entre autres par le maintien d’un couvert de fuite et d’une variété de hauteurs de peuplements
(Bélanger, 1999).
Toutefois, la dispersion d’ouvertures du système de coupes en mosaïque pourrait accentuer les
différences d’organisation spatiale entre la forêt naturelle et la forêt aménagée sur un territoire (Belleau,
2012; Tittler, 2010). Pour des raisons purement géométriques, la dispersion des coupes en mosaïque
entraine notamment une quantité plus élevée de lisières pour une même superficie de coupes (Figure
1). Cette dispersion oblige également l’augmentation du nombre de chemins pour accéder à chaque
zone de coupe (MFFP, 2017). Le MFFP a lui-même mentionné que la coupe en mosaïque engendrerait
la perte et la fragmentation des habitats de forêts à couvert fermé, en plus de pertes de forêts
profondes. Elle pourrait ainsi limiter les déplacements des espèces associées à ces types de forêts.
Figure 1. Superficie des forêts de lisière et des forêts profondes selon le type d'organisation spatiale des coupes forestières.
(Inspiré de MFFP)
Dans les domaines de la sapinière et de l’érablière, c’est l’approche par coupes en mosaïque qui est
actuellement en place. Toutefois, des travaux sont en cours dans le domaine de la sapinière afin
d’élaborer un modèle de répartition des coupes qui soit davantage adapté à ce territoire. Certaines
parties du domaine de la sapinière à bouleau jaune et à bouleau blanc font l’objet d’une dérogation à
la coupe en mosaïque (MFFP, 2019a). Ces zones de dérogation intègrent plutôt une approche
d’aménagement qui repose sur une plus grande délimitation des massifs forestiers. L’approche est
déjà utilisée et adoptée dans le domaine de la pessière à mousse (Bureau du forestier en chef, 2018).
Une échelle spatiale plus grossière d’aménagement permet la concentration des coupes et
2l’augmentation de la taille des blocs résiduels. Il deviendrait alors possible de maintenir une dominance
de forêt à couvert fermé à l’échelle du paysage en plus de favoriser la présence de forêt profonde à
l’échelle du paysage (MFFP, 2019a).
Faune et lisières forestières
Du point de vue de la faune, une lisière ou un écotone est le point de rencontre entre deux différents
habitats. Elle est définie par Turner et Gardner (2015) comme « la portion d’un écosystème ou d’un
type de couvert près de son périmètre et dans laquelle les conditions environnementales peuvent
différer de la portion intérieure de l’écosystème en question ». En milieu forestier, on peut distinguer la
lisière comme la limite qui sépare ces deux habitats et la forêt de lisière comme la portion forestière
qui subit les effets de cette lisière. En principe, l’intensité de l’effet de lisière dépend surtout des écarts
d’âge entre les peuplements qui les délimitent (Harris, 1984). En pratique, les lisières sont souvent
étudiées comme la limite entre une portion boisée et non boisée (Saunders et al., 1991). La forêt de
lisière est davantage exposée et subit par conséquent, une plus grande influence de plusieurs
éléments abiotiques incluant la lumière, la température, l'humidité et le vent (Chen et al., 1995;
Saunders et al., 1991). Ces perturbations induisent des changements de composition et de structure
(Harper et al., 2005; Marozas et al., 2005). Un gradient de ces impacts s’exerce selon la distance à la
lisière (Harper et al., 2005). Une forêt de lisière se termine théoriquement à l’endroit où ces facteurs
cessent d’exercer une influence et devient la portion de forêt profonde (Turner & Gardner, 2015). Ces
facteurs et leurs influences sont difficiles à mesurer en pratique et la délimitation entre forêt de lisière
et forêt profonde est donc assez ardue. Le MFFP utilise présentement une distance fixe de 75 m pour
délimiter les 2 types de forêts en sapinière (MFFP, 2019a). Toutefois, il est facile de croire que la
distance réelle peut varier en fonction des caractéristiques de la forêt, mais également, en fonction de
la perception des différentes espèces fauniques qui en sont affectées. L’échelle et l’intensité à laquelle
la lisière affecte une espèce dépendraient largement des caractéristiques intrinsèques de cette même
espèce. Ainsi, cette mesure généraliserait et simplifierait un effet beaucoup plus complexe en réalité.
L’effet d’une lisière sur une espèce dépendrait également de l’amplitude de la rupture (abrupte ou
graduelle) et du contraste entre les habitats en plus de sa propre capacité d’adaptation (Larivière,
2003). On peut considérer trois types de spécialisations liées aux lisières chez la faune forestière. Il y
a d’abord les espèces de lisières qui par définition bénéficient de leur présence. Il y a également les
3espèces généralistes qui sont liées aux forêts résiduelles, mais sur lesquelles les lisières ne présentent
pas d’effet. Finalement, il y a les espèces spécialistes des forêts profondes, liées aux forêts résiduelles
pour lesquelles les lisières ont un effet négatif. Une espèce reconnue comme spécialiste de forêts
profondes est donc beaucoup plus affectée par la création de lisières.
Planification forestière et gestion intégrée des ressources
Au Québec, lors de la planification forestière, le ministre collabore avec les tables locales de gestion
intégrée des ressources et du territoire (TLGIRT) afin de concilier les travaux d’aménagement avec les
différents utilisateurs de la forêt (MFFP, 2018). Un des enjeux de la planification forestière intégrée est
l’intégration des besoins d’habitats fauniques. Pour ce faire, le ministère a élaboré un système de suivi
d’espèces focales, sensibles à des enjeux d’aménagement forestier dans sa Stratégie d’aménagement
durable des forêts du Québec (défi 2, orientation 2, objectif 3; MFFP, 2015).
Le but est de s’assurer que les nouvelles conditions environnementales modifiées par l’aménagement
forestier demeurent favorables aux espèces et à la diversité biologique. Les différents enjeux
d’aménagement écosystémique sont la structure d’âge des forêts, la composition végétale, la structure
interne et le bois mort, les milieux humides et riverains, le maintien des forêts brûlées ainsi que l’enjeu
d’organisation spatiale (Cheveau, 2015). Les TLGIRT peuvent également proposer davantage de
composantes et d’enjeux aux habitats fauniques selon les caractéristiques locales et les
préoccupations fauniques spécifiques à leur territoire, notamment les besoins en matière de gestion
de la faune. Les préoccupations quant aux animaux à fourrure et au gibier sont ainsi discutées
davantage aux TLGIRT. L’intégration de ces nouveaux enjeux est importante étant donné que la
sélection des espèces focales entraine une certaine simplification des besoins de l’entièreté de la faune
forestière. Le MFFP a choisi la martre d’Amérique (Martes americana) (ci-après nommée martre)
comme espèce focale pour les domaines bioclimatiques de la pessière et de la sapinière concernant
les principaux enjeux de structure d’âge, de composition végétale, de structure interne et de bois mort
ainsi que pour l’enjeu d’organisation spatiale (Cheveau, 2015).
4La martre d’Amérique
Habitat de la martre d’Amérique
La martre possède un domaine vital qui peut s’étendre de 0.6 km2 à plus de 20 km2 (Buskirk &
McDonald, 1989). Dans la sapinière, la taille des domaines vitaux des martres est d’approximativement
5 km2 (Potvin & Courtois, 1998). Un habitat est défini par Morrison et al. (2012) comme « un espace
combinant des ressources comme de la nourriture, un couvert ou une source d’eau, ainsi que des
conditions environnementales favorisant la présence d’individus d’une espèce donnée et permettant à
ces individus de survivre et se reproduire ». L’habitat de la martre doit ainsi contenir une abondance
de proies, un couvert contre les prédateurs, une protection face aux intempéries ainsi que des sites de
repos et de tanières (Joyce, 2013; Thompson & Harestad, 1994). Dans l’est de l’Amérique du Nord, le
régime alimentaire de la martre est très saisonnier. En été, la martre est plutôt opportuniste et peut se
nourrir de fruits et d’insectes bien que les campagnols seraient sa proie de prédilection (Lachowski,
1997; Soutiere, 1979). En hiver, son régime alimentaire est essentiellement carnivore. À cette période,
les passages sous-nivéaux deviennent importants pour accéder aux micromammifères (Hargis &
McCullough, 1984; Sherburne & Bissonette, 1994; Thompson & Colgan, 1990). Toutefois, à cette
période, de plus grosses proies, principalement le lièvre, mais aussi les écureuils constituent les
sources principales de l’alimentation de la martre possiblement due à l’apport énergétique qu’ils
procurent (Fuller & Harrison, 2005; Martin, 1994; Thompson & Colgan, 1990).
On estime que l’hiver est la saison critique pour la martre puisque l’accès et la disponibilité de la
nourriture diminuent. De plus, son métabolisme est rapide et elle détient peu de réserves de gras, ce
qui l’oblige à être un prédateur actif (Buskirk & Harlow, 1989; Mustonen & Nieminen, 2012). Pour limiter
la perte d’énergie liée à la thermorégulation, la martre se réfugie dans des abris sous-nivéaux qui lui
fourniraient également des sites de repos (Buskirk & Harlow, 1989). En plus de leur permettre de se
protéger contre le froid, ces accès formés principalement par la présence de débris ligneux et de
branches basses donnent un accès aux proies sous-nivéales (Jung et al., 2021; Raine, 1987; Snyder
& Bissonette, 1987; Soutiere, 1979) et une protection face aux prédateurs (Drew, 1995). Les tanières
des martres se retrouvent également dans les milieux forestiers complexes contenant de nombreux
débris ligneux (Joyce, 2013). On retrouve généralement un plus grand nombre de ces accès dans les
5peuplements matures à couvert résineux où la neige est moins profonde (Wiebe et al., 2014) et où la
quantité de débris ligneux est plus élevée (Sherburne & Bissonette, 1994). Dans ce type de
peuplement, la température est également plus modérée et le couvert offre possiblement une
protection face aux prédateurs (Godbout & Ouellet, 2010).
Ainsi, beaucoup d’auteurs ont longtemps associé la martre aux forêts matures à dominance résineuse
(Buskirk & Powell, 1994; Strickland & Douglas, 1987). Toutefois, il semble qu’elle soit plutôt associée
à une complexité de la structure verticale et horizontale des peuplements (Chapin et al., 1997). Ce
type de structure se retrouve bien souvent en forêts matures et surannées, mais il est aussi possible
de la retrouver dans de plus jeunes peuplements atteints par une épidémie d’insecte ou un chablis par
exemple (Potvin et al., 2000). Les conditions nécessaires à la martre ne se retrouvent pas dans les
zones de coupes et les peuplements en régénération principalement dus à l’ouverture du couvert et
elle éviterait ces milieux (Buskirk & Powell, 1994; Poole et al., 2004; Thompson & Harestad, 1994). La
martre est donc souvent reconnue comme une espèce spécialiste des forêts profondes (Hargis et al.,
1999; Potvin et al., 2000). Toutefois, sa relation avec les lisières semble complexe. Ces milieux ne
sembleraient pas contenir un couvert idéal à sa protection (Ruggiero, 1994; Drew, 1995) comme c’est
le cas en forêt profonde, mais pourraient contenir davantage de proies (Cheveau, 2010; Vigeant-
Langlois & Desrochers, 2011) et un compromis pourrait alors émerger concernant la fréquentation des
lisières par la martre. La martre est considérée comme une espèce grandement sensible aux
perturbations forestières anthropiques (Chapin et al., 1998; Payer & Harrison, 1999; Potvin et al., 2000;
Thompson, 1991). C’est d’ailleurs pourquoi elle a été désignée comme espèce focale pour les
différents enjeux d’aménagement forestier (Cheveau, 2015).
Espèce d’intérêt socio-économique
Si la martre détient dorénavant un rôle d’espèce focale pour l’aménagement des forêts par le
gouvernement du Québec, dans le cas de certains peuples autochtones, elle détenait déjà un rôle
semblable. Pour les Innus de Pessamit, elle est perçue comme une sentinelle de l’état de santé des
forêts (Ménard, 2019). Pour plusieurs peuples autochtones, le piégeage de la martre représentait une
source de revenus très importante historiquement et beaucoup d’individus continuent à pratiquer cette
6activité culturelle malgré la chute du marché (Todd & Boggess, 1987; White et al., 2015). Récoltée
d’abord majoritairement par les piégeurs autochtones, l’engouement pour la martre a augmenté
radicalement avec le début de la traite des fourrures avec les Européens (Innis, 1962). Le prix élevé
de sa fourrure couplé avec un succès élevé de capture causé par son comportement curieux en ont
fait une cible prisée. D’ailleurs, suivant la colonisation européenne et la popularité de la traite des
fourrures aux XVIIe, XVIIIe, et XIXe siècles, les populations d’animaux à fourrure, incluant la martre,
auraient connu une importante chute en Amérique du Nord. On estime que cela résulte principalement
de la surexploitation due à l’absence de quotas pour ces espèces (White et al., 2015). Dans le cas de
la martre, cela a mené certaines populations près de l’extinction comme c’est le cas pour Terre-Neuve,
la Nouvelle-Écosse et quelques états américains (Thompson, 1991). C’est aussi possiblement une
cause majeure de la disparition de la population de l’Île-du-Prince-Édouard (Sobey, 2007). Toutefois,
à la suite des efforts de conservation et de réintroduction mis en place depuis la fin du XX e siècle, la
plupart des populations se sont stabilisées (Thompson, 1991; White et al., 2015). La tendance actuelle
est en diminution des populations selon l’UICN dans l’ensemble de son aire de répartition, mais
l’espèce conserve un statut de préoccupation mineure. Les principales menaces demeurent la récolte
d’individus ainsi que la perte d’habitat causée par la récolte forestière (Helgen & Reid, 2016). La martre
est aujourd’hui piégée dans la plupart de son aire de répartition sauf pour certains secteurs où elle a
été surexploitée comme en Nouvelle-Écosse (Fur harvesting regulations, N.S. Reg. 285/2015, Art. 6.5).
Au Québec, il n’y a jamais eu d’épisode de surexploitation reconnue et la martre est toujours piégée
dans l’ensemble de la province (MFFP, 2014). Le piégeage ne représente plus actuellement une
activité aussi lucrative qu’auparavant pour les piégeurs québécois et pour la majorité d’entre eux, il
s’agit d’un loisir qui engendre davantage de dépenses que de revenus (MFFP, 2019b).
Échelles et résolution spatiale
En écologie du paysage, les échelles de perception sont reconnues comme un concept fondamental.
Les processus écologiques ou objets d’études sont influencés par différentes échelles spatiales
(Desrochers et al., 2010; Wiens, 1989). Ces échelles sont caractérisées par la mesure de la zone
d’étude physique ainsi que par le niveau de résolution des données spatiales (Turner & Gardner, 2015).
Au Québec, les données sur la forêt proviennent principalement des couches écoforestières qui sont
générées à partir de photo-interprétation. Cette technique vise à déterminer les caractéristiques de la
7forêt à partir de photos aériennes. La résolution spatiale de celle-ci est à l’échelle du peuplement et la
résolution minimale d’un peuplement est de 40 000 m2 (4 ha), tandis que la résolution minimale pour
une perturbation est de 20 000 m2 (2 ha, MFFP, 2021a). Il s’agit donc de l’échelle la plus fine que peut
générer ce type de données. Or, certaines espèces ont des besoins d’habitat particuliers à fine échelle
qui peuvent être effacés lorsque perçus à une échelle plus grossière résultant potentiellement à des
conclusions biaisées (Bissonette et al., 1997; McCann et al., 2014; Shirk et al., 2012; Turner et al.,
1989).
Depuis quelques années, des produits spatiaux provenant du LiDAR (Light Detection and Ranging)
ont émergé. Le LiDAR est une technologie de renvoi laser permettant de mesurer à haute résolution,
les caractéristiques de la forêt en trois dimensions (Lefsky et al., 2002). La résolution spatiale du
dernier inventaire est de 1 ou 2 mètres carrés selon le type de modèle (MFFP, 2021c). Cette résolution
est donc 20 000 fois plus précise que celle de la couche écoforestière. Trois types de modèles dérivés
de cette technologie sont disponibles au Québec soit le modèle numérique de terrain, les pentes ainsi
que le modèle de hauteur de canopée.
En superposant des données de présence ou de déplacement d’espèces aux données LiDAR, il est
possible d’obtenir un portrait plus précis de l’utilisation et de la sélection de l’habitat des espèces (Ganz
et al., 2019). La disponibilité des images satellites et LiDAR a permis de faciliter l’utilisation de
différentes échelles en écologie du paysage et mieux comprendre les relations écologiques entre ces
échelles (Ackers et al., 2015). Il est donc possible d’évaluer facilement à quelle échelle l’effet de
différents paramètres d’habitat s’applique, en plus d’en augmenter la précision (Tweedy et al., 2019).
La meilleure résolution permet ainsi de mesurer à fine échelle différentes caractéristiques de la forêt
(Tweedy et al., 2019) et ainsi en calculer leur utilisation et sélection à ces échelles (Vierling et al.,
2008). En sélection d’habitats, le climat est souvent un facteur déterminant à une échelle grossière
(Pearson & Dawson, 2003) tandis qu’à plus fine échelle, c’est entre autres, la structure de la végétation
qui devient davantage impliquée (Zellweger et al., 2013). Une conception en trois dimensions de cette
structure végétale permettrait de mieux identifier, comprendre et conserver certains attributs de la forêt
(Davies & Asner, 2014). La complexité des données LiDAR retarde le développement des nouveaux
produits, mais le produit est prometteur (Bakx et al., 2019). Par exemple Joyce et al. (2019) ont
8démontré qu’il est possible de détecter la présence de grands débris ligneux à partir des données
LiDAR avec une densité de points allant jusqu’à 24 points/m 2. Les données disponibles présentement
au Québec ont une densité variant de 1 à 8 points/m2 (MFFP, 2021c). Néanmoins, les produits
disponibles dérivés du LiDAR permettent déjà d’obtenir de nouvelles informations potentiellement
utiles à notre compréhension des relations entre les espèces et leurs habitats.
Objectif et hypothèses
Le but de cette recherche était de déterminer si la répartition spatiale de mammifères, en particulier la
martre, est affectée par la distance à une lisière en contexte hivernal. La martre étant considérée
sensible aux perturbations forestières anthropiques, nous avons testé si elle évite la proximité de ces
perturbations. Afin de valider notre hypothèse, nous avons prédit que les pistes de martre se
retrouveraient plus éloignées des lisières, dans la forêt profonde, que ce qui serait attendu en l’absence
d’effet de lisière. Nous avons aussi prédit que l’association à la forêt profonde serait plus prononcée
chez la martre que chez les autres mammifères. Nous avons également prédit que plus la taille des
ouvertures forestières était grande, plus les pistes de martre s’en trouveraient éloignées. Finalement,
nous avons comparé les résultats en fonction de la couche thématique utilisée, soit LiDAR ou
écoforestière, en raison de leurs résolutions différentes. Les résultats de notre étude permettront de
mettre en lumière le rôle et les impacts potentiels des résolutions spatiales utilisées en écologie du
paysage. De plus, nos conclusions amèneront des arguments relatifs à la présence de lisières en forêt
et à l’enjeu d’organisation spatiale en aménagement forestier durable au Québec.
9Chapitre 1. Importance de l’imagerie spatiale à haute
résolution pour la détection de l’effet de lisière en hiver
Introduction
Les activités de récoltes forestières entrainent une altération du paysage pouvant causer la perte et la
fragmentation des habitats de la faune forestière et participer à leur déclin (Fahrig, 2003; Lindenmayer
et al., 2006). La création de lisières forestières induite par les coupes pourrait également ajouter à la
dégradation des habitats et aussi participer à ce déclin (Gascon et al., 2000; Harper et al., 2005;
Saunders et al., 1991). Du point de vue de la faune forestière, une lisière peut être définie comme la
limite entre deux habitats distincts comme un milieu ouvert et forestier (Matlack & Litvaitis, 1999;
Saunders et al., 1991). La portion de forêt résiduelle adjacente à la lisière est influencée par des
variations de facteurs abiotiques comme la lumière, le vent, la température et l’humidité qui diffèrent
de la forêt profonde (Chen et al., 1995; Saunders et al., 1991). Les conséquences incluent entre autres
davantage de chablis et de débris ligneux causés par l’augmentation des rafales de vent ainsi qu’un
changement de composition végétale et l’intensification de la régénération en sous-bois causés par
l’augmentation de la lumière pénétrante (Saunders et al., 1991). L’effet de lisière correspond à ces
différentes variations et fluctue en fonction de la distance à l’ouverture, diminuant progressivement
dans le temps ainsi que dans l’espace (Harper et al., 2005; Harris, 1984; Matlack, 1994). Toutefois, la
création de lisières forestières n’est pas exclusive aux activités anthropiques. En forêt boréale, le
régime de perturbations repose principalement sur les feux, les épidémies d’insectes, ainsi que les
chablis (Bonan & Shugart, 1989). Ces perturbations entrainent des trouées de différentes dimensions
et donc, la présence de lisières naturelles fait partie de ce type de paysage. L’impact de la création de
lisières anthropiques sur la faune pourrait donc être moins important que dans d’autres types de forêts.
Toutefois, la structure et donc, l’effet des lisières naturelles et anthropiques peuvent différer (Harper et
al., 2004; Larrivée et al., 2008). La densité de lisières dans le paysage peut également influencer l’effet
sur la faune (Fletcher, 2005). La densité de lisières en aménagement forestier est influencée par la
taille, mais également, par la configuration spatiale des coupes. L’agglomération des coupes
permettrait de réduire la quantité de lisières et de conserver de plus larges blocs de forêts résiduelles
profitables pour la faune sensible aux lisières (Boucher et al., 2011). L’effet des lisières sur la faune
peut varier selon l’intensité du contraste entre les deux milieux, mais également selon les
10caractéristiques intrinsèques de l’espèce (Larivière, 2003) et certaines espèces boréales pourraient
aussi en profiter.
Parmi les espèces dites de lisières, plusieurs cervidés comme les cerfs et les orignaux y sont associés
parce qu’ils utilisent à la fois les milieux ouverts et les forêts avoisinantes, respectivement comme lieux
d’alimentation et de couvert (Månsson et al., 2012; Ruzicka et al., 2010). La présence de lisières a
longtemps été considérée favorable en gestion de la faune puisqu’elle augmenterait entre autres la
densité et la diversité de ces espèces gibiers (Leopold, 1933). La végétation basse des lisières
représente un garde-manger attirant pour plusieurs espèces d’herbivores. Les lièvres bénéficieraient
également de ce garde-manger, en plus du couvert que leur procure les lisières (Forsey & Baggs,
2001; Litvaitis et al., 1985; Wolff, 1980). Ce couvert serait essentiel puisque la prédation pourrait être
plus forte dans ces milieux (Dijak & Thompson, 2000; Forsey & Baggs, 2001). En plus des facteurs
abiotiques mentionnés plus haut, la pression exercée par les herbivores influence la structure, la
densité et la composition de ces habitats (Cadenasso & Pickett, 2001). De nombreuses espèces
d’oiseaux fréquentent également les lisières (Bender et al., 1998) et quelques-unes parmi celles-ci en
seraient des spécialistes en forêt boréale, comme Mésangeai du Canada (Perisoresu canadensis ;
Ibarzabal et Desrochers 2000). Par ailleurs, certaines espèces boréales sont des utilisateurs facultatifs
de lisières, comme la mésange à tête brune (Poecile hudsonicus) en hiver, dont l’association aux
lisières lors des journées de grand froid diffère de celle lors de journées plus clémentes (Hadley &
Desrochers, 2009).
L’évitement des lisières peut s’expliquer par l’absence d’attributs recherchés par une espèce ou par
les risques encourus à sa fréquentation. Pour plusieurs espèces proies, la diminution du couvert
forestier entraine une hausse du risque de prédation. On sait depuis plusieurs décennies que
l’ouverture de ces milieux peut faciliter les déplacements des prédateurs (Bider, 1968; Dijak &
Thompson, 2000; Forsey & Baggs, 2001). Dans le cas des oiseaux, la prédation et le parasitisme des
nids près des lisières varierait selon le type de paysage, mais augmenterait dans les forêts fragmentées
(Batáry & Báldi, 2004; Paton, 1994). Les lisières offrent parfois un garde-manger intéressant, mais
11aussi, une augmentation du risque de prédation. Leur fréquentation peut donc entrainer un compromis
chez certaines espèces.
Pour un mésoprédateur comme la martre d’Amérique, la relation avec les lisières devient complexe,
car ces espèces doivent composer simultanément avec leur statut de prédateur et de proie. Certains
résultats présentent leur utilisation principalement comme habitat d’alimentation puisque les proies,
notamment le lièvre et l’écureuil y seraient plus abondantes (Cheveau, 2010; Vigeant-Langlois &
Desrochers, 2011). D’autres, en revanche, rapportent une réticence à l’utilisation des zones de densité
élevée de lisières (Hargis et al., 1999; Potvin et al., 2000). Les risques de prédation augmenteraient
beaucoup plus avec l’ouverture de la canopée et principalement dans les milieux ouverts, tant pour
ses prédateurs aviaires que terrestres (Ruggiero, 1994). Ce risque exercerait une influence sur
l’utilisation de l’habitat par la martre (Drew, 1995). Aussi, davantage d’énergie doit être allouée à la
thermorégulation en lisière en hiver dû à l’exposition aux intempéries. L’utilisation des lisières pourrait
ainsi autant profiter que nuire à la martre. Selon Buskirk et Powell (1994), l’utilisation des lisières serait
dépendante des caractéristiques des deux habitats adjacents. Le type de lisière exercerait ainsi une
influence sur sa fréquentation. Si cette utilisation n’est pas si claire, Cheveau (2010) et Frouin (2011)
indiquent toutefois que la martre adopte un déplacement généralement parallèle aux lisières en hiver
et qu’elles agissent donc comme une forme de barrière, la zone non boisée n’étant pas utilisée.
Pour bien comprendre la perception de l’effet des lisières par la faune, le choix de l’échelle et le niveau
de résolution des données utilisées sont importants puisqu’ils peuvent engendrer des résultats
imprécis ou des biais s’ils ne sont pas adéquats (McCann et al., 2014). Au Québec, les données sur la
forêt ont évolué et la nouvelle technologie LiDAR de renvoi laser permet d’obtenir une image d’une
résolution d’un mètre carré comparativement aux données écoforestières utilisées précédemment où
la taille minimale d’une perturbation était de 2 hectares (20 000 m2). Les besoins en habitat de la martre
se rencontrent à différentes échelles, mais plusieurs d’entre eux se matérialisent à des échelles
inférieures au peuplement forestier comme à l’échelle du micro-habitat (Godbout & Ouellet, 2010;
Nams & Bourgeois, 2004). D’ailleurs, Thompson et al. (2012) indique que dans le cas de la martre,
peu de facteurs limitants de son habitat se présentent à l’échelle du peuplement. Les raisons possibles
12Vous pouvez aussi lire