L'utilisation d'espèces indicatrices dans un contexte de changement climatique : Cas des oiseaux de l'est de la forêt boréale canadienne
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
L'utilisation d'espèces indicatrices dans un contexte de changement climatique : Cas des oiseaux de l'est de la forêt boréale canadienne Mémoire Alexandre Terrigeol Maîtrise en biologie - avec mémoire Maître ès sciences (M. Sc.) Québec, Canada © Alexandre Terrigeol, 2021
Résumé Plusieurs contraintes peuvent entraver le suivi à large échelle nécessaire à l’évaluation et à la mise en place de mesures pour atténuer la perte de biodiversité, notamment en milieu nordique. L’utilisation d’espèces indicatrices d’assemblages est une méthode qui permet de simplifier le suivi de la biodiversité à partir d’un nombre limité d’espèces. Malgré des études prometteuses à fines échelles, l’efficacité d’une telle pratique sur de vastes étendues spatiales demeure incertaine. À l’aide de 22 041 points d’écoutes provenant d’atlas d’oiseaux nicheurs, nous avons évalué l’efficacité des espèces indicatrices d’oiseaux à prédire de manière cohérente la richesse spécifique malgré les variations climatiques observées sur 1,75 M km2 de forêt boréale. Afin de tester l’effet des conditions climatiques sur les assemblages d’espèces, nous avons séparé notre zone d’étude en huit régions climatiques relativement similaires. À l’aide d’analyses par modèle nul sur la co-occurrence des espèces, nous avons identifié le nombre minimal d’espèces indicatrices nécessaires pour prévoir 70 % de la variation de richesse spécifique dans chacune des régions climatiques. Bien que nous ayons réussi à identifier un nombre restreint d’espèces indicatrices par région climatique, nous avons également montré que les espèces indicatrices variaient largement entre les régions. Les espèces qui conservaient leur capacité à être indicatrices dans plusieurs régions climatiques étaient également représentatives d’assemblages d’espèces différentes. Cette étude montre que les espèces identifiées comme indicatrices pourraient ne plus indiquer le même assemblage d’espèces dans un avenir proche. L'utilisation d’indicateurs à large échelle s'accompagne donc du défi de devoir déterminer sous quelles nouvelles conditions un ensemble d'indicateurs doit être remplacé, puis d’identifier le nouvel ensemble approprié. ii
Abstract Many constraints can impede the broad-scale monitoring needed to assess and mitigate human-induced loss of biodiversity. The use of indicator species of species assemblage can simplify the monitoring of biodiversity based on a limited number of species. Despite promising fine-scale studies, it remains unclear whether indicator species remain effective over broad spatial extents. Using 22,041 point-counts coming from breeding bird atlases, we assessed the value of indicator species to consistently predict bird species richness despite variation in climate conditions observed over 1.75 M km2 of boreal forest. To test for the effect of climate conditions on species assemblages, we partitioned our study area into eight relatively similar climatic regions. Using null model analyses of species co-occurrence, we identified the minimum number of indicator species required to predict 70% of the variation in species richness in each of the climatic regions. Although we were able to identify a small number of indicator species in each climatic region, we observed that the identity of indicator species varied strongly among regions. When species remained an indicator in several climatic regions, they were representative of different species assemblages. This study shows that species currently identified as indicators may no longer indicate the same species assemblage in a near future. Broad-scale use of indicators thus comes with the challenge of having to determine under which new conditions a given set of indicators must be replaced by another, and by which one. iii
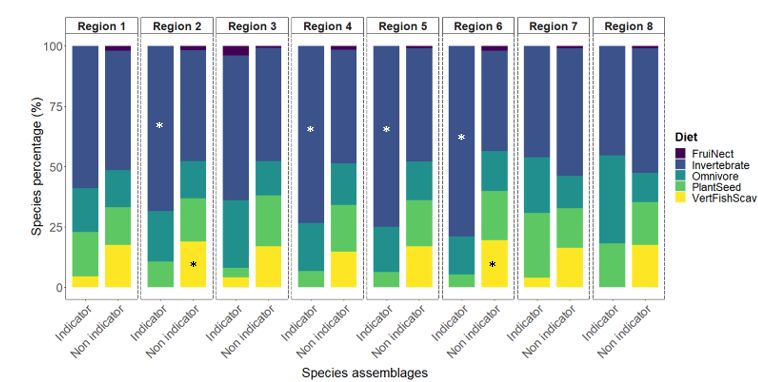
Table des matières Résumé ..............................................................................................................ii Abstract ........................................................................................................... iii Liste des tableaux ............................................................................................ vi Liste des figures ..............................................................................................vii Remerciements ................................................................................................ ix Avant-propos ................................................................................................... xi Introduction ...................................................................................................... 1 Contexte général ............................................................................................................................ 1 Les changements climatiques et la forêt boréale ........................................................................ 2 Résistance des assemblages d’espèces aux changements de la forêt boréale ........................... 3 L’utilisation d’indicateurs ............................................................................................................ 5 Les modèles nuls pour décrire les patrons de co-occurrence .................................................... 6 L’utilisation de la faune aviaire ................................................................................................... 7 Objectif de l’étude ......................................................................................................................... 8 Chapitre principal - On the efficiency of indicator species for broad-scale monitoring of bird diversity under climate change .................................... 10 Résumé ......................................................................................................................................... 10 Abstract ........................................................................................................................................ 11 Introduction ................................................................................................................................. 12 Method ......................................................................................................................................... 14 Study area and data sources ...................................................................................................... 14 Statistical analysis ..................................................................................................................... 16 Results .......................................................................................................................................... 19 Climate regions ......................................................................................................................... 19 Species assemblages .................................................................................................................. 19 Identification of indicator species ............................................................................................. 20 Discussion ..................................................................................................................................... 21 Species assemblages and climate conditions............................................................................. 21 Indicator species among climate regions................................................................................... 22 The use of indicator species under global change ..................................................................... 23 Acknowledgements ...................................................................................................................... 24 iv
Conclusion générale ....................................................................................... 28 Perspectives ..................................................................................................... 31 Références : ..................................................................................................... 33 Appendix 1: Climate regions clustering. ...................................................... 41 Appendix 2: General characteristics descriptions of climate regions. ...... 43 Appendix 3: Positives and negatives associations observed among the indicator species assemblages........................................................................ 47 Appendix 4: Species information and life history traits. ............................ 51 v
Liste des tableaux Table 1: Parameter estimates () and standard errors (SE) of the top-ranking logistic regression (lowest AIC, Supplementary material Appendix 2, Table A3b) predicting the probability that a bird species was an indicator, as a function of Total Links, r2 and Proportion of occupied stations, for each climate region. Total model is based on observations from all climate regions combined, with region considered as a random effect. N corresponds to the total number of species……………….…………………………………………………….38 Table 2: Jaccard index comparing the similarity in bird indicator species between climate regions. A value of 1 would indicate an identical assemblage of indicator species, and a value of 0, a complete turnover.………………………………………………………………..…38 vi
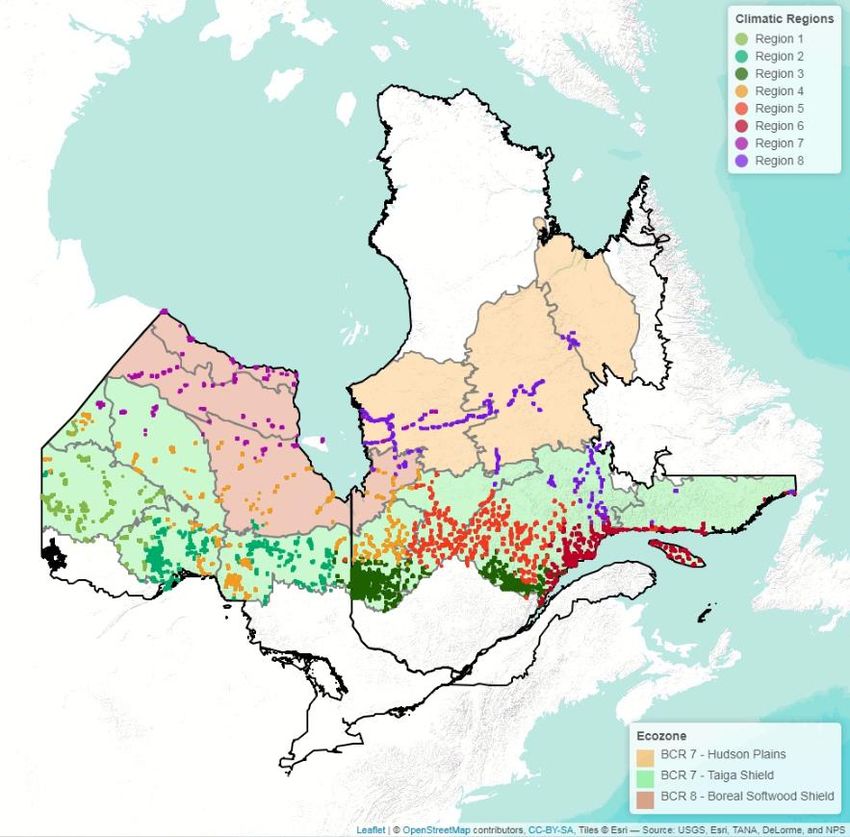
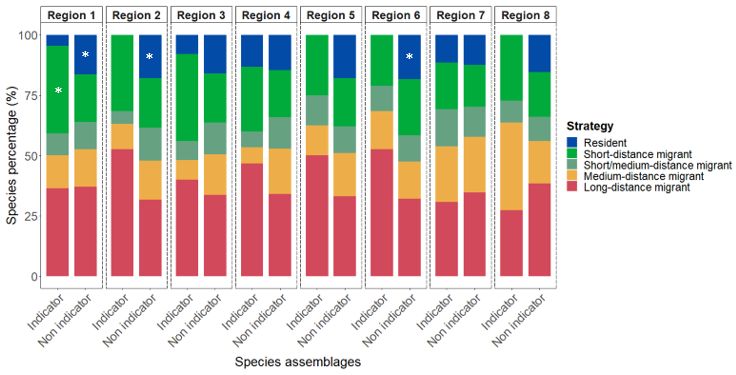
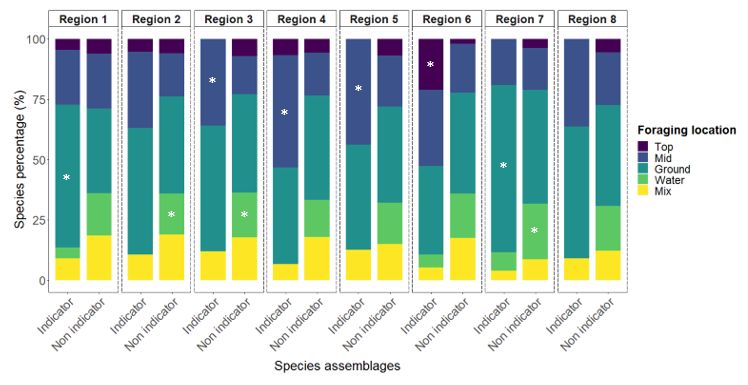
Liste des figures Figure 1: Distribution of point-count stations among climate regions represented by region- specific colored dots. The two bird conservation regions (BCR 7 and 8) are represented with light colors in the background and are further subdivided into ecoregions delimited by grey lines.………………………………………………………………………………………..36 Figure 2: Number of significant aggregations (in black) and segregations (in red) that indicator species (Supplementary material Appendix 4, Table A1 for description of species codes) have with other species in climate region 1 and 2. Species under the dotted lines are present in both climate region..……………………………………………………………..37 Figure 3: Percentage (%) of wintering strategies for all indicator and non-indicator bird species detected in each climate region. Only species recorded in 10 stations of the study area were considered. * p
« Soyez comme l'oiseau posé pour un instant sur des rameaux trop frêles qui sent plier la branche, et qui chante pourtant, sachant qu'il a des ailes » - Victor Hugo viii
Remerciements Je tiens tout d’abord à remercier l’ensemble des personnes qui ont été impliquées dans ce projet, tant sur le terrain, qu’en laboratoire et par visioconférence ; je n’aurais pu effectuer ce travail sans l’aide et le soutien de vous tous. Je remercie tout d’abord mon directeur de recherche Daniel Fortin pour m’avoir offert l’opportunité de prendre part à ce projet en milieu nordique. Merci de m’avoir fait confiance pour partir échantillonner dans le nord et faire des analyses statistiques poussées pour lesquelles je ne me sentais pas vraiment confiant. Merci également d’avoir été là pour m’orienter et me corriger lorsque je partais dans des directions un peu trop aléatoires. Je remercie également mon co-directeur de recherche Marcel Darveau pour m’avoir fait découvrir le monde fascinant des oiseaux. Merci d’avoir su montrer ta passion pour les oiseaux tout en m’aidant à interpréter certaines relations d’espèces qui m’étaient encore inconnues. Je remercie évidemment Christian Hébert, qui fut mon co-directeur de recherche tout au long de l’étude, jusqu’au changement de cap, pour des raisons administratives. Merci de m’avoir soutenu et encouragé durant le terrain sur la Côte-Nord. Merci également pour nos discussions sur le monde fascinant des insectes. Bien que j’aie préféré rédiger le mémoire sur les oiseaux, les insectes et notamment les coléoptères restent des organismes qui ne cesseront de m’intéresser. J’aimerais également remercier Frédéric Maps et Stéphane Boudreau d’avoir accepté de faire partie de mon comité d’encadrement. Vos commentaires m’ont permis de grandement améliorer ce projet du début à la fin. Merci également à Sergio Ewane Ebouele et Louis-Paul Rivest pour nos discussions traitant des aspects statistiques du projet. Les modèles nuls étaient une boîte noire au début, je les aborde avec un peu plus de confiance maintenant grâce à vous ! Merci à toutes les personnes qui ont participé au terrain sur la Côte Nord, ce fut une épreuve particulièrement difficile qui n’aurait pas été possible sans votre aide. Merci notamment à Jean-Michel Béland, Olivier Jeffrey, Anne Cotton-Gagnon, Sabrina St-Onge et Sébastien Dagnault. Merci également à Kristen Lalla et Pierre-Luc Brin pour votre résistance sans limites aux mouches et aux conditions climatiques. Je remercie également la Station Uapishka et toutes les personnes qui m’ont aidé sur place à rendre ce terrain le plus agréable possible ! Merci également à Marie-Soleil Morneau, Georges Pelletier, Ludovic Leclerc et Nicolas Bédard pour tout le travail effectué sur les insectes. Ces années auraient été bien différentes sans mes collègues de laboratoire. Merci à Marie- Caroline Prima et Ricardo Simon pour vos conseils, vos encouragements, pour ces soirées jeux pleines de suspens et pour les concerts du nouveau Jimmy Hendrix. Merci également à Virginie Vanlandeghem pour ton aide avec mes analyses cartographique et statistique, et également pour ton humeur si particulière qui te rende particulièrement attachante comme partenaire de bureau. Évidemment je ne pourrais pas oublier de remercier Guillemette ix
Labadie avec qui j’ai commencé ces études au Québec, supportrice sans faille, ton aide inestimable m’a été d’un grand secours. Je ne saurais probablement pas te remercier assez pour tout ce que tu as fait, pour avoir toujours pris le temps de m’expliquer des concepts qui m’étaient incompréhensibles, pour m’avoir supporté durant le terrain et le laboratoire, puis par visioconférence. Je souhaiterais remercier également l’ensemble des personnes que j’ai rencontré dans les associations étudiantes, autant à l’AELIÉS, l’AEGSEG ou l’ACCEBUL. J’y ai rencontré des personnes incroyables et j’ai adoré défendre certains enjeux étudiants et organiser des événements de vulgarisation scientifique avec vous tous. Merci notamment à Camille Lavoie, Charlotte Carrier-Belleau, Marie-Pier Brochu, Marianne Valcourt, Catherine Chagnon Audrey-Jade Bérubé, Mathilde Poirier, Frédéric LeTourneux et Frédéric Dulude-de-Broin. Merci également à mes amis rencontrés durant mes sorties ornithologiques et plus particulièrement la dernière année ; merci à Pierrick Bloin, Clara Casabona I Amat, Robin Besançon, Thierry Grandmont, Bruno Drolet, Andrew Coughlan, Marie O’Neill et Louise Bilodeau. Vous m’avez permis de développer une passion qui m’a fait énormément de bien en ces temps de pandémie, tout en réorientant d’une certaine façon mon avenir professionnel. Je souhaite également remercier tous les colocataires que j’ai rencontrés depuis que je suis à Québec et particulièrement Gabrielle Jolicoeur, Gabriel Piette-Lauzière et Maëliss Hoarau. Votre soutien, vos encouragements, nos soirées jeux et bouffe, particulièrement en temps de pandémie m’ont vraiment aidé et je vous en suis particulièrement reconnaissant. Merci d’avoir pris le temps de vous inquiéter sur ma santé mentale et de vous être intéressés aux oiseaux que j’avais vus sur les plaines d’Abraham. Je n’oublie pas non plus Croquette, aussi attachante qu’insupportable. Merci également à Mathilde Flahaut, Marine Queffeulou et Régis Ongaro-Carcy pour nos belles soirées et journées passées ensemble. Votre soutien et vos encouragements ont été inestimables. Merci également à tous mes amis avec qui j’ai passé des moments inoubliables et notamment Justine Faramia et Christel Blot. Je remercie également mes amis en France qui malgré la distance m’ont encouragé et plus particulièrement Ophélie Silvio, Sophia Lambert et Sara de Maio. Finalement je remercie ma famille ; les vacances passées durant ces dernières années sont inoubliables et malgré nos appels Skype parfois un peu trop espacés, je n’ai pas cessé de penser à vous pour me donner de la force de continuer. Merci d’avoir cru en moi et de m’avoir continuellement encouragé. Je vous souhaite une bonne lecture. x
Avant-propos Ce mémoire de maîtrise vise à évaluer l’utilisation d’espèces indicatrices d’assemblages dans un contexte de changement climatique. Le mémoire comporte trois parties : Une introduction générale, un chapitre présenté sous la forme d’un article scientifique rédigé en anglais et une conclusion générale. En tant que premier responsable du projet et contributeur (développement des objectifs, extraction des données, analyse des données, réalisation des simulations, écriture du mémoire et de l’article), je serai premier auteur de l’article. Les chercheurs coauteurs de l’article sont : - Daniel Fortin, directeur de maîtrise et professeur au département de biologie à l’Université Laval. Il a contribué à l’élaboration du projet, au développement de la méthode statistique, à l’interprétation des résultats et à la rédaction et la révision de l’article. - Marcel Darveau, codirecteur de maîtrise et professeur associé au département des sciences du bois et de la forêt à l’Université Laval. Il a contribué à l’élaboration du projet, à la rédaction et à la révision de l’article. - Christian Hébert, professeur associé au département des sciences du bois et de la forêt à l’Université Laval et chercheur pour le Service Canadien de la Forêt à Ressources Naturelles Canada. Il a contribué à l’élaboration du projet, à la rédaction et à la révision de l’article. - Sergio Ewane Ebouele, professionnel de recherche au département de mathématiques et de statistique à l’Université Laval. Il a contribué au développement de la méthode statistique et à l’analyse des données. - Louis-Paul Rivest, professeur au département de mathématiques et de statistique à l’Université Laval. Il a contribué au développement de la méthode statistique. Le chapitre principal servira de base à un article qui sera soumis à une revue scientifique internationale en anglais. xi
Introduction Contexte général Partout sur la planète, la biodiversité décline à un rythme sans précédent, laissant présager une sixième extinction de masse causée cette fois par l’Humain (Butchart et al. 2010, Barnosky et al. 2011). Ce déclin risque d’avoir des conséquences importantes sur le fonctionnement des écosystèmes et devrait affecter leur capacité des à fournir des services (Loreau et al. 2001, Hector et Bagchi 2007). Les espèces participent en effet de façon importante à la formation d’habitat, au cycle des nutriments et à la productivité des écosystèmes (Cardinale et al. 2012). La surveillance et la protection de cette biodiversité représentent toutefois un défi d’envergure. En effet, on estime que seulement 10 % de la diversité totale en espèces de la planète est actuellement décrite (Mora et al. 2011). De plus, il a été montré que les espèces réagissaient de diverses façons aux changements planétaires (Walther et al. 2002). Une solution pour mieux surveiller la biodiversité consiste en l’identification d’espèces indicatrices des conditions du milieu ou représentatives d’assemblages d’espèces et de groupes d’espèces associés à des milieux en particulier, sans avoir à en effectuer un inventaire exhaustif (Noss 1990, Carignan et Villard 2002). Ces espèces peuvent également, par leur présence, absence ou abondance, indiquer le stade de succession lors du processus de rétablissement des fonctions écologiques (González et al. 2013) ou agir comme des espèces focales pour permettre d’anticiper des changements critiques au sein des communautés (Lambeck 1997, Lindenmayer et Likens 2011). Agissant en synchronie avec la modification des conditions environnementales, due en grande partie aux activités humaines, les changements climatiques ont déjà des conséquences importantes sur la biodiversité (Parmesan et Yohe 2003, Root et al. 2003, Bellard et al. 2012). Avec les changements planétaires rapides auxquels nous faisons face, il est particulièrement important d’identifier les conséquences possibles du choix de certains indicateurs d’assemblages d’espèces comme outils de conservation. Dans ce contexte, le programme de recherche Sentinelle Nord de l’Université Laval finance depuis 2016 un grand nombre de projets de recherche en santé, optique, microbiologie et environnement. En appuyant la recherche transdisciplinaire et nordique, l’objectif est de mieux anticiper les conséquences des changements planétaires en milieu 1
nordique tout en permettant le développement d’outils innovants. En effet, les écosystèmes boréaux et arctiques, qui font déjà face à des changements rapides et majeurs (p. ex., réduction de l’épaisseur de glace), devraient voir ces changements s’amplifier dans les prochaines années (IPCC 2013). En considérant l’effet des changements planétaires sur la biodiversité, et plus spécifiquement sur les assemblages aviaires, la présente étude devrait permettre de fournir des informations complémentaires à l’utilisation du caribou comme espèce parapluie, pour suivre et conserver la biodiversité. Les changements climatiques et la forêt boréale Les différents scénarios climatiques produits par le Groupe d’experts Intergouvernemental sur l’Évolution du Climat (GIEC) prévoient une augmentation globale de la pluviométrie et des températures dans les prochaines décennies (IPCC 2013). Les changements climatiques affectent déjà de façon globale la biodiversité et le fonctionnement des écosystèmes, notamment en modifiant la fréquence et la sévérité des perturbations (Walther et al. 2002, Scholze et al. 2006). Ces changements climatiques devraient avoir des effets plus importants dans les latitudes nordiques avec une augmentation comprise entre 3 °C et 5 °C, ainsi qu’une légère augmentation des précipitations par rapport à la période 1980-1999, dépendamment des régions et des scénarios considérés (IPCC 2013). À l’échelle du Canada, les modèles prévoient que les précipitations devraient augmenter dans l’est, mais diminuer dans l’ouest, tandis que l’augmentation des températures se ferait de façon plus importante autour de la baie d’Hudson et au nord (Price et al. 2011). Ce changement dans la pluviométrie, associé à une augmentation de la température devrait permettre une augmentation de la productivité primaire nette et de la biomasse au nord de la forêt boréale (Boisvenue et Running 2006). Néanmoins, dans la portion sud de la forêt boréale canadienne, la diminution prévue des précipitations devrait engendrer un stress hydrique plus important, limitant ainsi la productivité des écosystèmes (Girardin et al. 2016). Cette réponse risque toutefois de varier spatialement puisque toutes les essences forestières de la forêt boréale ne réagissent pas de la même façon à ces changements climatiques. Alors même que les différentes essences de la forêt boréale devraient faire face à des conditions de croissance différentes, les régimes de perturbations naturelles, tels que les feux et les insectes, devraient également être affectés (Price et al. 2013). Ces changements 2
toucheront l’ensemble du Canada, avec une fréquence (Bergeron et Flannigan 1995), une sévérité (Stocks et al. 1998) et sur des superficies plus élevées (Jain et al. 2017). Ils devraient être particulièrement importants pour les forêts de l’est du Canada (Boulanger et al. 2018) et avoir des conséquences directes sur la structure et la composition forestière de la forêt boréale en diminuant la proportion de forêts matures. Les espèces qui dépendent d’environnements récemment brûlés durant tout ou une partie de leur cycle vital, c.-à-d., les espèces « pyrophiles », seraient alors favorisées (Weber et Flannigan 1997, Bergeron et al. 2002). Cependant, selon la région géographique étudiée, ces perturbations naturelles ne devraient pas avoir les mêmes conséquences sur les écosystèmes. Au Québec par exemple, le cycle de feu est d’environ de 270 ans dans l’ouest alors qu’il dépasse 500 ans dans l’est (Bouchard et al. 2008). Un patron semblable est également observé dans la province de l’Ontario (Bridge 2001). Une augmentation dans la fréquence des perturbations naturelles telles que le feu pourrait donc avoir des conséquences plus importantes sur certains assemblages d’espèces, moins adaptés aux perturbations récurrentes (Willig et Presley 2018). Résistance des assemblages d’espèces aux changements de la forêt boréale En raison d’une augmentation des perturbations anthropiques et naturelles, qui aura pour conséquence la modification des conditions environnementales, il est utile de se demander comment la disparition et le gain locaux d’espèces vont modifier la structure des assemblages. Le « paradoxe de le biodiversité nordique » prévoit en effet une augmentation de la richesse spécifique, autant animale que végétale, malgré la disparition de certaines espèces, du fait de la remontée d’autres présentes plus au sud (Berteaux et al. 2010). Cela n’empêchera néanmoins pas la disparition de certaines espèces plus spécialistes. La structure des assemblages d’espèces devrait donc être affectée avec des conséquences multiples sur les fonctions écologiques (Kerbiriou et al. 2009, Gallagher et al. 2013). La résistance des assemblages aux perturbations pourraient ainsi être réduite, modifiant de fait l’intégrité biologique (c.-à-d., capacité d’un écosystème à supporter et maintenir une composition en diversité d’espèces et en diversité fonctionnelle, représentative de la variabilité naturelle, Karr 1991) de la forêt boréale. Celle-ci pourrait ainsi devenir plus sensible aux activités de développement en milieu nordique (Noss 2000, Frego 2007). 3
Toutefois, ces changements spécifiques devraient se faire de façon inégale, selon les capacités des espèces à faire face à ces nouvelles conditions. On s’attend en effet à un remaniement des assemblages d’espèces, de près de 10 % par décennie (Dornelas et al. 2014). Or celui-ci devrait se faire de façon variable, dépendamment des capacités de déplacement des espèces (p. ex., direction, vitesse ; Lenoir et al. 2008) ce qui pourrait avoir des conséquences sur les interactions trophiques notamment. L’hypothèse d’assurance suggère que la biodiversité permet de protéger les écosystèmes d’une perte de fonctions écologiques, en raison de la présence de certaines espèces capables de maintenir ce fonctionnement (Yachi et Loreau 1999, García-Palacios et al. 2018). Avec l’arrivée de nouvelles espèces, le nombre de liens entre espèces pourrait considérablement augmenter dans les régions plus nordiques (Kortsch et al. 2015). Ces nouveaux liens pourraient ainsi modifier la structure des assemblages d’espèces, avec des conséquences possiblement importantes sur le fonctionnement des écosystèmes (Albouy et al. 2014). En étudiant comment les associations significatives entre espèces changent selon les conditions climatiques, nous serons davantage en mesure de prévoir les effets des changements climatiques sur les assemblages d’espèces. Dans le but de préserver l’intégrité écologique, les stratégies de conservation actuelles tentent de protéger différents types de zones écologiques, qui conjointement, permettent de préserver une proportion importante de la biodiversité (Groves et al. 2012, Stralberg et al. 2018). Toutefois, cela présente plusieurs limites. D’une part, avec des conditions climatiques changeantes de façon variable, les zones supportant une grande diversité en espèces pourraient ne plus correspondre aux réserves naturelles mises en place actuellement (Araújo et al. 2004, Heller et Zavaleta 2009). D’autre part, les changements induits par la disparition ou l’arrivée d’une espèce avec un trait d’histoire de vie nouveau dans un assemblage sont encore peu pris en compte, alors qu’ils peuvent accentuer les effets des changements climatiques sur les espèces associées (Galetti et al. 2013). Par exemple, dans une étude sur la relation entre les assemblages aviaires et certains végétaux, Galetti et al. (2013) ont montré que la disparition de certaines espèces granivores avait pour conséquence une réduction de la taille des graines observée sur quelques dizaines d’années, diminuant ainsi la régénération de certaines espèces de palmiers au Brésil. Finalement, la rapidité de ces effets peut différer en fonction du type, de la sévérité de la perturbation et de la taille de l’assemblage. Étant donné que l’abondance des espèces n’est pas uniforme (Preston 1948), il semble cependant 4
difficile d’identifier et d’anticiper ces changements sur l’ensemble des espèces. Face à l’enjeu titanesque que représente l’inventaire de l’ensemble des espèces vivantes, l’utilisation d’un nombre réduit d’espèces peut donc s’avérer prometteuse. L’utilisation d’indicateurs Dans les dernières décennies, les scientifiques ont proposé l’utilisation d’indicateurs dans le but de rapidement suivre des modifications dans la composition des communautés, les changements planétaires ou les propriétés d’un système, à différentes échelles spatio- temporelles. Ces indicateurs pouvaient être très spécifiques (p. ex., fèces de lièvres pour estimer leurs densités ; Murray et al. 2002) ou plus général (type d’humus pour déterminer le régime hydrologique d’un site ; Ponge et Chevalier 2006). Afin d’évaluer les effets de changements planétaires sur les communautés, le concept d’indicateur peut être utilisé en étant associé à de nombreux attributs (p. ex., quantité de bois mort) au sein d’un site (Carignan et Villard 2002, Azeria et al. 2009). Il peut également informer d’un changement de conditions environnementales (Fausch et al. 1990) ou de la biodiversité (Correia et al. 2017). Fleishman et al. (2005) ont ainsi montré qu’il était possible de prévoir au Nevada, États-Unis, plus de 66 % de la richesse en espèces d’oiseaux et de papillons à partir de seulement six espèces de papillons. On retrouve ainsi dans la littérature scientifique plusieurs définitions des termes « indicateur » et « espèce indicatrice », dépendamment de l’objectif de l’étude, sans qu’ils ne soient toujours bien définis (Siddig et al. 2016). Certaines espèces ou groupes d’espèces représentent des conditions biotiques et abiotiques du milieu. D’autres ont été identifiés comme plus sensibles aux changements environnementaux. D’autres espèces encore permettent d’indiquer la richesse spécifique d’une région donnée, sans avoir à effectuer d’inventaire exhaustif de cette biodiversité (Lawton et Gaston 2001). Ces espèces indicatrices, du fait de leurs présences, peuvent expliquer, et donc prévoir, une partie de la variation de richesse spécifique parmi des sites. C’est cette dernière définition d’indicateur qui sera utilisée dans cette étude. Alors que la forêt boréale devrait subir de multiples changements rapides, son échantillonnage reste difficile, pour des contraintes techniques et économiques. De fait, 5
l’utilisation d’espèces indicatrices d’assemblages peut être une alternative intéressante. Leur utilisation reste toutefois débattue, notamment à cause de la difficulté d’inférer des liens entre les espèces et les variables environnementales, rendant difficile l’interprétation écologique de la présence/absence des espèces (Lindenmayer et al. 2015). Les critiques portent également sur la prise en compte limitée de l’effet de l’hétérogénéité spatio-temporelle sur le potentiel indicateur des espèces (Paltto et al. 2006). En effet, Zettler et al. (2013) ont montré qu’une même espèce pouvait réagir de façon différente à des facteurs de stress naturels et anthropiques, selon son stade de vie, mais également selon les conditions environnementales. La répartition des espèces et la composition des assemblages d’espèces peuvent également être variables spatialement. Les interactions biotiques semblent en effet plus importantes à fines échelles spatiales, tandis qu’à plus larges échelles, ce sont les conditions climatiques et les traits d’histoire de vie qui impacteraient davantage la composition des assemblages (Zettler et al. 2013). Si certaines études mettent en évidence les variations spatio-temporelles des assemblages d'espèces associées au climat, on connaît encore peu les conséquences attendues sur les espèces indicatrices et les traits d’histoire de vie (Santillán et al. 2018). Les modèles nuls pour décrire les patrons de co-occurrence Depuis des décennies, la compréhension des mécanismes expliquant les patrons de co-occurrence d’espèces demeure un thème de recherche important en écologie des communautés. Tandis que Diamond (1975) proposait l’existence de « règles d’assemblages des communautés » basées principalement sur les interactions entre espèces pour expliquer leur coexistence, Connor et Simberloff (1979), à l’aide d’une analyse par modèle nul, soutenaient que la co-occurrence des espèces n’était pas différente de celle attendue grâce au hasard. Depuis, la recherche de règles d’assemblages des communautés se fait essentiellement à l’aide de deux types de tests : les modèles nuls par paires et les modèles probabilistes par paires (Lavender et al. 2019). Malgré une propension aux erreurs de type II (c.-à-d., faux négatifs), ces tests ont un faible taux d’erreurs de type I (c.-à-d., faux positifs) et sont donc particulièrement adaptés pour identifier les associations significatives entre espèces. Toutefois, la difficulté en écologie des communautés réside dans l’utilisation de matrice de présence-absence, contenant des milliers de paires d’espèces possibles (Gotelli et 6
Ulrich 2010). L’analyse par modèles nuls est particulièrement adaptée à ce genre de situation et elle consiste à comparer la matrice de base avec des milliers de matrices générées aléatoirement (Gotelli 2000). La façon de générer ces matrices aléatoires peut dépendre des questions écologiques, en imposant ou non des contraintes sur la randomisation des occurrences par sites et/ou par espèces. Néanmoins, il est important de considérer que chacune des méthodes choisies peut influencer le nombre d’associations négatives et positives (Azeria 2004). L’interprétation de ces associations d’espèces obtenues à partir de leur co-occurrence reste cependant débattue. En effet, un certain nombre d’articles scientifiques considèrent ces associations significatives comme représentatives d’interactions écologiques et de besoins environnementaux, tandis qu’une partie de ces associations peuvent être dues à d’autres facteurs (Blanchet et al. 2020). L’utilisation de modèles nuls permet néanmoins de réduire le nombre d’espèces ayant un potentiel d’indicateur d’assemblages d’espèces en considérant uniquement les espèces ayant des associations significatives (D’Amen et al. 2018). Cependant, l’identification d’espèces indicatrices, à partir des associations d’espèces, est souvent faite sur des petites bases de données et sans nécessairement prendre en compte les variations climatiques existantes (Azeria et al. 2009, Thorn et al. 2016). Avec le développement des sciences participatives, des données de qualité à large échelle spatiale sont maintenant de plus en plus accessibles. Les changements planétaires ayant lieu sur des superficies variables, il est donc pertinent de chercher à identifier des indicateurs à plusieurs échelles spatio-temporelles, de façon à mieux anticiper les conséquences futures. L’utilisation de la faune aviaire Les oiseaux sont parmi les taxa les plus fréquemment suivis par les programmes scientifiques professionnels et citoyens (Feldman et al. 2021). Grâce aux milliers d’ornithologues amateurs et professionnels, la science citoyenne, par exemple les atlas des oiseaux nicheurs (Cadman et al. 2007, Robert et al. 2019), permet aux chercheurs d'accéder à des ensembles de données à vaste échelle spatiale. La répartition et le mode de vie des espèces, notamment en Amérique du Nord sont donc de mieux en mieux connus, bien que les territoires les plus au nord restent encore peu échantillonnés. Ce suivi important et les connaissances générées ont permis de conclure que certaines espèces déclinent 7
dangereusement ces dernières années (DesGranges et Morneau 2010, Rosenberg et al. 2019). Les causes sont multiples (p. ex., changement d’occupation du territoire) et inégales selon les espèces. On s’attend ainsi que les changements climatiques modifient la répartition actuelle des oiseaux. Des zones identifiées comme refuges climatiques pourraient alors être trop éloignés, voire impossible d’accès pour certaines espèces (Stralberg et al. 2019). On observe ainsi depuis plusieurs décennies une arrivée des oiseaux migrateurs significativement plus hâtive dans l’année (Butler 2003), et un déplacement des espèces à des latitudes plus élevées (Møller et al. 2008). En France, par exemple, la composition des communautés d’oiseaux s’est déplacée de 91 km vers le nord, tandis que durant cette même période l’augmentation de la température aurait permis un déplacement de 273 km (Devictor et al. 2008). Parallèlement, ce changement dans les températures a permis un développement plus précoce des insectes, correspondant de moins en moins avec la date d’arrivée des oiseaux migrateurs, créant un décalage trophique et un déclin de certaines espèces d’oiseaux (Visser et Both 2005, DesGranges et Morneau 2010). En Europe par exemple la date d’éclosion des œufs de la mésange charbonnière (Parus major), est de moins en moins synchronisée avec le pic de biomasse des chenilles qui constituent leur principale source de nourriture (Visser et al. 2006). Les espèces d’oiseaux de la forêt boréale étant majoritairement migratrices, ces changements dans les conditions environnementales pourraient donc réduire leurs capacités reproductrices. En modifiant l’abondance de ces espèces, la variabilité et la redondance dans les traits d’histoire de vies observés devraient également être modifiées, suite à la sélection d’espèces migratrices de plus faibles distances ou d’espèces résidentes. Cette sélection pourrait ainsi avoir des conséquences importantes sur les assemblages d’espèces en forêt boréale. Les oiseaux sont couramment utilisés comme espèces indicatrices (Fleishman et al. 2005, Zhao et al. 2017), il semble important d’identifier les conséquences de ces changements sur leur potentiel indicateur. Objectif de l’étude L’objectif général de cette étude est d’évaluer l’utilisation d’espèces indicatrices pour prévoir la richesse en espèces sur de grandes étendues spatiales, malgré des conditions climatiques largement variables. Afin de répondre à l’objectif général, trois objectifs spécifiques ont été poursuivis : déterminer l’effet des conditions climatiques à large échelle 8
spatiale sur les assemblages d'espèces (objectif 1), évaluer si les meilleures espèces indicatrices restent les mêmes entre les régions climatiques (objectif 2) et identifier certains traits d’histoire de vie (p. ex., régime alimentaire, stratégie d’hivernage) qui font qu'une espèce a plus de chances d'être un indicateur (objectif 3). Nous avons utilisé les observations acquises sur 22 041 stations d'échantillonnage réparties sur plus de 1 750 000 km2 pour atteindre nos objectifs. Nous avons également considéré l’abondante littérature scientifique traitant de la faune aviaire pour extraire des données de traits d’histoire de vie. 9
Chapitre principal - On the efficiency of indicator species for broad-scale monitoring of bird diversity under climate change Alexandre Terrigeol1, Sergio Ewane Ebouele2, Marcel Darveau3, Christian Hébert4, Louis-Paul Rivest2, Daniel Fortin1 1Département de biologie, Université Laval, Québec, QC, Canada G1V 0A6 2 Département de mathématiques et de statistique, Université Laval, Québec, QC, Canada G1V 0A6 3 Département des sciences du bois et de la forêt, Université Laval, Québec, QC, Canada G1V 0A6, 4 Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, 1055 du P.E.P.S., PO Box 10380, Succ. Sainte-Foy, Québec, QC, Canada G1V 4C7 Résumé Diverses contraintes (p. ex., temps, financement, expertise) peuvent entraver la surveillance à large échelle qui est nécessaire à l’évaluation et l’atténuation de la perte en biodiversité causée par l'homme. La co-occurrence non aléatoire d'espèces offre l’opportunité d'estimer la richesse spécifique locale en ne suivant que quelques espèces qui sont des indicateurs d'assemblages plus planétaires. Malgré des études prometteuses à fine échelle, il demeure incertain dans quelle mesure l’utilisation de ces espèces indicatrices restent un outil efficace sur de vastes étendues spatiales. Nous avons évalué l’efficacité des espèces indicatrices à prédire de manière cohérente la richesse en espèces malgré des variations climatiques. Notre étude a impliqué 22 041 points d’écoute répartis sur 1,75 M km2 de forêt boréale en Ontario 10
et au Québec (Canada), où 197 000 oiseaux de 216 espèces ont été détectés. En utilisant une analyse par modèle nul de la co-occurrence des espèces, puis des régressions multiples, nous avons identifié le plus petit ensemble d'espèces indicatrices permettant de prévoir 70 % de la variation spatiale en richesse aviaire dans chacune de huit régions climatiques. Nos analyses montrent que les indicateurs étaient des sous-ensembles non aléatoires du pool d'espèces car il s'agissait d'espèces dont la présence expliquait un pourcentage relativement élevé de la variation en richesse spécifique au sein de la région climatique et qui partageaient un nombre important d'associations significatives de co-occurrence (positives ou négatives). Bien que seules 11 à 26 espèces indicatrices étaient nécessaires pour prévoir la richesse spécifique à l'échelle régionale, un total de 57 des 216 espèces (26 %) était requis pour faire des prévisions sur l'ensemble de l’aire d'étude. Ce grand nombre d'indicateurs reflète les variations régionales des espèces significativement associées l’une à l’autre par leur co-occurrence. Les espèces indicatrices différaient largement entre les régions climatiques, et celles qui demeuraient des indicateurs dans plusieurs régions étaient représentatives de différents assemblages d'espèces. Nos observations soulèvent donc un doute sur l'utilisation des espèces indicatrices comme outil permettant le suivi de la biodiversité sous des conditions climatiques changeantes. Nous inférons de notre étude spatiale qu’avec le changement climatique en cours, les espèces actuellement identifiées comme indicateurs de richesse pourraient ne plus représenter les mêmes assemblages d’espèces dans un avenir proche. L'utilisation d’indicateurs à large échelle spatio-temporelle s'accompagne donc du défi de devoir déterminer dans quelles nouvelles conditions un ensemble donné d'indicateurs doit être remplacé par un autre, et par lequel. Abstract Various constraints (e.g., time, funding, expertise) can impede the broad-scale monitoring needed to assess and mitigate human-induced loss of biodiversity. Non-random species co- occurrence provides an opportunity to estimate local species richness by tracking only a few species that are indicators of more global assemblages. Despite promising fine-scale studies, it remains unclear whether such indicator species remain effective over broad spatial extents. We assessed the value of indicator species to consistently predict species richness despite variation in climate conditions. Our study involves 22,041 point-count stations distributed 11
over 1.75 M km2 of boreal forest in Ontario and Québec (Canada), where 197,000 birds from 216 species were detected. Using null model analysis of species co-occurrence followed by regression analysis, we identified the minimum set of indicator species that can predict 70% of spatial variation in local bird richness in each of eight climate regions. We found that indicators were non-random subsets of the species pool, as they were species whose presence explained a relatively high percentage of variations in species richness within the climate region, and those sharing a relatively high number of significant pairwise associations (positive or negative). Although only 11 to 26 indicator species were needed to predict species richness at the regional scale, a total of 57 of the 216 species (26%) were necessary to make predictions over the entire study area. This large number of indicators reflects regional variations in the species involved in significant pairwise associations. Indicator species largely differed among climate regions, and those that remained indicators in several regions were representative of different species assemblages. Our observations thus cast doubts on the use of indicator species as a tool for biodiversity monitoring under changing climate conditions. We infer from our spatial study that, with the ongoing climate change, species currently identified as indicator of species richness may no longer indicate the same assemblages in the near future. Broad-scale use of indicators thus comes with the challenge of having to determine under which new conditions a given set of indicators must be replaced by another, and by which one. Introduction The worldwide decline in biodiversity (Pereira et al. 2010, Johnson et al. 2017) has led many governments to commit to taking actions expected to reduce species extinction rate (e.g., CBD 2016). A necessary step for assessing the effectiveness of action plans is the broad-scale monitoring of biodiversity. The frequent lack of expertise required to identify species of many taxa (Kim and Byrne 2006, Troudet et al. 2017) and time and funding limitations to conduct extensive surveys have stimulated the search for simple – yet effective – biodiversity monitoring techniques (Stevenson et al. 2020). The use of indicator species has been proposed to track changes in species assemblages at various scales (Dufrêne and Legendre 1997, Heink and Kowarik 2010). In this context, indicator species are a set of species with complementary co-occurrence patterns that, collectively, can predict total 12
Vous pouvez aussi lire

















































