Lutte intégrée contre Varroa destructor : comparaison des comportements hygiéniques de l'abeille domestique Apis mellifera - Mémoire Marie-Lou ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Lutte intégrée contre Varroa destructor : comparaison
des comportements hygiéniques de l’abeille domestique
Apis mellifera
Mémoire
Marie-Lou Morin
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Marie-Lou Morin, 2022
Lutte intégrée contre Varroa destructor :
comparaison des comportements hygiéniques de
l’abeille domestique Apis mellifera
Mémoire
Marie-Lou Morin
Sous la direction de :
Pierre Giovenazzo, directeur de recherche
Nicolas Derome, codirecteur de recherche
Résumé
La sélection génétique d’abeilles mellifères (Apis mellifera) offre à l’industrie apicole la
possibilité de combattre le parasite Varroa destructor et ainsi réduire la dépendance envers
les acaricides. En effet, certaines populations d’abeilles possèdent des traits
comportementaux hygiéniques héritables qui leur permettent de résister au Varroa.
Cependant, les liens entre ces différents traits comportementaux ne sont pas encore bien
définis, ce qui limite le progrès génétique. Dans ce contexte, nous avons mesuré les traits
comportementaux de résistance au Varroa suivants : les tests de mort du couvain par
congélation (freeze-kill brood assay, FKB) et de l’aiguille (pin-kill brood assay, PKB), le
varroa-sensitive hygiene (VSH), le retrait des pupes, le mite non-reproduction (MNR) et la
réoperculation des cellules de couvain. Ces mesures ont été utilisées pour calculer les
corrélations et deux d’entre elles étaient négatives et significatives: (1) entre la réoperculation
des cellules infestées par le Varroa et la réoperculation totale et (2) entre la réoperculation
des cellules infestées par varroa et le varroa-sensitive hygiene (VSH). Nous avons également
sélectionné le meilleur modèle prédictif des niveaux d'infestation par le varroa dans les
colonies en utilisant l'approche « step-wise » basée sur le critère AIC. Notre modèle révèle
que le MNR et le FKB sont significativement liés à la population de varroas avec une relation
négative, tandis que la réoperculation était significativement liée à des niveaux d'infestation
de varroas avec une relation positive. Ainsi, les scores MNR ou FKB plus élevés sont
significativement reliés à de faibles niveaux d'infestation d'acariens dans les colonies à la fin
août, tandis qu'une activité de réoperculation des cellules de couvain plus élevée est
significativement reliée à un niveau d'infestation élevé d'acariens dans les colonies. Notre
travail indique que le comportement de réoperculation pourrait être un trait de caractère utile
pour aider la sélection de lignées d’abeilles résistante à Varroa mais que d’autres travaux
doivent être faits pour mieux comprendre sa signification.
ii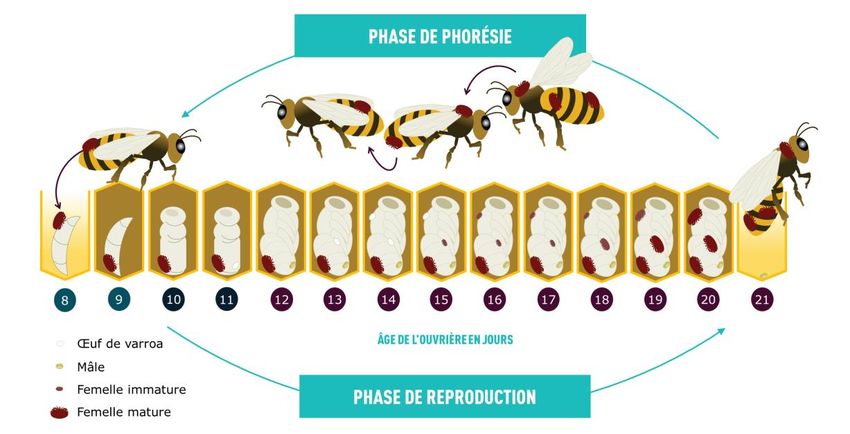
Abstract
The genetic selection of honeybees (Apis mellifera) offers the beekeeping industry the
possibility of combating the Varroa destructor parasite and thus reducing its dependence on
acaricides. Indeed, some bee populations have heritable hygienic behavioural traits that allow
them to resist Varroa infestation. However, the links between these different behavioural
traits are not yet well defined, which limits genetic progress. In this context, we measured
the following behavioural Varroa resistance traits: freeze-kill brood assay (FKB) and pin-kill
brood assay (PKB), varroa-sensitive hygiene (VSH), pupae removal, mite non-reproduction
(MNR) and recapping activity. Correlations between these measures show that two are
negative and significant: (1) between the recapping of cells infested with Varroa and the total
number of recapped cells and (2) between the recapping of cells infested with Varroa and
varroa-sensitive hygiene (VSH). We also selected the best predictive model of varroa
infestation levels using the “step-wise” approach based on the AIC criterion. Our model
revealed that MNR and FKB were significantly related to the varroa population levels with
a negative relationship, while recapping was significantly related to mite infestation levels
with a positive relationship. Thus, a higher MNR or FKB score is significantly related to low
levels of mite infestation in colonies at the end of August, while a higher recapping activity
is significantly related to a high level of mite infestation. Our work indicates that recapping
behavior could be a useful trait to aid the selection of Varroa-resistant bee lines but that
further work needs to be done to better understand its significance.
iii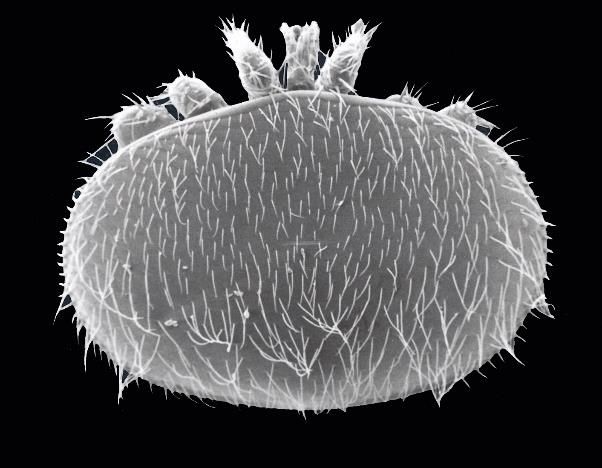
Table des matières
Résumé ................................................................................................................................... ii
Abstract .................................................................................................................................. iii
Table des matières ................................................................................................................. iv
Liste des tableaux .................................................................................................................. vi
Liste des figures .................................................................................................................... vii
Liste des abréviations, sigles, acronymes ............................................................................ viii
Remerciements ....................................................................................................................... x
Avant-propos ........................................................................................................................ xii
Introduction générale .............................................................................................................. 1
Chapitre 1 – État des connaissances ....................................................................................... 3
1.1 L’abeille domestique................................................................................................ 3
1.1.1 L’abeille domestique au Canada....................................................................... 3
1.1.2 Développement de l’Abeille et reproduction.................................................... 4
1.1.3 Dynamique annuelle de la population d’une colonie ....................................... 5
1.1.4 L’Abeille dans son environment....................................................................... 6
1.2 Varroa destructor : biologie et impacts et contrôle ................................................. 8
1.2.1 Cycle de reproduction intimement lié à celui de son hôte ................................ 8
1.2.2 Relation parasitaire en déséquilibre ................................................................ 10
1.2.3 La varroose ..................................................................................................... 11
1.2.4 Contrôle du Varroa par la lutte intégrée ......................................................... 13
1.3 La sélection génétique des comportements hygiéniques procurant la résistance à
Varroa ............................................................................................................................... 15
1.3.1. Comportements hygiéniques : mécanisme général ............................................ 17
1.3.2. Les traits impliqués dans le mécanisme de résistance au Varroa ....................... 18
1.4 Objectifs et hypothèses .......................................................................................... 19
1.4.1. Objectifs ............................................................................................................. 19
1.4.2. Hypothèses ......................................................................................................... 20
Chapitre 2 – Varroa destructor infestation level can be predicted by brood cell recapping
rate and mite non reproduction in honeybee colonies (Apis mellifera L.) ........................... 22
2.1 Résumé........................................................................................................................ 23
2.2 Abstract ....................................................................................................................... 24
iv
2.3 Introduction ................................................................................................................. 25
2.4 Materials and methods ................................................................................................ 29
2.4.1 Colony establishment and management .............................................................. 29
2.4.2 Hygienic testing: freeze-kill brood assay & pin-kill brood assay ....................... 29
2.4.3 Varroa-sensitive hygiene, removal of pupae, mite non-reproduction and
recapping activity ......................................................................................................... 30
2.4.4 Varroa infestation rate in colonies ....................................................................... 32
2.4.5 Statistical analyses ............................................................................................... 32
2.5 Results ......................................................................................................................... 34
Colony-level phenotypes .............................................................................................. 34
Predicting mite infestation level ................................................................................... 36
2.6 Discussion ................................................................................................................... 40
2.8 References ................................................................................................................... 44
Conclusion générale ............................................................................................................. 51
Retour sur les hypothèses et prédictions....................................................................... 51
Difficultés rencontrées et pistes d’amélioration ........................................................... 53
Perspectives de recherche futures ................................................................................. 54
Bibliographie ........................................................................................................................ 56
v
Liste des tableaux
Tableau 2.1: Mean, standard error (SE), minimum (Min), maximum (Max) for each
variable measured in tested colonies (N= 34 colonies). See Materials and
methods section for description of variables.
Tableau 2.2: Estimates, standard errors (SE), z-values, p-values of fixed-effect variables in
a general linear model of mixed effects for Poisson regression (glmer).
Random effect = Apiary, offset variable = log of the total honeybees in
samples for varroa counts, n = 34 colonies.
viListe des figures
Figure 1.1: Varroa destructor, parasite d'Apis mellifera, en vue dorsale (gauche) et
ventrale (droite). Les structures nommées dans le texte sont visibles : bouclier
dorsal et ventral, les pattes et apothèles ainsi que les chélicères. Photos prises
par microscope par balayage électronique par Cayambe (Licence Creative
Commons).
Figure 1.2: Schéma représentant la phase phorétique et la phase de reproduction de
Varroa destructor en parallèle avec le développement des pupes d'Apis
mellifera. (MAPAQ, Varroa destructor, mieux comprendre l’ennemi)
Figure 1.3: Photo d’Apis mellifera sur un cadre de couvain dans une colonie fortement
infestée par Varroa. L'ouvrière encerclée présente des signes cliniques
d'infection au Virus des ailes déformées (ailes atrophiées). À la droite de cette
dernière, on peut apercevoir un varroa en phase phorétique sur le thorax d’une
ouvrière. Photo par Marie-Lou Morin (2019).
Figure 1.4: Photo d'un cadre de couvain operculé lors d'un test hygiénique avec la
technique du couvain mort par congélation (freeze-kill brood assay). Photo
prise par Marie-Lou Morin (2019).
Figure 1.5: Photo d'un cadre de couvain operculé lors d’un test hygiénique utilisant la
technique du test de l’aiguille (pin-kill brood test). Photo prise par Marie-Lou
Morin (2019).
Figure 2.1: Modified frames used for varroa mite infestation: A- Langstroth frame with
foundation cut in half. B- Langstroth frame taken apart, foundation cut in half.
C- Half of foundation mounted in frame. (Photos by S. Rouleau Breton).
Figure 2.2: Spearman’s correlation matrix. Correlation coefficients (reported as ρ values)
are shown for each pair-wise comparison. Statistically significant estimates (p
< 0.05) are marked with an asterisk. Shaded blue cells represent positive
correlations and red cells represent negative correlations. Darker hues indicate
stronger correlations as indicated by the correlation color gradient. All
variables shown are also listed in Table 2.1.
Figure 2.3: Relationship between MNR and mite infestation level in colonies on August
14. Prediction model is represented by a full line, 95% confidence intervals by
dotted lines and original data of each colony by points.
Figure 2.4: Relationship between total recapping activity and mite infestation level in
colonies on August 14. Prediction model is represented by a full line, 95%
confidence intervals by dotted lines and original data of each colony by points.
Figure 2.5: Relationship between freeze-kill brood test (FKB) and mite infestation level
in colonies on August 14. Prediction model is represented by a full line, 95%
confidence intervals by dotted lines and original data of each colony by points.
viiListe des abréviations, sigles, acronymes
AIC : Critère d’information d’Akaike
CAPA : Canadian apiculturists professionnals association
CRSAD : Centre de recherche en sciences animales de Deschambault
FKB : Freeze-killed brood (assay)
MAPAQ : Ministère de l’Agriculture, des Pêches et de l’Alimentation du Québec
MNR : Mite non-reproduction
SE : Standard error
SMR : Suppressed mite reproduction
PKB : Pin-killed brood (assay)
VSH : Varroa-sensitive hygiene
viiiÀ Roland Morin, qui nous a quittés avant de
pouvoir lire le fruit du travail au cours
duquel il m’a beaucoup encouragée
ixRemerciements
Je tiens d’abord à remercier mon directeur de maîtrise Pierre Giovenazzo, pour m’avoir
donné l’opportunité de travailler avec lui sur les abeilles. Depuis notre rencontre initiale en
2016 pour initiation à la recherche, j’ai grandement apprécié ton enthousiasme, tes conseils
et toutes les idées que tu proposes. C’est grâce à toi si j’ai pu découvrir cet univers
passionnant qu’est la recherche appliquée en apiculture. Merci de m’avoir confié ce projet
stimulant, qui m’a permis de vivre des expériences si formatrices. Sans ton support et tes
encouragements, je ne sais pas si j’aurais réussi à me rendre au bout de cette aventure avec
les années pandémie qu’on a vécu, et toutes les épreuves qu’elles ont amenées.
Merci à Nicolas Derome d’avoir accepté de devenir mon co-directeur et à Nadia Aubin-Horth
de faire partie de mon comité d’évaluation. Votre accompagnement et vos suggestions à des
moments charnières de mon parcours m’ont été très précieux.
Merci aux organismes subventionnaires, le ministère de l’Agriculture, de l’alimentation et
des affaires rurales de l’Ontario (OMAFRA) et au Centre de recherche en sciences animales
de Deschambault (CRSAD), sans qui le projet n’aurait pas pu avoir lieu.
Un merci particulier à mes collègues étudiantes : Mireille Levesque pour m’avoir suivie
presque tous les jours, tu as été mon bras droit et ton aide m’a été indispensable, et Ségolène
Maucourt pour sa rigueur exemplaire, t’avoir à mes côtés m’a mis en confiance et je t’admire!
Merci infiniment à toute l’équipe CRSAD, Andrée Rousseau, Georges Martin, Mickaël
Benoit, Marc-André Corriveau, Martine Bernier et Émile Houle, de m’avoir tout appris et
m’avoir accompagnée dans la partie terrain. Un merci particulier à Marilène Paillard, qui a
été à mes côtés tout au long des évaluations dans le rucher infesté. Ta compétence et ta bonne
humeur ont rendu cette tâche agréable, ce qui est un exploit en soi. Merci également à
Stéphanie Rouleau-Breton et Matthieu Guichard de m’avoir permis de me familiariser avec
les tests avant de les appliquer dans mon projet.
Merci également à Kelsey Duscharm et Les Eccles de l’équipe Tech Transfer for apiculture
en Ontario, pour votre accueil et votre aide, collaborer avec vous fût un réel plaisir.
xPour l’aide avec les analyses statistiques, je dois remercier Gaétan Daigle pour sa
disponibilité et son expertise. Merci également à Karen Grilis pour les corrections de l’article
en anglais.
Merci au centre d’aide aux étudiants de l’Université Laval pour la documentation et le service
d’accompagnement individuel, qui a été comme un phare dans la tempête.
Pour finir, je tiens à remercier les personnes proches de moi qui m’ont soutenu tout au long
de mon parcours. Merci à ma très chère amie Rebecca, pour ton écoute, ta générosité et de
m’avoir fait décrocher avec nos sessions de Stardew Valley. Merci à ma famille,
particulièrement ma mère Emmanuella pour ton support inconditionnel, malgré la distance.
Merci à Johnny, mon compagnon de vie, pour tout.
xiAvant-propos
Ce manuscrit a été rédigé sous la forme de mémoire par insertion d’article, ce qui signifie
que l’un des deux chapitres qui forment le cœur de ce manuscrit consiste en un article
scientifique rédigé en anglais et qui sera soumis à un journal scientifique. Le corps du
mémoire comporte également une introduction générale, un chapitre de revue de littérature
rédigée en français ainsi qu’une conclusion générale, revenant sur l’ensemble des résultats et
des observations qui découlent des chapitres 1 et 2.
Le chapitre 2, intitulé « Varroa destructor infestation level can be predicted by recapping
rate and mite non reproduction (MNR) in honeybee colonies (Apis mellifera)» sera soumis
au journal MDPI Agriculture après le dépôt final de ce mémoire. L’auteure principale a fait
l’ébauche originale de ce manuscrit, de même que la récolte et l’analyse des données. La
conceptualisation, la méthodologie et l’administration du projet ont été réalisées par l’auteure
principale avec la collaboration de Pierre Giovenazzo, Ph. D., professeur-chercheur au
département de biologie et titulaire de la Chaire de leadership en enseignement en sciences
apicoles, Université Laval. La validation, la révision et l’édition du manuscrit ont été
réalisées par l’auteure principale avec la collaboration Pierre Giovenazzo. Le projet a
entièrement été supervisé par Pierre Giovenazzo et ce dernier a assuré l’acquisition du
financement.
xiiIntroduction générale
L’importance économique de l’Abeille mellifère (Apis mellifera) dans le monde est
indéniable. Bien que l’espèce soit originaire des continents africain et européen, son élevage
est essentiel pour le rendement de nombreuses cultures ici au Canada, comme le canola, la
canneberge et le bleuet. C’est pourquoi il est primordial pour les apiculteurs d’ici de pouvoir
élever des colonies d’abeilles populeuses et en santé. En effet, les abeilles doivent tolérer
l’hiver long et rigoureux de notre climat nordique, en plus de pouvoir survire en présence de
nombreuses maladies et parasites.
Un parasite en particulier attire beaucoup d’attention puisqu’il cause des dommages mortels
depuis quelques décennies, et ce, pratiquement partout dans le monde. Il s’agit de Varroa
destructor, un ectoparasite obligatoire de l’Abeille domestique, A. mellifera. L’adulte se
nourrit des corps gras et de l’hémolymphe des larves et des ouvrières adultes, et est par le
fait même un vecteur de nombreux virus potentiellement mortels. Il accomplit son cycle de
reproduction à même le couvain d’abeilles. La femelle varroa entre dans une alvéole juste
avant l’operculation de la larve. Pendant le développement de la pupe, le Varroa pondra ses
œufs et sa progéniture viendra à maturité. Ainsi, la population de varroas dans une colonie
non traitée augmente de manière exponentielle du printemps à la fin de l’été, et atteint
généralement un seuil critique à l’automne, lorsque la population d’abeilles commence à
diminuer avant l’hiver. À ce stade d’infestation, il est souvent trop tard pour empêcher des
dommages à la colonie. Malgré un niveau variable d’immunité sociale au sein de l’espèce A.
mellifera, les abeilles sont dépendantes des traitements acaricides administrés par les
apiculteurs. Sans traitement efficace, la colonie est vouée à mourir au bout de 2 à 3 ans.
Des solutions alternatives aux acaricides chimiques sont recherchées, et la sélection
artificielle de traits héritables avantageux est considérée comme une avenue intéressante par
de nombreux chercheurs. En effet, en étudiant des colonies d’abeilles sauvages qui
survivaient avec le Varroa sans intervention humaine, il a été découvert que celles-ci
exhibaient des comportements dits hygiéniques, qui procuraient la résistance.
1Les comportements hygiéniques font partie de l’immunité sociale des insectes eusociaux, et
consistent en la capacité d’une colonie à percevoir les molécules odorantes émises par du
couvain mort ou malade et de retirer ce dernier rapidement de la colonie afin de prévenir la
propagation des agents pathogènes au couvain sain. Ces comportements sont complexes et
difficiles à étudier à même la ruche, car les différentes étapes du comportement peuvent être
effectuées par des ouvrières différentes, et son expression peut être influencée par différents
facteurs environnementaux, comme l’abondance de nectar dans l’environnement. Cette
complexité fait en sorte qu’on tente toujours de bien définir les mécanismes de l’immunité
sociale, afin d’élaborer des moyens pour les évaluer fidèlement. Avec des tests
phénotypiques ou des outils génomiques et transcriptomiques mesurant les traits de
résistance avec précision, il sera plus facile de sélectionner les colonies les plus hygiéniques
et d’éventuellement distribuer massivement ces nouvelles lignées. Le but final serait que les
apiculteurs puissent avoir des abeilles capables de coexister avec le Varroa sans dépendre
des traitements annuels et diminuer les mortalités.
C’est dans ce contexte que s’est inscrit mon projet de maîtrise, au cours duquel j’ai tenté
d’approfondir les connaissances des relations entre différents traits de comportements
hygiéniques, certains étant spécifiquement associés à la lutte contre V. destructor.
2Chapitre 1 – État des connaissances
1.1 L’abeille domestique
L’Abeille mellifère (Apis mellifera) est un insecte de l’ordre des Hyménoptères, et est l’une
des neuf espèces constituant le genre Apis. Sa répartition naturelle s’étend aux continents
africain et européen, ainsi qu’au Moyen-Orient (Gould et Gould, 1988). D’autres espèces du
même genre (A. cerana, A. dorsata et A. florea) étaient jusqu’à tout récemment séparées
géographiquement d’A. mellifera et vivaient donc en allopatrie avec cette dernière (Oldroyd
et al., 1999). Aujourd’hui, A. mellifera est présente sur tous les continents, excepté
l’Antarctique. Les services de pollinisation rendus par l’Abeille mellifère engendrent
d’importantes retombées économiques, en plus de sa production de nombreux produits de la
ruche tels le miel, la cire, la propolis et la gelée royale.
1.1.1 L’abeille domestique au Canada
L’importance de l’abeille domestique au Canada est loin d’être négligeable. En effet, on
comptait en 2019 un total de 773 182 colonies gérées par 10 344 apiculteurs dans tout le pays
(Agriculture et Agroalimentaire Canada, 2020). Bien que les revenus engendrés par la vente
de miel soient importants (197 M$ par année), ce sont les services de pollinisation des
cultures comme le canola, la canneberge ou les bleuets qui font de l’abeille un outil important
pour l’industrie agricole canadienne. Pollinisant plus de 90 % de ces cultures, la valeur de ce
service est estimée à près de 2 milliards de dollars par année en rendements.
Bien que le nombre de colonies totales augmente chaque année, les apiculteurs enregistrent
annuellement des pertes qui varient entre 15-35 % depuis une quinzaine d’années (RAIZO,
2019). Ces pertes, qui surviennent durant l’hiver, augmentent drastiquement les coûts
d’exploitation de l’élevage, car le cheptel doit être reconstruit, soit par l’achat de « paquets
d’abeilles » avec une reine ou par la division des colonies restantes et l’ajout d’une jeune
reine (aussi appelé nucléi). Les causes de cette mortalité accrue sont multiples. En sondant
les apiculteurs en 2019, la Canadian Association of Professional Apiculturists a identifié
notamment le manque de nourriture, les mauvaises conditions météorologiques, une faible
3viabilité ou fertilité des reines ainsi qu’un contrôle inefficace du varroa comme étant les
causes probables les plus fréquentes.
1.1.2 Développement de l’Abeille et reproduction
Le développement de l’Abeille, traversé d’une métamorphose complète (Insecte
holométabole), passe par quatre états : l’œuf, la larve, la nymphe (pupe) et l’adulte. Pendant
les trois premiers stades de vie, rassemblés sous le terme « couvain », l’abeille se développe
dans une alvéole et dépend des ouvrières adultes pour se nourrir. Tout juste avant que la larve
entre en métamorphose, les ouvrières vont apposer un opercule de cire poreux afin de sceller
l’alvéole tout en permettant les échanges gazeux pour la respiration. La larve tisse un cocon
et se transforme en nymphe, puis émerge en adulte à la fin de son développement (Caron et
Connor, 2013). Le cycle prend 21 jours pour se compléter chez l’ouvrière, et varie entre la
reine et le faux-bourdon (16 et 24 jours respectivement).
Comme plusieurs autres espèces de fourmis ainsi que d’autres hyménoptères, A. mellifera est
une espèce eusociale (Crespi et Douglas, 1995). Ce caractère se définit par un système de
castes, la cohabitation des générations et le partage des tâches et des soins aux juvéniles par
les différents individus de la colonie. En effet, tous dépendent les uns des autres pour leur
survie et leur développement, c’est pourquoi on qualifie la colonie de superorganisme
(Seeley, 1989). La colonie est toujours composée de trois castes : une reine, de milliers
d’ouvrières et des faux-bourdons en quantité variable au cours de l’année (Caron et Connor,
2013).
La reine est le pilier de la colonie, puisqu’elle assure la pérennité de sa colonie en donnant
naissance aux nouveaux individus. En effet, cela est nécessaire puisque contrairement à la
reine qui vit en moyenne 3 ans, les ouvrières vivent en moyenne de 15 à 38 jours seulement
en été (Caron et Connor, 2013). De plus, elle émet constamment des phéromones qui assurent
la cohésion de la colonie (Slessor et al., 2005). L’un des effets de ces composés chimiques
sur le comportement des ouvrières est d’assurer la formation de la cour qui nettoie et nourrit
la reine. En plus des soins à la reine, les ouvrières effectuent toutes les tâches liées au
nettoyage du nid, au nourrissage du couvain, à la construction des rayons de cire et à la
4collecte de nourriture (nectar et pollen). Elles sont aussi les gardiennes de la ruche et
communiquent à leurs sœurs l’emplacement des ressources avec des danses élaborées. La
ségrégation des tâches se fait selon l’âge de l’ouvrière, qui débute près du couvain et termine
sa vie à l’extérieur de la ruche. Finalement, les faux-bourdons constituent la troisième caste.
Ce sont les mâles de cette espèce dont le rôle est de féconder des reines vierges lors des vols
nuptiaux des jeunes reines.
La différenciation mâle-femelle est haplodiploïde et se fait au moment de la ponte de l’œuf
par la reine. Si l’œuf est fécondé par un spermatozoïde, l’individu sera une femelle
(diploïdie). S’il ne l’est pas, ce sera un mâle (haploïdie). La différenciation entre une ouvrière
et une reine est d’origine épigénétique et vient d’un facteur nutritionnel, la dernière étant
nourrie plus abondamment et avec plus de gelée royale (Winston, 1987). Cette alimentation
agit sur l’expression génétique et lui permettra de développer complètement son système
reproducteur, tandis que celui des ouvrières demeure atrophié. La reine effectue jusqu’à trois
vols nuptiaux quelques jours après sa naissance. Lors de ces vols, elle sera fécondée par une
douzaine de faux-bourdons et entreposera cette réserve de sperme dans un organe spécialisé,
la spermathèque (Winston, 1987).
Comme la reine est diploïde et qu’elle féconde ses œufs avec le bagage génétique de plusieurs
mâles différents, les ouvrières d’une colonie n’ont pas toutes le même lien de parenté. En
effet, les filles issues du même père seront des super-sœurs et auront un lien de parenté très
grand (75%), tandis que celles ayant des pères frères seront des sœurs (50% lien de parenté)
et celles ayant des pères non apparentés seront des demi-sœurs (25% lien de parenté).
1.1.3 Dynamique annuelle de la population d’une colonie
La population d’abeilles varie grandement selon la saison, surtout dans les régions tempérées
comme au Canada. Elle diminue rapidement à l’automne lorsque les températures extérieures
sont plus froides et que les ressources se font rares, et continue de diminuer pendant l’hiver,
puisque la reine ralentie graduellement sa ponte avant de l’arrêter complètement en à la fin
de l’automne (Melathopoulos, 2007). Cela permet aux abeilles de diminuer leur métabolisme
pour maintenir le nid à une température d’au moins 15°C, alors que le couvain requiert d’être
5maintenu à 33°C (Melathopoulos, 2007). Les abeilles sont confinées au sein de la ruche et
doivent survivre avec les réserves de miel accumulées pendant la saison, d’où l’importance
de réduire leur métabolisme. La ponte reprend néanmoins lentement après le solstice d’hiver,
afin de remplacer les abeilles mortes au cours de l’hiver (Seeley et Visscher, 1985). Afin de
garder la ruche à température convenable pour assurer la survie de la colonie, les ouvrières
vont former une grappe dense et agiter leurs muscles thoraciques afin de produire de la
chaleur. La grappe d’abeilles se resserre et prend de l’expansion proportionnellement avec la
diminution ou l’augmentation de la température extérieure. Au printemps, avec l’arrivée des
ressources florales, la population dans la colonie augmente rapidement grâce à l’activité de
la reine et se maintient pendant l’été. Pendant cette période, la reine peut pondre jusqu’à 2000
œufs par jour. Une production importante de couvain en début de saison est un gage de bon
développement de la colonie pour le reste de l’été et donc de meilleures récoltes de miel
(Szabo et Lefkovitch, 1989). C’est également important pour la pollinisation des grandes
cultures québécoises (pommes, bleuets, canneberges), dont les besoins sont très tôt au
printemps. La population atteint un maximum au mois de juin, et se maintient pour le reste
de l’été.
1.1.4 L’Abeille dans son environment
La colonie d’abeilles évolue dans un nid à l’abri des intempéries. En apiculture nord-
américaine, on élève l’abeille domestique A. mellifera dans des ruches standardisées afin de
faciliter l’inspection et l’entretien du nid, ainsi que l’extraction du miel. Une ruche est donc
une boite remplie de cadres rectangulaires sur lesquels les abeilles vont construire les rayons
de cires. Ces rayons sont constitués d’alvéoles, des prismes à base hexagonale dans lesquels
seront entreposés le miel et le pollen, mais serviront aussi de cellules pour élever le couvain.
En plus d’évoluer au sein de son nid, la colonie dépend de l’environnement extérieur pour
s’alimenter. D’abord, les abeilles vont chercher leur source d’énergie dans le nectar des fleurs
disponibles. Les butineuses parcourent des distances variables selon l’abondance des
ressources et la météo, et 95% des vols se font à l’intérieur d’un rayon de 6 km autour de la
ruche (Visscher et Seely, 1982). Les butineuses rapportent aussi du pollen qu’elles amassent
en pelotes sur les corbiculae, structures situées sur leurs deux pattes postérieures (Parker,
61926). Le pollen fournit les acides aminés nécessaires au développement du couvain, et peut
lui aussi être entreposé dans les alvéoles, sous forme de pain de pollen. Dépendamment de
l’espèce florale dont le pollen est issu, ce dernier aura une composition en acides aminés, une
couleur et une forme de grain unique. Par exemple, le pollen du pissenlit (Taraxacum
officinaIe) manque sévèrement de valine et d’isoleucine, deux des dix acides aminés
essentiels aux abeilles (Loper et Cohen, 1987). Donc, afin d’aller chercher en quantité
suffisante tous les acides aminés essentiels, plus d’une espèce florale est nécessaire. De plus,
la période de floraison des végétaux est limitée à quelques jours ou quelques semaines selon
l’espèce. Ces deux facteurs expliquent pourquoi les abeilles ont besoin d’une variété et d’une
abondance de fleurs dans leur environnement pour combler leurs besoins nutritionnels tout
au long de la saison apicole.
En plus de la nourriture, les butineuses vont rapporter de la propolis et de l’eau au nid. Les
abeilles utilisent l’eau pour la thermorégulation à l’intérieur de la ruche. La propolis, quant à
elle, est une substance résineuse produite par certains arbres. Les abeilles s’en servent comme
antiseptique et pour tapisser l’intérieur de la ruche ou colmater les brèches. Les effets
antimicrobiens de la propolis aideraient les abeilles à prévenir l’entrée de certains agents
pathogènes dans la colonie. En effet, la proximité entre les individus rend la colonie
susceptible aux infections et aux parasites, puisque la transmission est facile entre les
individus, notamment en raison de la trophallaxie. L’utilisation de la propolis comme
antiseptique, avec le nettoyage du nid, la sécrétion de composés antimicrobiens dans la gelée
royale (Fujiwara et al., 1990), le toilettage (grooming) (Peng et al., 1987) et la production de
venin par les ouvrières (Baracchi et al., 2011), font partie de l’immunité sociale de la colonie.
En effet, tel que décrit par Cremer et al., 2007), l’immunité sociale est l’ensemble des moyens
employés par les insectes sociaux pour maintenir la santé de la colonie. Certaines abeilles
présentent des comportements dits « hygiéniques », ce qui signifie qu’elles retirent
rapidement le couvain mort ou malade de la colonie afin de limiter la propagation des
pathogènes aux individus sains. Le sujet sera abordé davantage à la section 1.3.
71.2 Varroa destructor : biologie et impacts et contrôle
Varroa destructor (Anderson et Trueman, 2000)
B est un acarien ectoparasite d’A. mellifera,
A
et représente la plus grande menace en apiculture (Sammataro et al., 2000 ; Rosenkranz et
al., 2010 ; Nazzi et Le Conte, 2016). L’anatomie et la physiologie des varroas sont bien
adaptées à leur hôte (Figure 1.1). L’idiosome, qui représente le corps du Varroa, est formé
des boucliers ventral et dorsal, des quatre paires de pattes, courtes et fortes, et dotées
d’apothèles, leur permettant de s’accrocher à l’abdomen de l’hôte. (Rosenkranz et al., 2010).
D’autre part, les chélicères permettent aux varroas femelles de percer la cuticule d’une larve
ou d’une abeille adulte et de se nourrir de l’hémolymphe et des corps gras (Ramsey et al.,
2019).
Figure 1.1: Varroa destructor, parasite d'Apis mellifera, en vue dorsale (gauche) et ventrale
(droite). Les structures nommées dans le texte sont visibles : bouclier dorsal et ventral, les
pattes et apothèles ainsi que les chélicères. Photos prises par microscope par balayage
électronique par Cayambe (Licence Creative Commons).
1.2.1 Cycle de reproduction intimement lié à celui de son hôte
Dans une colonie infestée, la femelle Varroa peut être présente sous deux phases : la phase
phorétique et la phase de reproduction. La phase phorétique correspond à la phase
d’alimentation sur l’abeille adulte, soit entre les cycles de reproduction ou lorsque le couvain
est absent de la ruche. Bien qu’ils s’attachent préférentiellement aux abeilles nourrices via la
perception de kairomones, il est aussi possible que certains individus s’accrochent à des
ouvrières butineuses. Cela permet le transfert horizontal de plusieurs femelles varroas, qui
peuvent ainsi infester d’autres colonies à proximité (Nazzi et Le Conte, 2016). La figure 1.2
8illustre un cycle de reproduction complet du Varroa. La femelle Varroa en phase phorétique
est attirée par des kairomones émises par une larve au 5e instar (Nazzi et Le Conte, 2016) et
pénètre son alvéole quelques heures avant l’operculation pour s’y reproduire (Ifantidis,
1988). Puis, environ 70 h plus tard, elle pond un premier œuf, qu’elle ne fertilise pas et qui
se développera en mâle (Rosenkranz et al., 2010). Le deuxième œuf ainsi que les suivants,
pondus à des intervalles de 30 h, sont fécondés avec les spermatozoïdes de la spermathèque
de la mère et donneront des femelles varroas diploïdes.
Après quelques heures, l’œuf éclot et la progéniture se développe en passant par les stades
de proto- et deutonymphe, jusqu’à la mue finale, qui survient 5.8 et 6.6 jours plus tard pour
les femelles et les mâles respectivement (Donzé et Guérin, 1994; Ifantidis, 1983; Martin,
1994). Pour assurer leur développement jusqu’au moment de l’accouplement, la mère crée
un site d’alimentation sur le 5e segment de l’abdomen de la larve, dédié aux progénitures
(Donzé et Guérin, 1994). Les accouplements répétés ont lieu dès que le mâle et la première
fille ont atteint maturité, soit un peu moins de 11 jours après que la mère ait pénétré l’alvéole
(Donzé et Guérin, 1994; Ifantidis, 1983; Martin, 1994). La femelle possède une
spermathèque, vers laquelle les spermatozoïdes migrent pour y être entreposés. Comme le
cycle de reproduction du Varroa est limité par le temps de développement de la nymphe
d'abeille, d’une durée d’environ 12 jours pour l’ouvrière, la moyenne est de 1.3-1.45 filles
produites par cycle de reproduction (Martin 1994). Ce nombre augmente à 2.2-2.6 dans les
cellules de faux-bourdons, puisque le développement de la nymphe dure 14 jours (Martin,
1995). Les jeunes femelles varroas fécondées adéquatement pourront à leur tour réaliser entre
1,5 et 3 cycles de reproduction au cours de leur vie (Oldroyd et al., 1999 ; Rosenkranz et al.,
2010). La population de varroas dans une colonie d’abeilles dépend donc directement de la
quantité de couvain et du nombre de mois où le couvain est disponible dans la colonie.
9Figure 1.2: Schéma représentant la phase phorétique et la phase de reproduction de Varroa
destructor en parallèle avec le développement des pupes d'Apis mellifera. (MAPAQ, Varroa
destructor, mieux comprendre l’ennemi)
1.2.2 Relation parasitaire en déséquilibre
Un rapprochement des colonies d’Apis mellifera aux colonies d’Apis cerana, aurait permis
au parasite de cette dernière, Varroa jacobsoni de changer d’hôte (Oldroyd et al., 1999) et
une spéciation en Varroa destructor (Anderson et Truemann, 2000). Depuis le changement
d’hôte, V. jacobsoni et V. destructor démontrent des trajectoires évolutives bien distinctes
(Techer et al., 2019).
La relation hôte-parasite chez A. cerana est équilibrée par trois mécanismes de défense ayant
évolués pendant plusieurs milliers d’années. Premièrement, la reproduction des varroas est
limitée au couvain de faux-bourdons (Rath, 1999). Cela a un effet majeur sur le
développement de la population de varroas, puisque le couvain de faux-bourdon n’est
disponible qu’à certaines périodes de l’année. Deuxièmement, l’Abeille asiatique a des
comportements de défense active : le toilettage (grooming) et le comportement hygiénique
(Rath, 1999 ; Rosenkranz et al., 2010). Respectivement, le toilettage et le comportement
hygiénique visent à déloger les varroas sur les abeilles adultes et à l’intérieur des cellules de
couvain operculées. Enfin, un phénomène « de mise en tombeau » du couvain de faux-
10bourdon peut se produire si la larve est infestée de plusieurs varroas femelles (Rosenkranz et
al., 2010). Les pupes ainsi infestées sont trop faibles pour défaire l’opercule de cire et
émerger de l’alvéole (Rath, 1999). Ainsi toute la progéniture de varroas meurt enfermée dans
l’alvéole avec la pupe infestée, d’où le terme « mise en tombeau ». Ces traits exceptionnels
permettent donc à A. cerana de limiter l’augmentation de la population de varroas au sein de
ses colonies.
Par contre, le changement d’hôte récent du parasite vers A. mellifera engendre un
déséquilibre à l’avantage de V. destructor, puisque l’hôte n’a pas ces résistances naturelles
au parasite. L’évolution de ces adaptations au sein de l’espèce est ralentie par les pratiques
de contrôle en apiculture, car en traitant les colonies avec des acaricides, on diminue la
pression de sélection de la virulence du varroa sur son hôte (Locke et al., 2012).
1.2.3 La varroose
Lorsqu’une colonie de A. mellifera est fortement infestée par V. destructor, ce dernier cause
une maladie dévastatrice nommée varroose. Les symptômes de la maladie ne sont pas
uniformes, ce qui la rend difficile à diagnostiquer au début de l’infestation. En effet, son
intensité augmente de manière proportionnelle à l’augmentation de la population du parasite,
soit de manière exponentielle entre le printemps et l’automne pour les populations en climat
tempéré (Harris et al., 2003). De plus, l’action du parasite a des effets directs sur la santé des
individus, tandis que d’autres effets sont indirects (couplés à d’autres agents) et à l’échelle
de la colonie.
Les effets directs sont les blessures de perforation causées par les chélicères des femelles
pour se nourrir. C’est la perte d’hémolymphe et de corps gras (Ramsey et al., 2019) répétée
qui va avoir un effet à l’échelle de l’individu, tant sur les adultes que sur les larves infectées
(Boecking et Genersch, 2008). D’abord, le retrait d’hémolymphe pendant le développement
de la larve diminue le poids au stade adulte (Duay et al., 2003), réduit l’espérance de vie
Boecking et Genersch, 2008), et peut causer des malformations aux organes en
développement (Garedew et al., 2004). Ce phénomène diminue la capacité de vol des adultes,
autant pour les mâles que pour les ouvrières (Rinderer et al., 1999 ; Kralj et Fuchs, 2006). De
11plus, comme l’alimentation des varroas diminue la taille des faux-bourdons (Duay et al.,
2003), cela réduit par le fait même leur fitness, puisque leur taille est positivement corrélée
avec le nombre de spermatozoïdes qu’ils produisent (Schlüns et al., 2003).
Un autre effet néfaste de la consommation d’hémolymphe et de corps gras par les varroas est
l’affaiblissement du système immunitaire de l’abeille. En effet, le parasitisme réduit
l’expression des gènes de l’immunité chez les abeilles naissantes (Marche et al., 2019), en
plus de modifier plusieurs éléments de la réponse immunitaire en cascade. En consommant
l’hémolymphe de son hôte, le varroa diminue le nombre d’hématocytes présents, ce qui a
pour effet de réduire l’expression du gène de la propènol oxydase, une enzyme impliquée
dans la synthèse de mélanine. Comme la mélanine et les hématocytes participent à la guérison
des plaies et à l’encapsulation d’agents pathogènes dans l’hémolymphe (Kanost et Gorman,
2008), l’immunité et la capacité de guérison sont compromises (Yang et Cox-Forster, 2007).
L’effet indirect de V. destructor le plus notable est son rôle de vecteur de virus. En effet, les
symptômes observables dans la colonie tels que le couvain en mosaïque, la baisse rapide de
la population et la présence d’abeilles avec les ailes déformées sont plutôt attribuables aux
virus transmis par le parasite (Rosenkranz et al., 2010). Les cinq virus les plus transmis par
Varroa sont le virus de l’abeille Kashmir (KBV), le virus de la paralysie aigüe de l’abeille
(ABPV), le virus de la paralysie aigüe Israélienne (IAPV) et le virus des ailes déformées
(DWV) (Boecking et Genersch, 2008). Le virus des ailes déformés est particulièrement
débilitant, puisqu’il réduit la taille des individus infectés et atrophie leurs ailes (Figure 1.3),
ce qui rend plusieurs comportements impossibles.
Pour l’apiculteur, la varroose représente des pertes économiques importantes.
L’effondrement des colonies infestées de varroas survient souvent parce que le diagnostic à
un niveau d’infestation faible ou modéré est difficile. À ces niveaux, la colonie pourrait se
développer moins rapidement ou produire moins de miel sans que les symptômes soient
visibles. Lorsque les effets se font sentir à l’automne, quand la population de varroas est à
son maximum et que le couvain est fortement infesté, il est souvent trop tard pour éviter les
dommages à la colonie (Rosenkranz et al., 2010). En effet, c’est dans cette période qu’on
12observe une relation entre la chute naturelle de varroas et la composition corporelle des
abeilles d’hiver. Plus la chute naturelle est élevée, plus le poids des adultes est réduit,
résultant soit des effets directs du retrait d’hémolymphe et de corps gras, soit de soins reçus
déficients lorsque ces individus étaient à l’état de couvain (Aronsten et al., 2012 ; Wegener
et al., 2016). Ces abeilles étant donc moins nombreuses et ayant plus de difficulté à réguler
leur teneur en eau, l’homéostasie thermique de la grappe d’abeille à l’hiver est plus
problématique (Wegener et al., 2016). Lorsque l’infestation est trop importante, il y a
effondrement en moins de trois ans sans traitement (Fries et al., 2006). Pour s’assurer de
conserver les niveaux d’infestation à des seuils économiquement viables, il est recommandé
de traiter les colonies infestées à l’automne et au printemps en cas de besoin (Giovenazzo et
Dubreuil, 2011; MAPAQ, 2019).
Figure 1.3: Photo d’Apis mellifera sur un cadre de couvain dans une colonie fortement
infestée par Varroa. L'ouvrière encerclée présente des signes cliniques d'infection au Virus
des ailes déformées (ailes atrophiées). À la droite de cette dernière, on peut apercevoir un
varroa en phase phorétique sur le thorax d’une ouvrière. Photo par Marie-Lou Morin (2019).
1.2.4 Contrôle du Varroa par la lutte intégrée
Différentes approches sont utilisées par les apiculteurs pour réduire l’infestation de varroas
dans leurs colonies. L’approche préconisée est l’utilisation d’acaricides de synthèse à
13diffusion lente. Des bandes médicamentées imprégnées d’acaricides synthétiques comme le
fluvalinate-tau (ex : Apistan®), le Coumaphos (ex : CheckMite®) ou la formamidine
amitraze (ex : Apivar®) sont faciles à utiliser et efficaces pour tuer les varroas présents sur
les abeilles adultes. Par contre, des résistances à ces produits au sein de certaines populations
de varroas sont apparues (Milani, 1999 ; Sammataro et al., 2005 ; Maggi et al., 2010). Il
existe néanmoins d’autres options de traitements efficaces, soit les acides organiques comme
l’acide formique ou l’acide oxalique, ainsi que les huiles essentielles comme le thymol
(Baggio et al., 2004 ; Rademacher et Harz, 2006 ; Rosenkranz et al., 2010). Tout comme les
acaricides de synthèse, ces produits sont efficaces en absence de couvain dans la colonie,
puisque le produit atteint seulement les varroas en phase phorétique. Malheureusement, tous
ces acaricides produisent des résidus qui risquent de contaminer les produits de la ruche
(Bajuk et al., 2017, Tihelka, 2018). De plus, l’effet de ces pesticides n’est pas sans
répercussions sur la santé de leurs abeilles (Reeves et al., 2018, Tihelka, 2018). Il existe donc
un besoin urgent de réduire notre dépendance aux traitements chimiques et de développer de
nouvelles stratégies de gestion efficaces.
Certaines méthodes alternatives sont capables de limiter la croissance de la population de
varroas en cas d’infestation. Par exemple, l’utilisation d’un plateau grillagé sur le bas de la
ruche permet d’emprisonner les acariens qui y tombent. La technique de retrait de cadres de
couvain de faux-bourdons operculés permet quant à elle de retirer les varroas qui y sont
piégés pour se reproduire. Aussi, comme la taille de la population de varroas dépend de la
taille de la population d’abeilles, en imitant le phénomène d’essaimage par la division des
colonies, cela limite la croissance de la population de varroas (Loftus et al., 2016; Maucourt
et al. 2018). D’autres stratégies de contrôle biologique par des prédateurs comme
Stratiolaelaps scimitus sont explorées, mais n’ont pas encore donné des résultats concluants
lors d’études sur le terrain (Rangel et Ward, 2018; Rondeau et al. 2018, 2019).
On voit qu’il existe une variété d’outils disponibles pour effectuer la lutte intégrée au Varroa,
c’est-à-dire de lutter tout en réservant l’utilisation de pesticides qu’en dernier recours et en
priorisant les méthodes biologiques et mécaniques pour résister au parasite. L’une des
stratégies prometteuses dans la lutte intégrée aux maladies de A. mellifera est la sélection de
14Vous pouvez aussi lire

















































