Utilisation de l'habitat par le cerf de Virginie après plus d'une dizaine d'années d'exclusion sur l'île d'Anticosti - Mémoire Aimie Baribeau ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Utilisation de l'habitat par le cerf de Virginie après plus
d'une dizaine d'années d'exclusion sur l'île d'Anticosti
Mémoire
Aimie Baribeau
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Aimie Baribeau, 2022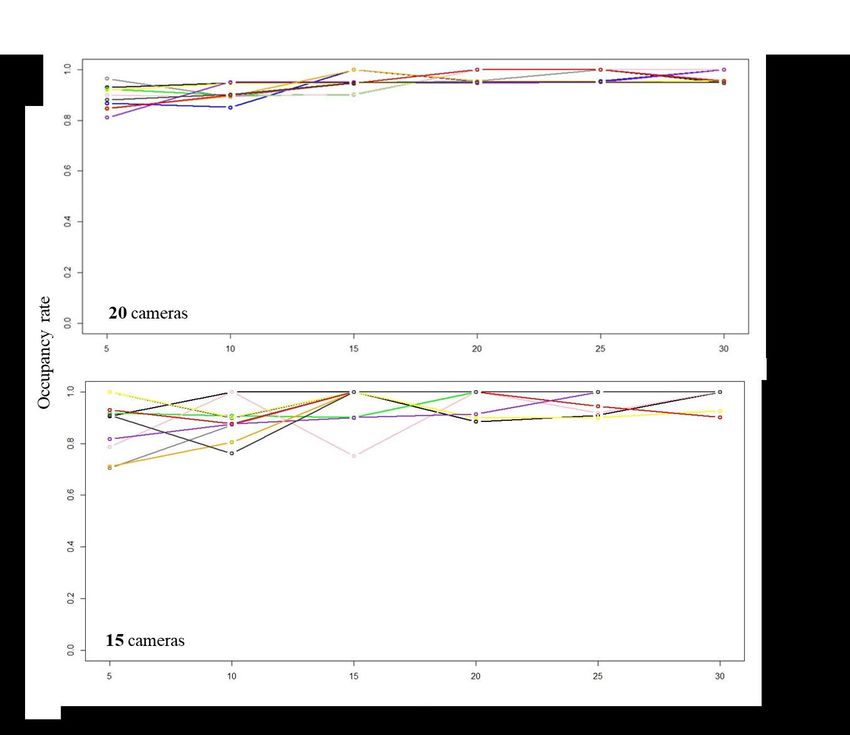
RÉSUMÉ
Le broutement sélectif par les grands herbivores peut altérer la composition et la structure
végétale d’un milieu. À l’île d’Anticosti (Québec, Canada), la forte densité de cerfs de
Virginie (Odocoileus virginianus) broutant le sapin baumier (Abies balsamea) comme
ressource hivernale induit une conversion des sapinières en pessières et en landes à épinettes
blanches (Picea glauca). Afin de permettre la régénération des sapinières, un plan général
d’aménagement intégré des ressources du milieu forestier a été mis en œuvre avec pour
stratégie phare la diminution de la densité du cerf de chantiers de coupe clôturés pendant une
douzaine d’années. L’objectif de cette étude était d’évaluer l’utilisation de l’habitat par les
cerfs à la suite du démantèlement des exclos. Durant l’été 2020, nous avons distribué un
réseau de caméras de surveillance dans six exclos démantelés depuis trois à huit ans afin de
déterminer les probabilités d’utilisation de l’habitat restauré par les cerfs selon les attributs
de l’habitat. Nous avons aussi récolté des informations sur l’environnement près des caméras
notamment la composition en espèces d’arbres ainsi que de la strate végétale au sol. Les
probabilités d’utilisation par les cerfs de l’habitat restauré étaient plus élevées lorsque les
sites de caméras présentaient une couverture abondante au sol en cornouiller du Canada
(Cornus canadensis). Nous avons aussi utilisé des mesures de distances entre les caméras et
la bordure des exclos ainsi qu’entre les caméras et les îlots de forêt résiduelle, ainsi que des
mesures de superficie d’îlots forestiers comme variables de la configuration spatiale des
exclos. Aucune de ces variables n’a expliqué l’utilisation de l’habitat par les cerfs. Nos
résultats permettent de mieux comprendre l’utilisation de l’habitat d’été du cerf de Virginie
sur l’île d’Anticosti dans un contexte d’aménagement forestier en illustrant l'importance de
l’abondance en ressources alimentaires.
iiABSTRACT
Selective browsing by large herbivores can alter the composition and the vegetation structure
of an area. On Anticosti Island (Québec, Canada), the high density of white-tailed deer
(Odocoileus virginianus) browsing on balsam fir (Abies balsamea) as a winter resource has
led to the conversion of fir stands to spruce stands and white spruce (Picea glauca) parklands.
To allow the regeneration of fir stands, a general integrated management plan of forest
resources was implemented with the main strategy of nearly excluding deer from logged and
fenced sites for more than a decade. Our objective was to evaluate restored habitat use by
deer following the removal of exclosures. During summer 2020, we distributed a network of
camera traps in six exclosures dismantled in the last three to eight years to assess the
probabilities of restored habitat use by deer according to habitat composition. We collected
information on the characteristics of the environment surrounding at each camera site
including tree species composition as well as ground vegetation layer. Probabilities of
restored habitat use by deer were higher when camera sites had higher ground cover of
Canadian dogwood (Cornus canadensis). To represent the spatial configuration of
exclosures, we also used measures of distance from cameras to the edge of exclosures and
from cameras to residual forest patches, as well as measures of forest patch areas. None of
these variables explained deer habitat use. Our results allow a better understanding of white-
tailed deer restored habitat use during summer on Anticosti Island in a forest management
context highlighting the importance of forage abundance.
iiiTABLE DES MATIÈRES
RÉSUMÉ ................................................................................................................................ ii
ABSTRACT .......................................................................................................................... iii
Liste des tableaux ................................................................................................................... v
Liste des figures ..................................................................................................................... vi
Liste des annexes .................................................................................................................. vii
Remerciements .................................................................................................................... viii
Avant-propos .......................................................................................................................... x
INTRODUCTION .................................................................................................................. 1
CHAPITRE 1: Habitat use by white-tailed deer after more than a decade of exclusion in the
boreal forest……….….………………………………………………………….................10
Résumé.............................................................................................................................. 11
Abstract ............................................................................................................................. 12
Introduction ....................................................................................................................... 13
Materials and Methods ...................................................................................................... 15
Results ............................................................................................................................... 19
Discussion ......................................................................................................................... 22
Acknowledgments ............................................................................................................ 25
Literature cited .................................................................................................................. 26
CONCLUSION GÉNÉRALE .............................................................................................. 30
BIBLIOGRAPHIE GÉNÉRALE ......................................................................................... 36
ANNEXES ........................................................................................................................... 43
ivLISTE DES TABLEAUX
Table 1. Model comparison for variables influencing the detection probability (p) of white-
tailed deer using camera traps in former management exclosures (n=6) on Anticosti Island,
Canada. The occupancy (Ψ) is fixed at 1 (null).
Table 2. All occupancy models and the null model for the probability that white-tailed deer
use restored habitat (Ψ) on Anticosti Island (Canada) following 10-12 years of exclusion in
clear-cuts and forested areas (n=6 exclosures).
Table 3. Means and standard deviations (sd) used to center and scale all the variables
included in the models.
vLISTE DES FIGURES
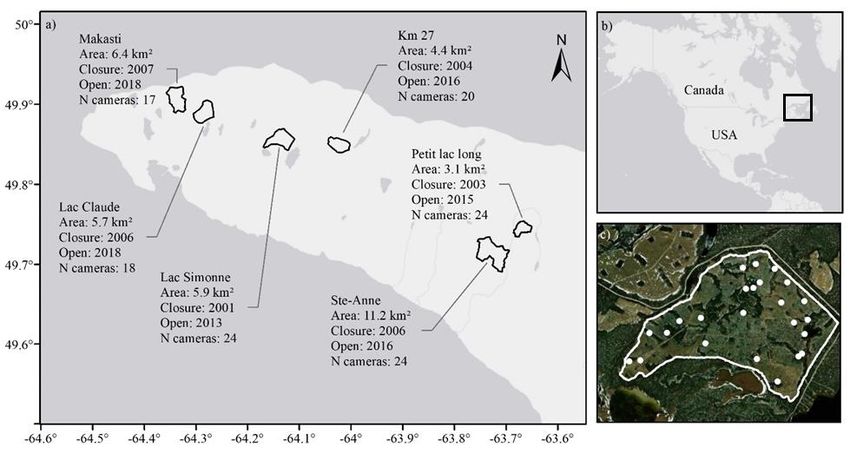
Figure 1. Location of the study area and attributes of the 6 exclosures under study on
Anticosti Island, Québec, Canada. In a) the specific information of every exclosure and their
position on the western part of the island, b) the location of Anticosti Island on the North
American continent and c) magnification of the Km 27 exclosure to show the camera
distribution.
Figure 2. The probability that white-tailed deer use restored habitat after the removal of
exclosures on Anticosti Island (Canada) in relation with the percentage of cover of Cornus
canadensis. The upper and lower dashed lines are 95% confidence intervals.
Figure 3. Occupancy rate to determine sampling effort according to a combination of days,
number of cameras and time intervals between every picture taken by the cameras. The
colored lines are the different time interval between pictures.
Figure 4. Detection probability (p) at 5 meters (a) and 10 meters (b) in front of the camera
trap sites in exclosures on Anticosti Island (Canada) during summer, 2020. The upper and
lower dashed lines are the 95% confidence intervals.
viLISTE DES ANNEXES
Appendix 1. Occupancy rate used to determine sampling effort according to data collected
on a pilot study in 2019 on Anticosti Island.
Appendix 2. Main assumptions necessary for the initial single-season single-species
occupancy model.
Appendix 3. Table including all mean and standard deviation (sd) of the scaled variable
included in the models.
Appendix 4. Graphics of maximal detection probabilities at 5 and 10 meters in front of the
camera.
viiREMERCIEMENTS
Je voudrais tout d’abord remercier mon directeur Jean-Pierre Tremblay. Merci de m’avoir
fait confiance pour la réalisation de ce projet. Mon parcours ne fut pas l’un des plus faciles,
mais j’ai apprécié ton support tout au long de ces deux dernières années. En espérant avoir
été à la hauteur de tes attentes.
Je voudrais aussi remercier mon co-directeur Steeve Côté. Merci également de m’avoir fait
confiance pour le projet. Tes commentaires ont toujours été pertinents et ont contribué à
l’amélioration de mon travail. J’ai commencé mon parcours en n’ayant qu’une mince idée de
l’ampleur de la tâche qui m’attendait, merci de m’avoir appuyée tout au long de celle-ci.
Un merci spécial pour Julien Hénault-Richard. Merci de m’avoir aidée avec toutes les étapes
de mon projet. De la gestion à l’analyse de mes données, tu as contribué à rendre chaque
étape de mon projet un peu plus clair! Merci aussi à Sandra Hamel pour tes conseils et
explications pour mes analyses statistiques. Tu m’as beaucoup aidée à cheminer et à
comprendre ce que je faisais tant au niveau pratique que théorique. Sans toi, la courbe
d’apprentissage aurait été beaucoup plus abrupte.
Je ne peux pas passer à côté de remercier mes aides sur le terrain :
Catherine Beaulieu, sans toi, mon terrain aurait été beaucoup plus difficile. Ton
expérience, ton humour et notre compatibilité a rendu cet été haut en émotions beaucoup plus
supportable et je t’en suis infiniment reconnaissante. Nos fous rires et nos aventures me
manquent, mais PU JAMAIS haha.
Baptiste Brault, merci d’avoir contribué à mon projet, d’y avoir apporté ton souci du
détail. Il est vrai que notre été fut assez chargé sur plusieurs aspects. Nous avons eu de bons
moments malgré tout, ce sont ces moments que je garde en mémoire et je suis reconnaissante
de ton aide.
viiiMerci aussi aux membres du labo Côté/Tremblay. Cette dernière année de pandémie ne nous
a pas permis de bien se connaître. Du moins, j’ai apprécié votre accueil et votre aide lors de
ma première session. Merci à Barbara pour m’avoir aidée avec mon écriture, tes
commentaires ont toujours été justes et utiles. Merci aussi à Catherine pour m’avoir offert
son aide pour mon séminaire et merci à Delphine de m’avoir écoutée à un moment où j’en
avais le plus de besoin, tes conseils m’ont été précieux pour persévérer en fin de parcours.
Finalement, merci à mes parents de m’avoir supportée tout au long de mon parcours parsemé
de hauts et de bas. Merci aussi à mon copain Julien de m’avoir supportée dans les moments
plus difficiles, ton écoute, ton soutien et ton amour m’ont beaucoup aidée.
ixAVANT-PROPOS
L’objectif de ce mémoire de maîtrise était de déterminer l’utilisation de l’habitat par le cerf
de Virginie dans six exclos démantelés à la suite de l’application du plan général
d’aménagement intégré des ressources du milieu forestier sur l’île d’Anticosti (Québec,
Canada). Pour ce faire, nous avons stratifié différents types d’habitat des exclos et déployé
aléatoirement dans ces strates un total de 25 caméras par exclos à l’été 2020. Les caméras
ont pris des photos aux cinq minutes durant 14 jours. Nous avons utilisé les données sous la
forme présence/absence de cerfs dans différents modèles d’occupation avec des
combinaisons et interactions de variables de végétation et de configurations spatiales.
Ce mémoire comporte 4 parties : une introduction générale, un chapitre principal présenté à
la façon d’un article scientifique rédigé en anglais en vue d’une future publication dans une
revue scientifique, une annexe présentant des compléments aux matériels et méthodes et aux
résultats ainsi qu’une conclusion générale. En tant que première auteure de l’article, j’étais
responsable du développement des hypothèses et prédictions, de l’élaboration et la mise en
pratique des protocoles, de la récolte des données sur le terrain, de la gestion, compilation et
de l’analyse des données ainsi que de la rédaction. Les deux chercheurs coauteurs de l’article
sont :
- Jean-Pierre Tremblay (Ph.D), directeur de maîtrise et professeur agrégé au département de
biologie de l’Université Laval.
- Steeve D. Côté (Ph.D), codirecteur de maîtrise et professeur titulaire au département de
biologie de l’Université Laval.
xINTRODUCTION
Les herbivores sont des animaux utilisant les espèces végétales comme ressources
alimentaires. Que ce soit par l’ingestion d’herbacées, de jeunes pousses ligneuses, de
mousses et lichens, de feuilles d’arbres ou bien de ramilles (Nixon et al. 1970, Bergerud
1972), les herbivores contribuent de différentes façons à modifier l’environnement dans
lequel ils évoluent. Le broutement sélectif sur les espèces végétales peut altérer la
régénération naturelle, les communautés végétales et les fonctions des forêts (Rooney 2001,
Kuijper et al. 2010). L'exclusion de grands herbivores après les coupes forestières peut être
utilisée pour prévenir les effets négatifs du broutement sur les forêts. Certaines études ont
montré l'efficacité de cette stratégie pour atteindre les objectifs de gestion forestière et pour
identifier les densités d'herbivores compatibles avec la dynamique de régénération naturelle
des forêts gérées (Kay et Bartos 2000, Horsley et al. 2003, Persson et al. 2007). L'évaluation
de la fréquentation des habitats protégés après l’exclusion est essentielle pour comprendre
comment les grands herbivores utilisent les paysages aménagés.
Impact des grands herbivores sur la composition et la structure des forêts
Les herbivores peuvent altérer la régénération des gaules d’arbres d’espèces
sélectionnées composant leur environnement en affectant la production de feuilles ainsi que
les structures de reproduction (Frelich et Lorimer 1985, Kaien 2006, Russell et al. 2017). Le
cas de l’orignal (Alces alces) au parc national de l’île Royale (É.-U.) en hiver montre que la
sélection d’une espèce végétale diminue sa régénération privant l’herbivore à long terme de
sa principale source d’alimentation. En effet, l’orignal se nourrit en été d’espèces à feuilles
caduques et de plantes herbacées, et en hiver de ramilles de sapin baumier en évitant de
brouter l’épinette (Picea spp.) (Timmermann et McNicol 1988, Augustine et McNaughton
1998). Ce broutement intensif réduit la régénération des espèces de feuillus et supprime la
régénération du sapin baumier (Augustine et McNaughton 1998). Conséquemment, les
épinettes, qui ne sont pas utilisées par les orignaux, finissent par dominer le paysage,
réduisant ainsi la principale ressource alimentaire hivernale de l’orignal (Brandner et al.
1990, McInnes et al. 1992, Augustine et McNaughton 1998).
1Un scénario similaire peut être observé à l’île d’Anticosti située au Québec. Les cerfs
de Virginie présentent un régime alimentaire se composant en majorité de sapin baumier
durant la saison hivernale (Lefort et al. 2007). Le broutement sélectif à long terme de cette
espèce végétale cause un ralentissement de sa régénération naturelle par recrutement de
semis. Un remplacement des sapinières par des pessières blanches et des landes à épinettes
blanches est observé sur l’île (Potvin et al. 2003, Barrette et al. 2014, Barrette et al. 2017). Il
est donc estimé que la majorité des peuplements de sapin seront inexistants d’ici 30 à 50 ans
(Potvin et al. 2003, Barrette et al. 2014, Barrette et al. 2017). Un autre exemple de l’impact
du broutement par les cervidés est celui du remplacement de la pruche du Canada (Tsuga
canadensis) par l’érable à sucre (Acer saccharum) au Michigan. Ce changement dans la
dominance semble être causé par le broutement sélectif du cerf de Virginie sur la pruche
(Frelich et Lorimer 1985). Engelmark et ses collaborateurs (1998) ont démontré que le
remplacement du bouleau (Betula sp.) par l’épinette (Picea abies) dans le nord de la Suède
serait en partie causé par le broutement des rennes (Rangifer tarandus).
La sélection d’habitat
À large échelle, la sélection d’habitat par les grands herbivores et la distribution des
espèces peuvent être influencées par la taille et la configuration spatiale des parcelles de
forêts (With et al. 1997, Royle et Dorazio 2008, Webb et al. 2009). Un habitat, dans ce
contexte, est le milieu géographique que l’animal utilise pour établir son domaine vital. Les
animaux sélectionnent un habitat où la disponibilité en ressources alimentaires est élevée,
mais aussi en fonction de la protection que ceux-ci peuvent leur offrir, selon la nature, la
forme et l’arrangement spatial des parcelles d’habitat (With et al. 1997, Tischendorf et Fahrig
2000, Bélisle 2005). Selon Johnson (1980), une sélection est un processus durant lequel un
animal choisit une ressource et qui dépend de sa disponibilité, c’est-à-dire ce qui est
accessible au consommateur.
À une échelle plus fine, la sélection d’habitat inclut les ressources alimentaires
choisies par le consommateur (Johnson 1980). Pour caractériser l’impact des espèces
animales sur l’habitat, il est nécessaire de déterminer quelles sont les espèces sélectionnées
2par les grands herbivores, et donc qui sont potentiellement plus susceptibles à la pression de
broutement sélectif (Nugent et al. 2001).
Selon la saison, la sélection d’habitat peut varier. Les deux saisons extrêmes en regard
de l’abondance en ressources alimentaires sont l’hiver et l’été. En hiver, les cerfs de Virginie
se retrouvent souvent à densité élevée dans des zones avec des ressources alimentaires
limitées (Ozoga et Gysel 1972, Brown et Doucet 1991). Les stratégies d’acquisition des
ressources changent en fonction de la disponibilité limitée des ressources (Nudds 1980). La
recherche d’abris devient un enjeu plus important pour la conservation d’énergie (Lang et
Gates 1985). Les cerfs se situant dans les latitudes plus au nord utilisent des habitats de
conifères comme abris, l’utilisation étant plus grande lorsque la température est basse pour
minimiser les pertes de chaleur (Ozoga et Gysel 1972). Ces habitats sont choisis pour leur
plus faible épaisseur de neige, des températures plus clémentes et moins fluctuantes et un
impact plus faible du vent (Lang et Gates 1985). Un couvert forestier continu est un facteur
important et recherché par les cerfs, car il diminue les accumulations de neige (Dumont 2004,
Massé et Côté 2012b) et réduit la perte de chaleur (Cook et al. 1998).
En été, les herbivores n’ont pas la contrainte des températures froides et de la neige
pour leurs déplacements. Pour ce qui est des cerfs, la saison estivale est la période de l’année
la plus productive en biomasse végétale. C’est donc une période critique pour l’accumulation
de réserves corporelles nécessaire à l’allaitement des jeunes, à la reproduction des femelles
à l’automne suivant ainsi qu’à la survie pour la saison hivernale à venir (Stewart et al. 2005,
Therrien et al. 2007). L’ingestion de plantes à forte teneur en protéines permet aux cervidés
d’accumuler les réserves nécessaires pour les saisons automnale et hivernale (DelGiudice et
al. 1992, Tixier et al. 1997). Lorsque disponibles, les plantes herbacées à feuilles larges sont
généralement sélectionnées par les cerfs (Chamrad et Box 1968, McCullough 1985,
Tremblay et al. 2006). Les résultats d’une étude menée par Simard et ses collaborateurs
(2014) ont montré qu’en été, les habitats présentant une forte biomasse en espèces végétales
peuvent favoriser la première ovulation des jeunes femelles cerfs de Virginie ainsi que le
succès de sevrage des jeunes. De plus, il a été conclu qu’une alimentation pauvre en été
pourrait réduire les taux de gestation des cerfs femelles en diminuant les chances d'œstrus et
d'ovulation (Cook et al. 2001, Stewart et al. 2005) ainsi qu’en retardant la conception (Cook
et al. 2001).
3Des études de sélection d’habitat en saison estivale ont aussi été réalisées à l’île
d’Anticosti permettant d’acquérir des connaissances quant à l’utilisation de différents types
d’habitats par les cerfs de Virginie. En effet, selon Massé et Côté (2009), la sélection
d’habitat à densité de population élevée et en absence de prédation en été est déterminée par
l’abondance des ressources alimentaires plutôt qu’un compromis entre l’acquisition de
nourriture et le besoin de protection. Les résultats montrent aussi que la distance entre une
bordure couvert forestier/coupe n’est pas un facteur influençant la sélection d’habitat par les
cerfs. Également, les cerfs en été sélectionnent différentes ressources alimentaires à
différentes échelles (Massé et Côté 2012b). Par exemple, il y a une sélection des tourbières
à l’échelle du paysage et une sélection des trouées forestières à l’échelle du peuplement
forestier. De plus, la taille du domaine vital des femelles diminue en fonction de
l’augmentation de la présence de plantes fourragères et d’arbustes, car leurs besoins
énergétiques sont comblés sans devoir se déplacer sur une plus grande distance (Massé et
Côté 2012b).
Stratégie de gestion forestière
L’utilisation d’exclos comme stratégie d’aménagement forestier est une méthode
utilisée pour induire une régénération naturelle des espèces végétales sous broutement
intensif (Russell et al. 2001). Formellement, les exclos sont des secteurs de forêt dont les
herbivores sont exclus par des clôtures pour favoriser la régénération des espèces se trouvant
à l’intérieur. À plus large échelle, il peut s’avérer difficile d’exclure tous les herbivores d’une
aire clôturée, alors le terme « exclos » peut parfois s’appliquer suite à une diminution
marquée de la population d’herbivores dans une telle aire. Plusieurs études ont utilisé cette
méthode et les résultats sont similaires : une diminution de la pression de broutement entraîne
une régénération des espèces végétales sélectionnées par les herbivores (Kay et Bartos 2000,
Horsley et al. 2003, Perrin et al. 2006). Dans une étude réalisée sur l’île d’Anticosti, des
comparaisons ont été faites entre des parcelles de sapinières clôturées et non clôturées de
même taille et distantes de 2 m (Potvin et Poirier 2004). Après 5 ans, une augmentation du
nombre de semis de sapin de taille moyenne (5-30 cm) a été observée dans les parcelles
clôturées ainsi qu’une augmentation du pourcentage de recouvrement des espèces. Par
4exemple, le sapin baumier et le bouleau à papier (Betula papyrifera) ont multiplié leur
recouvrement par dix en l’absence de broutement par le cerf de Virginie (Potvin et Poirier
2004).
Des résultats similaires ont été démontré dans une étude de Horsley et collaborateurs
(2003) en Pennsylvanie sur des exclos avec différents traitements de coupes (coupe totale,
éclaircie, sans coupe) et de densités de cerf de Virginie durant 10 ans. En effet, les tendances
observées pour les coupes totales étaient que la hauteur ainsi que le nombre de tiges d’espèces
telles que le cerisier noir (Prunus serotina), l’érable de Pennsylvanie (Acer pensylvanicum),
l’érable rouge (Acer rubrum), le bouleau noir et jaune (Betula lenta et B. lutea) et le cerisier
de Pennsylvanie (Prunus pensylvanica) augmentaient 3 à 5 ans après le début de l’expérience
et lorsque la densité de cerfs diminuait. Pour les zones d’éclaircies, l’abondance d’espèces
herbacées avait déclinée, mais la hauteur des grands arbres avait augmenté de près de 5 m
après 10 ans. Pour les zones non coupées, la densité de semis a augmenté en général durant
les 10 années de l’étude, mais les plus hautes tiges étaient quand même accessibles au
broutement (Horsley et al. 2003). Finalement, les résultats de Tripler et collaborateurs (2005)
révèlent aussi une croissance accélérée de certaines espèces végétales, notamment les gaules
de chêne rouge (Quercus rubra) et de pruche (Tsuga canadensis), qui étaient sous forte
pression de broutement avant l’exclusion des cerfs de Virginie. Ces nombreux exemples de
l’efficacité des exclos pour induire une régénération naturelle fait de cette technique
forestière une méthode empirique efficace pour mesurer l’effet du broutement des cerfs sur
les communautés végétales (Coomes et al. 2003).
Cette stratégie d’aménagement forestier a aussi été implantée à l’île d’Anticosti pour
assurer une cohabitation entre le cerf et son habitat dans une perspective de conservation des
espèces végétales et animales. À la fin du XIXe siècle, il y a eu une introduction volontaire
d’environ 220 cerfs de Virginie sur cette île. L’île ne présentait pas beaucoup de prédateurs
naturels du cerf ni de compétition interspécifique. À l’époque, il y avait de l’ours noir (Ursus
americanus), mais cette espèce est disparue de l’île au cours des années 1990 (Côté 2005).
La faible présence de prédateurs naturels du cerf a mené à une surabondance du cerf en une
trentaine d’années, entraînant une dégradation de l’habitat (Beaupré et al. 2005). Cette
surabondance a créé un problème pour l’écosystème forestier et la biodiversité végétale de
l’île (Bachand et al. 2015). Lors de la gestion de la relation entre le cerf et la forêt, le défi est
5de maintenir des densités de cerfs compatibles avec la régénération naturelle de la forêt, car
ce cervidé est à la fois une ressource économique pour l’île (Labonté 2015), mais aussi un
stress chronique sur les écosystèmes forestiers (Beguin et al. 2016). C’est cette relation
particulière entre le cerf et la forêt qui a mené à l’implantation de 29 sections aménagées
contenant coupes, plantations de sapins et îlots de forêt résiduelle clôturés (exclos).
Les exclos, d’une durée de vie d’environ 12 à 15 ans et recouvrant plus de 180 km2
de la partie centrale ouest de l’île d’Anticosti, ont été installés entre 2001 et 2017 (Labonté
2015). Cet intervalle de temps permet aux jeunes sapins d’atteindre une hauteur généralement
hors d’atteinte de broutement, soit au moins deux mètres (Beaupré et al. 2005). Il y a eu un
effort de chasse considérable à l’intérieur des exclos pour diminuer les densités de cerfs à
moins de 15 cerfs/km2, voire moins, afin de préserver le plus grand nombre de semis naturels
et plantés (Beaupré et al. 2005). La configuration spatiale des exclos se compose d’un
entremêlement entre des îlots de forêts résiduelles et des coupes incluant des plantations de
sapins pour fournir des zones d’abris et d’alimentation aux cerfs après le retrait des clôtures.
Évaluation de l’utilisation de l’habitat en écologie
Évaluer l’utilisation de l’habitat en écologie permet de mieux comprendre et étudier
les comportements d’espèces animales d’intérêt. Pour étudier ces comportements, il est
souvent nécessaire d’obtenir des données des déplacements spatiaux des espèces animales
dans différents milieux pour déterminer l’utilisation des différents habitats. Une méthode non
invasive disponible pour étudier la fréquentation de l’habitat est l’utilisation de caméras de
surveillance (O'Connell et al. 2011). En effet, les données des caméras peuvent être analysées
à partir d’approches modernes qui permettent d’évaluer à la fois le taux d’occupation et les
probabilités de détection de différentes espèces à plusieurs sites et dans différents types
d’habitat tout en incluant des variables pouvant affecter ces deux paramètres (Moeller et al.
2018). Les caméras de surveillance permettent de répondre à des questions à de plus larges
échelles spatio-temporelles et pour un plus grand nombre d'espèces animales dans une plus
vaste variété de milieux naturels (Delisle et al. 2021). Pour analyser les données associées
aux caméras, les modèles d’occupation sont souvent utilisés car ils intègrent à la fois les
probabilités d’occupation des espèces animales sous-étude, mais aussi les probabilités de
6détection des caméras. La probabilité de détection est la probabilité que l'espèce d'intérêt soit
présente au moment de l'enquête et détectée dans les types d’habitat utilisés (MacKenzie et
al. 2018). Dans un système ouvert, elle peut avoir un impact sur l'estimation de la probabilité
d’utilisation, qui est la probabilité que les types d’habitat soient utilisés par l'espèce cible
(MacKenzie et al. 2018). Donc, modéliser les deux paramètres à l’aide de variables pouvant
influencer ceux-ci permet d’obtenir des taux d’occupation des espèces animales non-biaisés
par la détectabilité.
Objectif, hypothèses et prédictions de l’étude
L’objectif de mon projet consiste à évaluer comment le cerf de Virginie utilise
l’habitat après le retrait des clôtures d’exclos érigés dans le cadre du plan général
d’aménagement forestier des ressources de l’île d’Anticosti. Plus spécifiquement, nous
évaluerons les probabilités d’utilisation de l’habitat restauré par les cerfs en fonction de la
configuration spatiale et de la composition en espèces végétales des exclos démantelés. Nos
hypothèses et prédictions sont les suivantes :
(1) L’intégration des exclos dans la sélection de l’habitat des cerfs est faite en fonction
de leur configuration spatiale.
Nous prédisons que la probabilité d’utilisation de l’habitat restauré par les cerfs sera
plus élevée dans les habitats se retrouvant en bordure de l’exclos, car la portion d’habitat
déclôturé n’est disponible que depuis trois à huit ans. Nous prédisons également que plus les
distances aux abris seront courtes, plus la probabilité d’utilisation de l’habitat restauré par les
cerfs sera faible, car en saison estivale et en situation d’absence de prédation, les cerfs ont
moins recours au couvert forestier (Massé et Côté 2009). Également, nous prédisons que la
probabilité d’utilisation de l’habitat restauré par les cerfs sera plus élevée près des îlots de
forêt résiduelle présentant une plus grande superficie, car ils représentent un plus grand
couvert de fuite.
(2) L’utilisation de l’habitat par le cerf dans les exclos en été reflète la nécessité
d’acquérir des réserves corporelles en préparation à la saison hivernale (Stewart et al. 2005).
Nous prédisons que la probabilité d’utilisation de l’habitat restauré par les cerfs sera
proportionnelle à la disponibilité des ressources alimentaires (Massé et Côté 2009). Nous
7prédisons aussi que la probabilité d’utilisation par les cerfs de l’habitat restauré sera plus
élevée dans les milieux ayant une surface terrière plus élevée en sapin baumier et en bouleau
à papier que dans les milieux ayant une surface terrière plus élevée en épinettes noire et
blanche.
Approches méthodologiques
Afin de tester les hypothèses ci-haut, nous avons utilisé des caméras de surveillance
déployées dans six exclos déclôturés depuis trois à huit ans sur la partie centrale-ouest de
l’île d’Anticosti de juin à août 2020. Nous avons déployé 25 caméras par exclos selon une
distribution aléatoire dans trois strates : 1) parterre de coupe en régénération, 2) forêt mature
dominée par le sapin baumier et 3) forêt mature dominée par les épinettes. Nous avons
échantillonné deux exclos à la fois à l’aide de caméras de surveillance programmées pour
prendre systématiquement une photo aux cinq minutes durant une période de 14 jours par
exclos. Sur les 150 caméras déployées (25 caméras * 6 exclos), 127 ont obtenu des photos
utilisables pour la période 14 jours, pour un total de 485 338 photos.
À chaque site de caméra, nous avons réalisé un inventaire forestier des gaules
d’espèces ligneuses par classe de hauteur et de diamètre ainsi que le recouvrement d’un sous-
ensemble des espèces végétales au sol. Ces variables ont été récoltées pour décrire la
composition en espèces végétales de chaque site de caméra. Finalement, nous avons mesuré
le pourcentage de visibilité latérale devant la caméra à des distances de 5 et 10 m à l’aide
d’une planche à profil pour modéliser les probabilités de détection de chaque caméra. Pour
les variables de la configuration spatiale de chaque exclos démantelé, nous avons utilisé le
logiciel ArcMap pour déterminer les distances entre chaque caméra et les îlots forestiers ainsi
que les bordures des exclos, de même que la superficie de chaque îlot forestier.
Pour le classement des photos en présence et absence de cerfs pour déterminer les
probabilités d’utilisation de l’habitat restauré par les cerfs, nous avons utilisé un programme
de reconnaissance visuelle. Nous avons choisi l’architecture d’EfficienceNet par couches
entraînées pour reconnaitre les cerfs sur les différents sites de nos caméras et classer plus de
8400 000 photos automatiquement en présence ou absence de cerf avec un degré de précision
de 94%. Nous avons utilisé ce classement pour obtenir la matrice finale d’observation de
présence/absence de cerfs par période de 24h pour nos 127 sites de caméras durant 14 jours.
Avec cette matrice ainsi que la table incluant les données d’inventaires forestiers et nos
mesures obtenues par ArcMap, nous avons procédé à l’évaluation de 20 modèles candidats
d’occupation. Ces modèles se composaient d’un paramètre modélisant les probabilités
d’utilisation de l’habitat restauré par les cerfs en fonction des différentes variables de
configuration spatiale et de composition en espèces végétales et d’un paramètre modélisant
les probabilités de détection en fonction des variables de pourcentage de visibilité à chaque
site de caméra.
9CHAPITRE 1
Habitat use by white-tailed deer after more than a decade of exclusion in the boreal forest
AIMIE BARIBEAU1, JEAN-PIERRE TREMBLAY2, STEEVE D. CÔTÉ3
1 Aménagement intégré Cerfs-forêt à l’île d’Anticosti;
Département de biologie, Université Laval, Québec, QC G1V 0A6, Canada
Courriel : aimie.baribeau.1@ulaval.ca
2 Aménagement intégré Cerfs-forêt à l’île d’Anticosti;
Département de biologie, Université Laval, Québec, QC G1V 0A6, Canada
Centre d’études nordiques et Centre d’étude de la forêt
Courriel : jean-pierre.tremblay@bio.ulaval.ca
3 Aménagement intégré Cerfs-forêt à l’île d’Anticosti;
Département de biologie, Université Laval, Québec, QC G1V 0A6, Canada
Centre d’études nordiques
Courriel : steeve.cote@bio.ulaval.ca
10RÉSUMÉ
L’exclusion d’herbivores dans certains secteurs forestiers est une stratégie utilisée
pour diminuer l’impact de leur broutement sélectif et induire la régénération naturelle des
espèces végétales désirées. Sur l’île d’Anticosti (Québec, Canada), le broutement sélectif par
le cerf de Virginie pose un problème pour la régénération des sapinières à bouleau à papier
menant à la conversion vers des forêts d’épinettes blanches. Des exclos dans lesquels des
coupes totales et des îlots de forêts résiduelles s’entremêlent ont été instaurés de 2001 à 2006
pour aider à la régénération naturelle des sapinières et fournir des abris et de la nourriture
aux cerfs. Notre objectif était d’évaluer comment le cerf utilise l’habitat suite au
démantèlement des exclos après plus d’une décennie, et ce en fonction de leur configuration
spatiale et de leur composition en espèces végétales. Nous avons distribué aléatoirement un
réseau de caméras automatiques en été pour des périodes de 14 jours dans six exclos de 3,1
à 11,2 km2 (n=25 caméras par exclos) où les cerfs ont été exclus pendant 10 à 12 ans. Nous
avons comparé des modèles candidats comprenant des variables de configuration spatiale et
de ressources alimentaires susceptibles d’expliquer l’utilisation des habitats par les cerfs tout
en contrôlant simultanément pour les variables affectant le taux de détection par caméra telles
que le pourcentage de visibilité devant les caméras. Notre modèle d’occupation le plus
supporté révèle que l’utilisation par les cerfs augmente de 19% lorsque les ressources
alimentaires, représentées par le recouvrement au sol en cornouiller du Canada, augmente de
0 à 100%. Aucune variable de la configuration spatiale des exclos (distance entre la limite
des exclos et les caméras, distance entre les parcelles de forêt et les caméras et taille des
parcelles de forêt) n’a influencé l’utilisation de l’habitat par les cerfs. Nous concluons que
les facteurs affectant l’utilisation de jeunes forêts par les cerfs après une période d’exclusion
sont majoritairement associés à la disponibilité en ressources alimentaires et donc, tous
travaux d’aménagement qui améliorent la production de nourriture en été devraient aider à
maintenir ou augmenter l’utilisation de l’habitat par les cerfs.
11ABSTRACT
The exclusion of herbivores in forest areas is a strategy used to reduce the impact of
selective browsing and increase the regeneration of desired plant species. On Anticosti Island
(Québec, Canada), selective browsing by white-tailed deer prevents the regeneration of
balsam fir – white birch forests leading to their conversion into white spruce forests. Large
deer exclosures were established in clear-cuts with patches of residual forest from 2001 to
2006 to assist in the natural regeneration of fir stands and to provide shelter and food
resources for deer. Our objective was to assess how deer use exclosures after the removal of
fences according to their spatial configuration, habitat composition, and time since opening.
We randomly distributed automatic cameras for periods of 14 days during summer in six
exclosures ranging from 3.1 to 11.2 km2 (n=25 cameras per exclosure) from which deer were
reduced for 10 to 12 years. We compared candidate occupancy models that included spatial
configuration and food resource variables while simultaneously controlling for variables
affecting detection probability. Our most supported model revealed that deer habitat use
increased by 19% when forage resources, represented by the cover of Cornus canadensis,
increased from 0 to 100%. None of the other variables (distance between the border of
exclosures and cameras, distance between forest patches and cameras, and forest patch sizes)
had an influence, suggesting that the use of regenerating forests by deer in summer after a
period of exclusion is mainly related to forage availability and therefore, any forest
management work that improves food production during summer should help maintain or
increase habitat use by deer.
12Introduction
The impact of chronic herbivory on plant communities has been extensively studied
over the past decades. Heavy selective browsing on plant species can alter natural
regeneration, plant communities and forest functions (Rooney 2001, Kuijper et al. 2010). For
instance, some large herbivores can affect seedlings’ regeneration of palatable species by
altering their growth (Frelich and Lorimer 1985, Kaien 2006). A study on browsed Scots
pine (Pinus sylvestris) by herbivores (goat (Capra hircus), ibex (Capra pyrenaica) and sheep
(Ovis aries)) showed that a constant browsing pressure on tree saplings can delay their
reproduction and growth by 12 years (Zamora et al. 2001).
Exclusion following harvesting can be used to prevent the negative effects of large
herbivores on forests. Some studies have shown the efficiency of this strategy to achieve
forest management objectives and to identify herbivore densities compatible with the natural
regeneration dynamics of managed forests (Kay and Bartos 2000, Horsley et al. 2003,
Persson et al. 2007). Exclosures are a management strategy that leads to exclusion of
consumers and that has been used to document the impact of large herbivores on individual
plant growth and survival (Kaien 2006, Hidding et al. 2012), changes in community
composition (Horsley et al. 2003, Tremblay et al. 2006), forest dynamics (Hidding et al.
2013, Nuttle et al. 2013, De Vrient et al. 2020) and ecosystem functions (Persson et al. 2005,
Vuorinen et al. 2020). Some studies have used this management strategy as a tool to enhance
biodiversity and conservation and yielded similar results: less browsing on selected species
and higher density of mature trees and saplings (Birhane et al. 2006, Leonardsson et al. 2015).
To understand how herbivores reclaim managed landscapes, we need to assess their
use of formally protected habitat. At a large scale, forest management inside exclosures can
create habitat fragmentation. Patches of regenerated forest in former exclosure can be isolated
from large areas of heavily browsed forest in the surrounding matrix. Large herbivores
require a minimum area of forest for foraging, resting, and social functions (Cargnelutti et al.
2002). Highly fragmented areas can negatively impact home range size of individuals
(Cargnelutti et al. 2002, Dechen Quinn et al. 2013). Thus, evaluating habitat use when the
13exclosure is removed is essential to assess the impact of exclosures on the behavior and
distribution of large herbivores, and hence the efficiency of this management action for long-
term forest dynamics.
On Anticosti Island, selective browsing induced by the high density of white-tailed
deer (Odocoileus virginianus) prevents the regeneration of native balsam fir forests leading
to their conversion into white spruce (Picea glauca) forests and parklands (Beaupré et al.
2005). Large management exclosures, in which clear cuts and patches of residual forest are
intermingled, have been established since 2001 to assist in the natural regeneration of fir
stands and to provide shelter and food resources for deer during winter. Deer densities in
exclosures were reduced though sport hunting. Several exclosures were considered a success
in terms of balsam fir regeneration after 10 to 12 years of reduced deer density. An evaluation
of the habitat use by deer is needed to provide insights about the efficiency of the
management strategy.
Our study objective was to evaluate summer habitat use by white-tailed deer in six
former management exclosures dismantled eight years ago. Based on our knowledge of
feeding habits and movements of white-tailed deer on Anticosti Island that revealed an
establishment of deer home ranges in habitats that present higher forage abundance (Massé
and Côté 2009, Coulombe et al. 2011), we hypothesized that habitat use would be determined
by forage resources in response to the need for deer to replenish body reserves for the winter
season. We predicted that deer would use more frequently areas with a higher basal area of
balsam fir and white birch as opposed to areas with a higher basal area of black and white
spruces. We also predicted that deer would more frequently use areas with a higher
percentage of ground forage. We also hypothesized that the integration of formal exclosures
into deer habitat use would be a function of their spatial configuration in terms of
regenerating cut blocks and mature forest patches and predicted that deer would use areas
closer to the exclosure border. In addition, we predicted that deer would use more frequently
areas closer to the largest forest cover patches as an escape cover. We used presence and
absence data obtained from camera traps to model habitat use while controlling for variables
14affecting detection probability. We discuss how the use of restored habitat by deer inform on
the sustainability of exclosures as deer and forest management method.
Materials and Methods
Study Area
Anticosti Island (49°30’N, 63°00’W) is located in the Gulf of St-Lawrence in Quebec,
Canada within the eastern balsam fir- white birch bioclimatic forest zone (Saucier et al.
2009). The vegetation is dominated by balsam fir, white and black spruces, as well as white
birch (Saucier et al. 2009). The long fire cycle makes spruce budworm (Choristoneura
fumiferana) outbreaks and windthrows the main disturbances leading to cyclic regeneration
of balsam fir stands (Baskerville 1975). The island is characterized by a maritime south-
boreal climate with fresh summers and long winters (Saucier et al. 2009). White-tailed deer
density on Anticosti Island is normally > 20 deer/km2 (Huot and Lebel 2012), but in 2017-
2019, it dropped to < 5 deer/km2 locally due to harsh winter conditions (Ayotte 2019). The
high deer density maintained over several decades induced heavy browsing pressure on
balsam fir, the main food resource of deer during winter (Lefort et al. 2007). Our study area
included six exclosures of 3.1 to 11.2 km2 built from 2000 to 2006 and dismantled from 2012
to 2017 (Figure 1).
15Figure 1. Location of the study area and attributes of the 6 exclosures under study on
Anticosti Island, Québec, Canada. In a) the specific information of every exclosure and their
position on the western part of the island, b) the location of Anticosti Island on the North
American continent and c) magnification of the Km 27 exclosure to show the camera
distribution.
Estimation of habitat use from camera traps
Camera traps can be used non-invasively to measure habitat use by animals
(O'Connell et al. 2011). The presence/absence data collected from cameras can be used within
an occupancy model framework that allows to assess the probability of occupancy while
controlling for the probability of detection. Because we measured occupancy in an open
system, occupancy estimates must be interpreted as the percentage of sites “used” by a
species rather than “occupied” (MacKenzie and Nichols 2004). To assess the effort needed
to obtain unbiased estimates of habitat use of white-tailed deer from camera traps (Hamel et
al. 2013), we performed a pilot study during all of July 2019 where we deployed 32 camera
traps programmed to take a picture every 5 minutes in the Lac Claude exclosure (Figure 1).
A minimum of 20 cameras programmed to take a picture every 5 minutes during 14 days was
sufficient to stabilize the precision of the estimates obtained from occupancy models
(Appendix 1).
16Vous pouvez aussi lire

















































