L'ADN, un polymère modèle - De la physique à la biologie
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
De la physique à la biologie
L’ADN, un polymère modèle
Traditionnellement, les cours de physique statistique modélisent un polymère sous la forme d’une
chaîne dont la configuration est celle d’une marche aléatoire (chaîne librement jointe). On montre
ainsi la nature entropique de l’élasticité d’un polymère. Depuis peu, ce modèle n’est plus
uniquement une vue de l’esprit : les expérimentateurs ont réussi à attraper un polymère par
chacune de ses extrémités et à mesurer la force nécessaire pour l’étirer. Il s’agit non pas de
n’importe quel polymère, mais de l’ADN. Ces expériences ne nous renseignent pas uniquement sur
l’élasticité entropique, elles révèlent également des transitions structurelles intrinsèques à la
molécule d’ADN, de sa forme classique en double hélice droite vers des structures plus exotiques.
epuis quelques années le comportement d’une molécule molécule d’ADN au microscope,
D l’ADN fait l’objet d’un inté-
rêt croissant de la part des
physiciens, non pas pour ses aspects
d’ADN, même longue de 20 à
30 µm, est proche de celui d’un po-
lymère idéal, dominé par l’entropie
chose jamais faite avec un quelcon-
que polymère synthétique. De fait,
les premières études sur l’ADN en
biologiques, mais comme modèle de la chaîne. La probabilité d’inter- tant que polymère utilisèrent cette
expérimental du comportement sta- action des maillons de l’ADN est en possibilité de visualiser une molé-
tistique d’un polymère idéal. effet si faible que les interactions di- cule pour étudier son comportement
Tout comme le nylon, le polysty- tes de volume exclu sont négligea- dynamique en électrophorèse ou en
rène ou tout autre polymère synthé- bles. écoulement, dans une solution, un
tique, l’ADN est en effet une Non seulement l’ADN est un gel ou un enchevêtrement d’autres
macromolécule très longue (l’ADN exemple de polymère très intéres- molécules d’ADN. Par la suite, la
d’un chromosome humain fait sant d’un point de vue théorique, possibilité de greffer l’ADN sur de
typiquement quelques centimètres) mais il possède, d’un point de vue nombreuses surfaces et le dévelop-
composé de maillons ou monomères expérimental, des qualités sans équi- pement de techniques de microma-
élémentaires : Adénine (A), Thy- valent parmi les polymères synthé- nipulation (pinces optiques, microfi-
mine (T), Guanine (G) et Cytosine tiques. Tout d’abord, il présente une bres, billes magnétiques, etc.) ont
(C), disposés régulièrement le long finesse de distribution de masse mo- permis les premières mesures
d’une chaîne sucre-phosphate. En léculaire inégalée, puisque l’ADN d’élasticité sur une seule molécule
fait, l’ADN est constitué de deux d’une souche particulière (virale, d’ADN, d’abord dans son régime
polymères complémentaires enrou- bactérienne, etc.) est constitué de entropique puis dans un régime
lés pour former la fameuse double- molécules rigoureusement identi- d’extension plastique jusqu’à la rup-
hélice de Watson-Crick et maintenus ques à la paire de base prés. Ainsi, ture de la molécule.
solidaires par des liaisons hydro- nous travaillons sur de l’ADN du On ne peut parler des propriétés
gène entre paires de bases (A-T, bactériophage k (un virus), qui pos- physiques de l’ADN sans mention-
G-C) (voir encadré 1). C’est cette sède exactement 48 502 paires de ner leur relation avec ses propriétés
structure en double hélice qui rend bases (pbs), soit une longueur de biologiques. En particulier, l’ADN
l’ADN beaucoup plus rigide qu’un 16.4 µm. Ensuite, il existe autour de étant une double hélice « main
polymère synthétique, 100 fois plus l’ADN une technologie chimique et droite », le degré de sur- ou de
environ que le polyéthylène. Ainsi, biochimique très développée. Il est sous-enroulement de la molécule est
la probabilité que deux parties ainsi possible de couper la molécule étroitement relié à sa structure tridi-
d’une molécule d’ADN soient en en des sites précis, de lui greffer un mensionnelle et à ses fonctions (ré-
contact est faible. En conséquence, grand nombre de groupements fonc- plication, transcription, régulation).
tionnels, de l’ancrer à un grand Cette hélicité est très importante,
nombre de surfaces de nature diffé- puisque la double hélice effectue un
rente, de raccorder ensemble plu- tour tous les 10,5 paires de bases.
sieurs molécules d’ADN, etc. Quelques tours de plus ou de moins
– Institut CURIE
– ESPCI Par ailleurs, un très grand nombre imposés à la molécule ont un effet
– ENS de molécules fluorescentes ont été déterminant sur la forme de celle-ci.
– Institut PASTEUR développées, qui s’intercalent dans Un changement de torsion de la mo-
– IBPC l’ADN. Cela permet de voir une lécule conduit principalement aux
111Encadré 1
LA MOLÉCULE D’ADN
L’acide désoxyribonucléique (ADN) est un polymère formé deux facteurs s’opposent à la répulsion électrostatique entre
d’un enchaînement de nucléotides constitués chacun d’un les groupements phosphates (porteurs de charge négative) et,
sucre (le désoxyribose), d’un groupement phosphate et d’une dans des conditions physiologiques, empêchent la séparation
base (Adénine : A, Thymine : T, Guanine : G, Cytosine : C) spontanée des brins.
(figure a). L’ADN existe normalement en forme de double La rotation entre deux paires de bases successives est
brin, qui permet aux bases de chaque brin de s’apparier en d’environ 36° et, ainsi, la double hélice fait un tour complet
formant des paires Adenine-Thymine et Guanine-Cytosine. environ toutes les dix paires de bases. Puisque les jonctions
Ainsi, les deux brins de la molécule portent des copies entre un sucre et les groupements phosphates adjacents se font
complémentaires du même message génétique. Les deux brins d’une manière asymétrique (aux sites C5′ et C3′), chaque brin
de l’ADN s’enroulent pour former la fameuse double hélice phosphodiester a un sens (conventionnellement pris dans la
découverte par Watson et Crick (figure b). direction 5′ → 3′. Au sein de la double hélice, les deux brins
La molécule de l’ADN est stabilisée à la fois par sont orientés d’une manière antiparallèle. La conformation
l’appariement des bases, à travers la formation de liaisons des brins phosphodiester est décrite par un total de sept
hydrogène, et par l’empilement des bases le long de l’axe de angles dièdres ~ a, b, c, d, e, f, v ! pour chaque nucléotide et
la molécule, à travers des interactions Van der Waals. Ces par le plissement de son sucre (figure a).
Figure a - Structure chimique d’un brin de l’ADN. Figure b - Conformation de double hélice de l’ADN dans sa conforma-
tion usuelle (ADN-B) (Rouge : le brin phosphodiester, bleu : Guanine,
jaune : Cytosine).
112De la physique à la biologie
entortillements de l’axe de la double RELAXATION DE LA MOLÉCULE D’ADN périence, très simple, consiste à ob-
hélice (writhe chez les Anglo- server et à quantifier par microsco-
Saxons) et aux structures plectonè- Pour étudier l’élasticité de pie de fluorescence la relaxation
mes ou solénoïdales. Les plectonè- l’ADN, une approche consiste à ob- d’une molécule unique, préalable-
mes sont ces structures torsadées, server directement par microscopie ment étirée par un écoulement dans
qui se forment bien souvent sur les de fluorescence la conformation un tube capillaire. On a ainsi une il-
cordons téléphoniques. Comme on adoptée par une molécule unique lustration très directe de « l’élasti-
peut facilement l’imaginer, leur (préalablement marquée à l’aide cité entropique », puisqu’on voit la
existence modifie l’élasticité du d’un intercalant fluorescent) sous molécule retrouver spontanément
polymère. l’action d’une sollicitation externe. une conformation compacte en « pe-
Ainsi, Perkins et al. ont récemment lote statistique » au sein d’un fluide
Dans cet article, nous voudrions étudié l’étirement d’une molécule en repos (voir figure 1), selon une
montrer l’apport important réalisé à d’ADN attachée par une de ses ex- loi qui est en bon accord avec la des-
l’heure actuelle par les techniques trémités à une bille de latex de di- cription théorique de cette élasticité.
fines de la physique dans l’étude mension micronique, cette dernière
des propriétés élastiques et plasti- étant déplacée à vitesse uniforme
ques de l’ADN. Nous décrirons dans un fluide à l’aide d’un laser fo- MESURE DIRECTE DE FORCE
successivement la dynamique de calisé formant une « pince opti- SUR UNE MOLÉCULE
relaxation d’une molécule après éti- que ». C’est dans ce cas la friction
rement et ses propriétés élastiques et du solvant sur l’ADN qui le dé- Les premières mesures des pro-
plastiques avec ou sans torsion. forme, et on a ainsi accès à ses priétés mécaniques d’un ADN dou-
propriétés hydrodynamiques. Ces ble brin unique dans le régime en-
auteurs ont également étudié la re- tropique ont été réalisées par le
laxation des molécules après l’arrêt groupe de C. Bustamante, à l’Uni-
de la sollicitation. Pour ce dernier versité d’Oregon. Cette expérience
type d’études, cependant, il est pré- utilisait une combinaison de forces
férable de s’affranchir de la pré- hydrodynamiques et magnétiques
sence des pinces optiques, qui peu- exercées sur une bille magnétique
vent perturber la relaxation, comme fixée a l’extrémité d’un brin d’ADN
cela a été fait très récemment à lui même relié par son autre extré-
l’Institut Curie : le principe de l’ex- mité à une lamelle de microscope.
Figure 1 - Relaxation d’une molécule d’ADN
initialement étirée dans un écoulement. La mo-
lécule est rendue visible grâce à une molécule
intercalante fluorescente. Durant ce régime
transitoire, la molécule retrouve sa configura- Figure 2 - Dispositif de mesure de force utilisant une micro-fibre. Celle-ci est une fibre optique (ver-
tion en pelote statistique (le temps entre deux ticale), les déplacements de son extrémité sont mesurés par un capteur de position. Une micro-pipette
clichés est de 3 secondes). (horizontale) permet d’attraper la bille et de tirer sur la molécule d’ADN.
113La bille est donc soumise à trois Cette technique présente une grande
forces : une force hydrodynamique sensibilité dans les faibles forces;
créée par un écoulement, une force par ailleurs, elle ne demande pas
magnétique créée en plaçant la bille d’établir un contact physique avec
dans un gradient de champ magnéti- la bille analysée, il est ainsi possible
que et la force due à l’élasticité de de faire tourner l’aimant, ce qui fait
l’ADN. Pour une valeur donnée de tourner la bille et permet d’enrouler
l’écoulement et du gradient de la molécule d’ADN sur elle-même.
champ magnétique, la bille adopte Évidemment, les attaches de la mo-
une position d’équilibre qui annule lécule d’ADN à la bille et au subs-
la résultante des trois forces aux- trat ont été spécialement préparées
quelles elle est soumise. En faisant pour empêcher la molécule de tour-
varier à la fois les composantes ma- ner sur elle-même.
gnétiques et hydrodynamiques, la
Figure 3 - Dispositif de mesure de force par Élasticité entropique d’une
bille adopte une succession de posi- mouvement brownien. L’aimant placé au-dessus
tions d’équilibre à partir desquelles molécule d’ADN
de l’échantillon permet de tirer et de faire tour-
on peut remonter à un diagramme ner la bille magnétique. Les fluctuations brow- S. Smith et al. ont mesuré l’élas-
niennes de la bille permettent de mesurer la ri-
force-extension. Ces expériences né- gidité du petit pendule magnétique que ticité d’une seule molécule d’ADN
cessitent un certain nombre de cali- constitue la bille attachée par la molécule double brin et simple brin. Leurs ré-
brations (pour chaque type de for- d’ADN.
sultats expérimentaux ont montré
ces) qui s’avèrent très délicates en que, si la fonction de Langevin (voir
particulier à cause des interactions encadré 2) décrivait bien les résul-
avec les parois. Néanmoins, elles tats à basse force, seul le modèle du
ont une extraordinaire sensibilité Mesure par mouvement brownien. ver était en excellent accord quelle
dans le domaine des très faibles for- A l’ENS, en collaboration avec que soit la force, comme on peut le
ces (forces inférieures au piconew- l’Institut Pasteur, nous avons déve- voir sur la figure 4.
ton). loppé une mesure de force
utilisant l’analyse du mouvement
Depuis, plusieurs groupes ont dé- brownien et permettant également
veloppé des techniques différentes de faire varier la superhélicité de la
pour améliorer la mesure de ces molécule. Comme dans les expé-
forces : riences réalisées par C. Bustamante,
l’échantillon est constitué d’une
Capteur utilisant une micro-fibre.
molécule d’ADN reliant un substrat
A l’Institut Curie a été développé un
plan à une micro-bille paramagnéti-
capteur de force basé sur une fibre
que de 2.8 µm de diamètre. Un
de verre très mince associée à un
aimant placé au-dessus de l’échan-
capteur de déplacement pour fabri-
tillon permet d’exercer une force sur
quer un nano-dynamomètre. Pour
la bille, qui se comporte ainsi
mesurer l’élasticité d’une molécule
comme un pendule (inversé). Cette
d’ADN, il ne reste plus qu’à accro-
force varie de façon importante (de
cher une extrémité de cette molé-
0,006 pN à 100 pN) lorsque l’ai- Figure 4 - Comparaison entre une courbe de
cule à la fibre et son autre extrémité force expérimentale réalisée sur une molécule
mant est éloigné ou rapproché
à une micro−bille. On attrape alors d’ADN et les prédictions théoriques du modèle
de l’échantillon. Pour déterminer marche aléatoire et du modèle du ver.
la bille avec une pipette et on tire
cette force, le mouvement brownien
sur la bille. En enregistrant les dé-
de la bille est analysé par un
placements de la fibre au cours de
traitement d’image en temps réel. ADN à forte extension
l’étirement, on obtient la courbe de
Le carré de l’amplitude des fluc-
force d’une molécule. La fibre doit 2
avoir une rigidité très faible (de tuations dx dans une direction Que se passe-t-il lorsque l’on tire
−3 −4 perpendiculaire à la force est inver- plus fort ? C’est à cette question que
10 à 10 N/m). Cela est réalisé
sement proportionnel à la raideur P. Cluzel et al. et Smith et al. ont
en réduisant par attaque chimique le
k = F/l du pendule magnétique apporté une réponse récemment :
diamètre de celle-ci à environ 1 kdx2 = 1 k T. En déterminant la
10 µm. La gamme de forces actuel- dans un premier temps, les liaisons
2 2 b
lement accessibles avec un tel dis- longueur l de ce pendule optique- atomiques se déforment de façon
positif expérimental est de 1 à ment il est possible de mesurer la élastique et la molécule s’allonge li-
1 000 pN. force F appliquée sur la molécule. néairement avec la force comme un
114De la physique à la biologie
Encadré 2
LE MODÈLE DE LA CHAÎNE LIBREMENT JOINTE
Une macromolécule comporte en général de très nombreux indépendants soumis à un champ. Comme dans le cas du
degrés de liberté conformationnels, liés aux potentiels de système paramagnétique, l’orientation des chaînons est
flexion et de rotation affectant les liaisons qui composent le contrecarrée par l’agitation thermique. L’allongement du
squelette et aux interactions avec le solvant. Cela rend une polymère en fonction de la force appliquée suit donc une
étude rigoureuse de l’élasticité a priori diffıcile. fonction de Langevin. L’énergie qu’il faut comparer à kb T
Heureusement, pour des macromolécules assez longues, les n’est autre que le travail de la force sur la longueur d’un
propriétés à grande échelle convergent vers un comportement maillon b soit Fb.
unique dit « universel », et la complexité des potentiels locaux
peut être oubliée au profit d’un petit nombre de paramètres
phénoménologiques. On peut appréhender l’essentiel de ces S D S
< RN > = L+ Fb = L coth Fb −
kb T
kb T
kb T Fb D (1)
propriétés à l’aide de quelques modèles simplifiés qui peuvent
En fait, l’approximation impliquant que chaque monomère
être décrits analytiquement par les méthodes de la physique
peut prendre n’importe quelle direction par rapport à ses
statistique.
voisins immédiats n’est pas toujours vérifiée pour les
Le modèle le plus simple est celui de la chaîne librement polymères réels. Kuhn a introduit une longueur effective de
jointe : il s’agit d’une succession de segments rigides maillon b qui est égale à deux fois la distance au-delà de
(baptisés « monomères ») reliés par leurs extrémités avec une laquelle les corrélations orientationelles disparaissent entre
entière liberté de rotation de l’un par rapport à l’autre et sans deux monomères (nommée aussi longueur de persistance n).
autre interaction. Si on numérote séquentiellement les points Pour décrire une chaîne réelle par le modèle de la chaîne
de liaison de 0 à N, la conformation, en l’absence de force librement jointe, il faut donc remplacer la longueur du
extérieure, peut être représentée par une « marche au monomère par la longueur de Kuhn b et leur nombre par
hasard » : le système ne met en jeu aucune enthalpie, et toutes Neff = L/b (où L est la longueur totale du polymère).
les conformations possibles, construites en donnant
successivement à chaque segment une orientation au hasard, Aux faibles forces on a un comportement de type hookéen.
sont équiprobables. Dans un bain thermique, une telle chaîne L’allongement est donné par < RN > = FNeff b/3kb T. Il tend
est cependant porteuse d’une élasticité de nature purement vers L pour une force F infinie.
entropique. Le nombre de configurations de la chaîne, porté Le modèle de la chaîne librement jointe n’est pas très réaliste
en fonction de la longueur effective séparant ses deux puisqu’il permet à deux maillons de se trouver au même
extrémités < RN > , présente une forme gaussienne centrée en endroit (absence des interactions dites de volume exclu) et il
zéro (c’est pourquoi on appelle souvent cette marche aléatoire ne prend pas en compte la courbure du polymère à une
non perturbée « chaîne gaussienne » ou « chaîne idéale »). échelle plus petite que celle de la longueur de persistance n.
Dès que l’on sépare les deux extrémités de la chaîne, Dans le cas de l’ADN, les interactions dites de volume exclu
l’entropie est réduite et il faut appliquer une force pour sont négligeables pour des molécules de quelques dizaines de
compenser cette perte d’entropie. kilo-bases. Il existe un second modèle dit de la « chaîne à
Le modèle de la chaîne librement jointe va nous permettre de longueur de persistance » ou « modèle du ver » (« wormlike
calculer l’allongement du polymère en fonction de la force chain » en anglais) qui est plus adapté : on représente la
exercée. Les forces opposées agissant sur chaque extrémité du macromolécule comme un fil de longueur fixe L présentant une
polymère sont appliquées sur chaque maillon de la chaîne, qui énergie de flexion en plus de son entropie. La résolution
se retrouve ainsi sous l’effet d’un couple cherchant à exacte de ce modèle est possible, le résultat est donné dans la
l’orienter dans la direction de la force. L’indépendance des figure 4. L’allure qualitative est la même que celle de la
maillons fait que la chaîne sous contrainte se comporte de la fonction de Langevin mais le comportement à haute force
même manière qu’une assemblée de moments magnétiques diffère de façon notable.
simple ressort. Mais pour une valeur eu pour une force inférieure à Cette transition de structure est
de ≈ 70 pN la molécule s’allonge 70 pN. Ce plateau révèle l’existence complètement abolie si on réalise la
brutalement de 60 %, comme on d’une transition de structure de même expérience en présence de
peut le voir sur la figure 5. Pour des l’ADN lorsque son extension dé- bromure d’éthidium, un agent inter-
forces encore plus grandes, la molé- passe la longueur habituellement re- calant qui par son insertion entre les
cule retrouve un comportement élas- connue comme correspondant à paires de bases déroule et allonge la
tique analogue à celui qu’elle avait l’extension normale (3,4 Å/bp). double hélice.
115Figure 6 - Augmentation de la longueur de la
double hélice de l’ADN par réduction de son
diamètre et par déroulement.
Figure 5 - Diagramme force-extension d’une
molécule d’ADN. Le plateau montre l’existence
d’une transition de structure. Une nouvelle
forme de l’ADN, qui correspond à une surex-
tension de la molécule, apparaît au début du champ de force paramétrisé spécifi-
plateau et se développe quand on parcourt
celui-ci, jusqu’à remplacer complètement la quement pour les acides nucléiques,
forme « classique » d’ADN-B. et mime la présence de l’eau et des
contre-ions à travers une fonction
Figure 7 - Trois façons d’étirer la double hé-
diélectrique et en tenant compte de lice de l’ADN : par les extrémités 3′, par les ex-
l’écrantage de la charge des groupe- trémités 5′ et par les deux extrémités d’un seul
MODÉLISATION DE L’ÉTIREMENT brin.
ments phosphates.
DE LA DOUBLE HÉLICE DE L’ADN
Ces études ont montré que
l’ADN se déforme d’une manière Ainsi, quand on étire l’ADN par
Pour comprendre ce qui se passe différente suivant les extrémités des les extrémités 3′, l’hélice s’allonge
quand on étire la double hélice de brins, qui sont étirées. Ce fait de- principalement par un déroulement
l’ADN, il faut d’abord tenir compte vient compréhensible si on consi- et les paires de bases se séparent le
de son enroulement. Du fait de la dère la figure 7 : l’étirement des long de l’axe de la molécule. Ce
rotation d’environ 36° entre les pai- deux extrémités 3′ (voir encadré 1), processus implique une perte d’em-
res de bases, la longueur du brin ou celui des deux extrémités 5′, pilement entre les bases, qui peut
phosphodiester qui relie deux bases auront tous les deux tendance à in- être partiellement compensée par la
successives (environ 7 Å) est à peu cliner les paires de bases, mais dans formation de liaisons hydrogène en-
près deux fois plus grande que la des directions opposées. Si, encore, tre les paires de bases successives.
distance qui sépare les paires de ba- on étire les deux extrémités d’un Pendant l’étirement, les bases res-
ses le long de l’axe de la double hé- seul brin (ou les deux extrémités tent à peu près perpendiculaires à
lice (3.4 Å). Ainsi, il est possible de des deux brins), il n’y aura pas de l’axe et la conformation finale res-
rallonger l’ADN par environ un fac- tendance vers l’inclinaison. La mo- semble à un ruban plat avec les pai-
teur deux, soit en réduisant son dia- délisation permet de raffiner cette res de bases au centre et les bords
mètre, soit en réduisant son enroule- vue schématique. Nous avons si- formés par les brins phosphodiester
ment (figure 6). Pour atteindre la mulé l’étirement des ADN polymé- (figure 8). Au-delà d’une extension
limite d’extension de l’ADN avant riques ayant des séquences simples de 2.1 fois la longueur initiale de la
cassure (environ 1.8 fois), il faut et répétitives, tels que le poly- molécule, les brins ne peuvent plus
donc invoquer un de ces deux mé- (dG-dC).poly(dG-dC) ou le poly- s’allonger et les liaisons hydrogène
canismes, couplé à des étirements (dA-dT).poly(dA-dT). Les résultats des paires de bases cèdent. Quand
relativement limités au sein de cha- démontrent, en premier lieu, que la on étire l’ADN par les deux extré-
que brin phosphodiester. rotation main droite de la double mités 5′, la déformation de la dou-
Pour mieux comprendre les chan- hélice implique qu’une inclinaison ble hélice induit principalement une
gements de conformation liés à positive (rotation dans le sens in- réduction de diamètre, obtenu à tra-
l’étirement de l’ADN, une étude par verse des aiguilles d’une montre vers une forte inclinaison négative
modélisation moléculaire a été ef- dans la figure 7) ouvre les sillons des bases. Cette déformation rap-
fectuée ; elle nous a permis de trou- alors qu’une inclinaison négative les proche les deux brins phosphodies-
ver la structure la plus stable de la referme. Deuxièmement, les incli- ter sur le côté petit sillon de la mo-
molécule en fonction de sa longueur naisons très positives sont contrain- lécule et expose les bases côté
et de sa séquence de bases. Cette tes par un couplage avec l’angle grand sillon (figure 8). Elle conduit
modélisation tient compte de l’éner- glycosidique dont le changement également à une perte graduelle des
gie interne du polymère grâce à un déstabilise les nucléotides C et T. liaisons hydrogène formant les pai-
116De la physique à la biologie
res de bases, et, comme dans le Du point de vue énergétique, l’étire- d’autres protéines qui sont capables
cas de l’étirement 3′3′, les paires ment 5′5′, qui permet de mieux d’étirer localement la double hélice
sont complètement rompues au-delà conserver l’empilement entre les d’ADN.
d’une extension d’environ 2.1. Fina- bases, semble être moins difficile
lement, quand on étire les deux ex- que l’étirement 3′3′. L’étirement
SUPERHÉLICITÉ DE L’ADN
trémités d’un brin (ou des deux 5′3′ conduit généralement aussi à
brins simultanément) la conforma- une conformation de type 5′5′, bien
tion finale ressemble soit à la que pour certaines séquences, par Que se passe-t-il si l’on tord une
conformation 5′5′, soit à la confor- exemple, le poly(dA-dC).poly(dG- molécule d’ADN ? Comme dans le
mation 3′3′, en fonction de la sé- dT), la forme 3′3′ soit préférée. cas du cordon téléphonique, on s’at-
quence de bases. Quelle que soit la tend à ce qu’il se forme des boucles
façon d’étirer l’ADN, la modélisa- Ces résultats peuvent être impor- entortillées et donc que la rigidité
tion indique que l’extension du po- tants pour comprendre certains phé- globale de la molécule augmente.
lymère est principalement associée nomènes de reconnaissance d’acides C’est précisément ce qui est ob-
au repositionnement relatif des brins nucléiques, l’allongement dû à la servé, comme on peut le voir sur la
de la double hélice plutôt qu’aux transition se révélant très voisin de figure 9, où l’on mesure l’allonge-
changements de conformation in- celui qu’induit une protéine de ment de la molécule soumise à une
terne. recombinaison recA ainsi que force constante en fonction de la
torsion appliquée r (= nombre de
tours appliqués/nombre de tours na-
turel de la double hélice). L’exten-
sion de molécule d’ADN est maxi-
male lorsque le sur-enroulement est
nul. A petite force (0,1 pN), la mo-
lécule se raccourcit de façon symé-
trique avec r. La sensibilité de la
molécule à la torsion est très impor-
tante puisque avec 100 tours soit
2 % de sur-enroulement, l’extension
de la molécule est réduite à 0. A
force moyenne (0,8 pN), une dissy-
métrie apparaît, la molécule ne
change plus de longueur lorsqu’elle
est sous-enroulée. A haute force
(8 pN), la molécule est quasiment
insensible à la torsion.
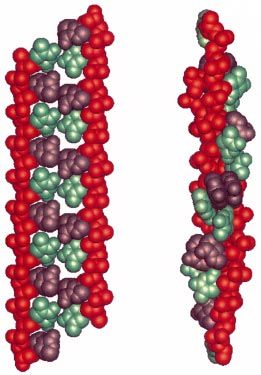
Figure 8 - La conformation de l’ADN étiré à 2.1 fois sa longueur normale : (a) étirement des deux Figure 9 - Extension relative d’une molécule
extrémités 3′, (b) étirement des deux extrémités 5′. (Séquence : poly(dA-dT).poly(dG-dC), rouge : le d’ADN en fonction du sur-enroulement ~ r ! à
brin phosphodiester, violet : Adenine, vert : Ihymine). force constante (dans 10mM PB).
117Comme notre expérience quoti- faibles peut modifier localement sa récente de Yin et al. (1995) : dans
dienne avec des cordons de télé- structure pour réduire la contrainte cette expérience de transcription,
phone nous le suggère, une molé- de torsion. Ces expériences sont par l’ARN polymérase est absorbé sur
cule d’ADN sur- (ou sous) enroulée ailleurs en très bon accord avec une une surface et l’ADN en train d’être
est plus rigide que relaxée. Ici aussi théorie récente due à J. Marko et transcrit a une extrémité accrochée à
la sensibilité de la molécule à la tor- E. Siggia des universités de Rock- une bille de polystyrène. A l’aide
sion est très importante puisque feller et de Cornell. Cette théorie d’un piège optique, on peut attraper
avec seulement 50 tours soit 1 % de décrit l’ADN comme un fil, caracté- et bloquer la bille. L’ADN est ainsi
sur-enroulement, la rigidité effective risé par des modules de flexion et tenu « par la queue ». On peut ainsi
de la molécule est augmentée de de torsion, en équilibre thermodyna- mesurer la tension de « tirage » né-
≈ 50 %. La dissymétrie entre r posi- mique avec la solution environ- cessaire pour bloquer la polymérisa-
tif et négatif apparaît ici lorsque la nante. Les transitions brusques ob-
tion et avoir une idée de la force dé-
force de traction atteint 0.4 pN : servées dans nos expériences sont
veloppée par la polymérase au cours
l’ADN sous-enroulé se rallonge bien décrites par les différences
brusquement. Cet allongement s’ef- d’énergie entre la forme B de de son déplacement le long de
fectue toujours à une force de l’ADN et ses autres structures. Nous l’ADN.
0.4 pN, indépendamment du degré disposons donc d’un système expé-
d’enroulement de la molécule. Cette rimental dont la compréhension
brusque transition reflète sans doute théorique va nous permettre d’abor-
la formation de régions (bulles) der des problèmes plus complexes, POUR EN SAVOIR PLUS
d’ADN dénaturé dans la structure tels que l’interaction de l’ADN avec
en double hélice standard (dite certaines protéines et en particulier
forme B). Ces bulles absorbent le avec les gyrases responsables de la Smith (S.B.), Finzi (L.) et Busta-
déficit d’enroulement et permettent relaxation des contraintes torsion- mante (C.), Direct mechanical measu-
donc la relaxation torsionnelle du nelles dans l’ADN lors de la trans- rements of the elasticity of single
reste de la molécule. A des forces cription et de la réplication de la DNA molecules by using magnetic
élevées (au-delà du piconewton), molécule. beads. Science, 258, 1122–1126, 1992.
l’extension de la molécule est simi- L’ensemble des résultats présen- Cluzel (P.), Lebrun (A.), Heller (C.),
laire à celle d’un ADN relaxé. tés ici montre le rôle potentiel Lavery (R.), Viovy (J.L.), Chatenay
On observe un comportement considérable que peuvent jouer les (D.) et Caron (F.), DNA : An extensi-
qualitativement similaire pour une propriétés plastiques de l’ADN. Il y ble molecule Science ; 271, 792-794,
molécule d’ADN sur-enroulée, mais a fort à parier que dans les dix an- (1996).
à une force plus élevée. La molé- nées à venir, des percées importan- Strick (T.R.), Allemand (J.–F.), Ben-
cule d’ADN étant chirale, on s’at- tes vont être faites dans ce domaine. simon (D.), Bensimon (A.) et Cro-
tend bien à des différences quantita- Un aspect sera très certainement de quette (V.), The Elasticity of a Single
tives suivant qu’on l’enroule dans le bien comprendre le rôle physique de Supercoiled DNA Molecule. Science
sens de la double hélice (sur- la superhélicité dans des phénomè- in press.
enroule) ou en sens inverse. Ainsi, à nes cellulaires aussi importants que
Smith (S.B.), Cui (Y.) et Bustamante
partir d’une force critique d’environ l’initiation de la transcription ou le
(C.), Overstretching B-DNA : The
3 pN, la molécule sur-enroulée se démarrage de la réplication (forma-
Elastic Response of Individual Double
rallonge brusquement. Comme pré- tion d’œil de réplication ou d’insta-
Stranded and Single Stranded DNA
cédemment cette transition reflète la bilité dans la double hélice) mais
Molecules. Science, 271, 795–799,
présence de bulles qui absorbent aussi le rôle de cette extensibilité
1996.
l’excès de sur-enroulement, peut dans la formation de structure
être par la formation d’ADN-D (une d’ADN plus complexe que la dou- Marko (J.F.) et Siggia (E.D.), Phys.
forme plus enroulée que la forme B ble hélice. Rev. E52, 2912 (1995).
naturelle) ou par dénaturation par- Un autre aspect important dans la Yin (H.) et al., Science, 270, 1653
tielle de la molécule. mécanique cellulaire réside aussi (1995).
Ces expériences montrent qu’une dans les interactions entre protéines Lebrun (A.) et Lavery (R.), Nucleic
molécule d’ADN enroulée et sou- et ADN. Quelques belles expérien- Acid Res. Modelling extreme stret-
mise à des tractions relativement ces ont déjà été faites comme celle ching of DNA 24, 2260-2267 (1996).
Article proposé par : D. Chatenay, PH. Cluzel, Ch. Heller, J.L. Viovy, J-F. Alle-
mand, D. Bensimon, F. Caron, V. Croquette, T. Strick, A. Bensimon, R. Lavery et
A. Lebrun
118Vous pouvez aussi lire

















































