La sélection d'habitat dépendante de la densité varie entre les mâles et les femelles chez l'éléphant d'Afrique (Loxodonta africana) - Corpus UL
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
La sélection d'habitat dépendante de la densité varie
entre les mâles et les femelles chez l’éléphant d’Afrique
(Loxodonta africana)
Mémoire
Audrey-Jade Bérubé
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Audrey-Jade Bérubé, 2021
Résumé
L’augmentation des activités anthropiques complexifie la conservation des grands
mammifères et de leur environnement dans les aires protégées. Ce constat est
particulièrement vrai lorsque l’on s’intéresse à la gestion des herbivores de grande taille dans
de petites réserves. En effet, comme les réserves permettent généralement de diminuer les
conflits humains-animaux, la population de ces derniers augmente, ce qui peut entraîner des
conséquences néfastes sur la qualité de leur habitat, conséquences amplifiées dans les aires
confinées et de petite superficie. La sélection d’habitat par un animal est une réponse
comportementale complexe à plusieurs variables environnementales et qui peut différer entre
les sexes et selon la densité de la population. Peu d’études ont toutefois démontré que les
mâles et les femelles diffèrent dans leur patron de sélection d’habitat dépendante de la
densité. Mon projet démontre que la réaction de l’éléphant africain (Loxondonta africana)
aux variables anthropiques de l’habitat varie avec la taille de la population et que ces
ajustements densité-dépendant diffèrent entre les sexes et les saisons. L’analyse de sélection
d’habitat a été réalisée sur 11 années en équipant de colliers GPS des éléphants de la réserve
faunique d’Ithala en Afrique du Sud. Mes analyses ont révélé que les éléphants d’Ithala
sélectionnaient les endroits près de l’eau et évitaient les secteurs avec une pente abrupte. Ils
évitaient également les secteurs dominés par la végétation herbacée alors qu’ils
sélectionnaient les endroits présentant une plus grande proportion d’espèces ligneuses. Mon
étude a démontré que, sans discriminer entre les sexes ni considérer l’effet de la densité
conspécifique, nous aurions conclue simplement que les éléphants sélectionnaient les
variables anthropiques de l’habitat. Nos modèles complexes permettent cependant de
démontrer que les mâles sélectionnaient davantage les infrastructures humaines, les endroits
à proximité des routes et les zones limitrophes de la réserve comparativement aux femelles.
Ces dernières évitaient d’ailleurs les infrastructures en tout temps et les endroits à proximité
des clôtures en saison humide. De plus, avec une augmentation de la densité d’éléphants, les
mâles augmentaient davantage que les femelles leur sélection de ces trois variables
anthropiques. Mon étude démontre que les femelles et les mâles éléphants n’utilisent pas
l’habitat de manière identique et qui plus est, ne réagissent pas de la même façon à une
augmentation de population. Considérer l’effet du sexe et de la densité sur la sélection
d’habitat permettrait de réduire le risque de conclusions erronées sur les interactions entre un
ii
animal et son environnement et donc potentiellement d’établir des décisions de gestion plus
efficaces et ciblées, par exemple la relocalisation de certains mâles dont leur sélection
d’habitat augmente les risques de conflits avec les humains.
iii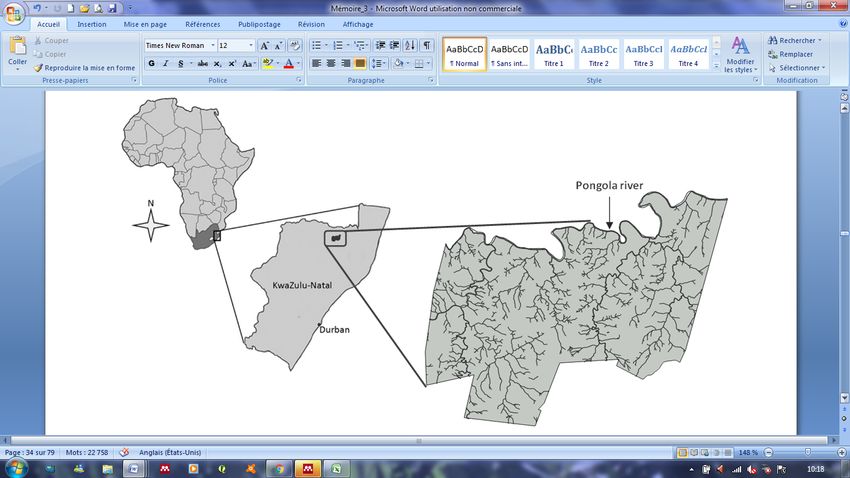
Abstract
The increase in anthropogenic activities has negative effects on the conservation of
large mammals and their environment in protected areas, and the management of large
herbivores in small reserves is particularly challenging. Indeed, the reduction of human-
animal conflicts in reserves often generally results in the increase of large herbivore
populations, with significant impacts on habitat quality and those impacts are generally
exacerbated in small and confined reserves. Habitat selection is recognized as a complex
process involving a response to multiple environmental features that can vary between sexes
and with population density. Yet there is still limited empirical evidence of males and females
displaying different patterns of density-dependent habitat selection. My project demonstrates
that the spatial response of the African elephant (Loxodonta africana) to human-related
habitat features varied with population size, and that density-dependent adjustments differed
between sexes and seasons. Habitat selection analysis was based on an 11-year monitoring
of GPS-collared elephants in Ithala Game reserve, South Africa. Globally, Ithala elephants
selected areas with abundant woody vegetation and close to water and tended to avoid areas
with steep slopes. Our analyses also revealed that, without discriminating between sexes and
accounting for conspecific density, the conclusion would have been simply that elephants
typically select human-related habitat features. We show, however, that males had a stronger
selection for both infrastructures and areas close to roads and fences compared to females
which avoided infrastructures all year long and areas close to fences in the wet season. With
an increase in population density, males also increased more strongly than females their
selection of these three human-related habitat features. My study demonstrates that females
and males differ in their habitat selection, a trend that could be exacerbate by an increase in
population density. Therefore, considering variations in habitat selection between sexes and
with conspecific density can help prevent faulty conclusions on the interaction between an
animal and its environment, and help develop more effective management tools, for example
relocation of certain males whose habitat selection might increase human-wildlife conflicts.
iv
Table des matières
Résumé .............................................................................................................................. ii
Abstract ............................................................................................................................ iv
Table des matières ............................................................................................................. v
Liste des tableaux ............................................................................................................ vii
Liste des figures .............................................................................................................. viii
Avant-propos .................................................................................................................. xiii
Introduction ....................................................................................................................... 1
Expansion des populations humaines et perte d’habitat .................................................. 1
Conservation des grands mammifères ............................................................................ 1
Sélection d’habitat : environnement ............................................................................... 3
Sélection d’habitat : saison ............................................................................................. 3
Sélection d’habitat : sexe................................................................................................ 4
Sélection d’habitat : densité............................................................................................ 5
Conservation des éléphants ............................................................................................ 7
Sélection d’habitat par l’éléphant ................................................................................... 8
Contexte de l’étude ...................................................................................................... 10
Site d’étude .................................................................................................................. 12
Objectifs de l’étude et approche méthodologique ......................................................... 13
Chapitre 1. Density-dependent habitat selection varies between male and female African
elephants .......................................................................................................................... 14
Résumé ........................................................................................................................ 15
Abstract ....................................................................................................................... 16
1.1 Introduction............................................................................................................ 17
1.2 Methods ................................................................................................................. 20
1.2.1 Study area ....................................................................................................... 20
1.2.2 Elephant locations ........................................................................................... 21
1.2.3 Habitat variables ............................................................................................. 21
1.2.4 Statistical analyses .......................................................................................... 22
1.2.4.1 Resource selection function ...................................................................... 22
1.2.4.2 Model selection and cross-validation ........................................................ 23
1.3 Results ................................................................................................................... 23
v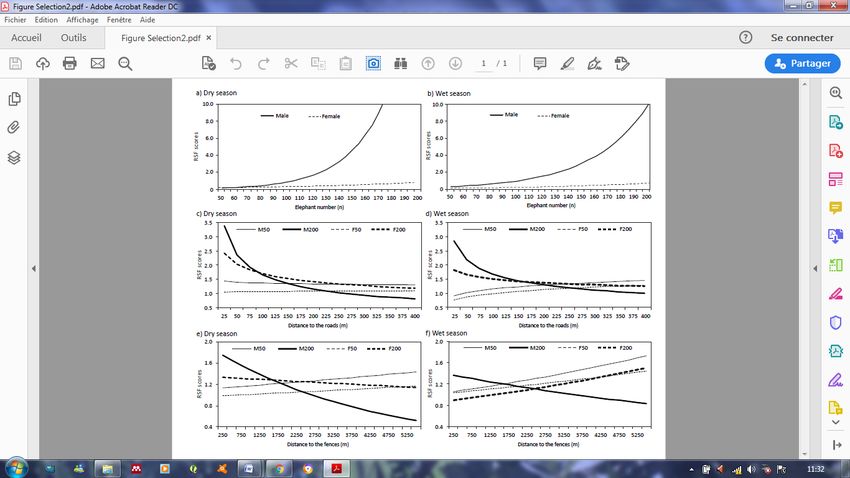
1.3.1. Simple models: effect of land cover types, slope and rivers on habitat selection
................................................................................................................................. 23
1.3.2. Complex models: effects of sex and density on the selection of human-related
habitat features ........................................................................................................ 24
1.4 Discussion .............................................................................................................. 27
1.5 Acknowledgements ................................................................................................ 31
Conclusion ....................................................................................................................... 32
Synthèse des résultats ................................................................................................... 32
Implication des résultats d’utilisation de l’habitat ......................................................... 33
Limites de l’étude ........................................................................................................ 34
Perspectives ................................................................................................................. 35
Bibliographie ................................................................................................................... 37
Annexe I. Distribution des données de colliers GPS dans le temps pour chaque individu.. 46
Annexe II. Représentation des différentes variables d’habitat de la réserve faunique
d’Ithala. ........................................................................................................................... 47
Annexe III. Ajustement d'un modèle exponentiel de croissance de la population aux
estimations de population disponibles pour la population d’éléphants dans la réserve
faunique d’Ithala en Afrique du Sud................................................................................. 50
Annexe IV. Coefficients de sélection (β) et intervalles de confiance (95%) de modèles de
sélection d’habitat par l’éléphant dans la réserve faunique d’Ithala, Afrique du Sud. ........ 51
vi
Liste des tableaux
Table 1. Parameter estimates (β) of resource selection functions by elephants in wet and
dry seasons, in Ithala Game Reserve, KwaZulu-Natal, South Africa. Unlike the simple
model, the complex model accounted for the interaction between Sex, Elephant density, and
human-related habitat features. Averaged (n = 100) Spearman rank correlation coefficient
(r̅ s) from 5-fold cross validation and their standard deviations (SD) are also presented. 95%
confidence intervals are provided in Annex IV. ................................................................ 25
Table 2. AICc and BIC model selection over two candidate glmer models of habitat
selection by elephants, A) in wet season and B) in dry season, in Ithala Game Reserve,
KwaZulu-Natal, South Africa. The model Simple contains: Land covers, Distance to road,
river and fence and Slope, whereas the model Complex contains all of these variables plus a
triple interaction between sex, elephant density and the human variables (Infrastructures,
Distance to road, Distance to fence). The number of parameters (K), the Akaike’s
information criterion corrected for small simple size (AICc), the difference in AICc from
the best model (∆AICc), the AIC model probability (weight) (wAICc), the Bayesian
information criterion (BIC), the difference in BIC from the best model (∆BIC), the BIC
model probability (weight) (wBIC) and the log-likelihood (LL) are presented for each
model. A) In wet season B) In dry season ........................................................................ 27
Table 3. Parameter estimates (β) and their 95% confidence intervals (CL) of resource
selection functions by elephants in Ithala Game Reserve, KwaZulu-Natal, South Africa. A
simple model was estimated for each season, together with a complex model that also
included Sex and Elephant number and their interactions with human infrastructures.
Averaged (n = 100) Spearman rank correlation coefficients (r̅ s) from k-fold cross validation
and their standard deviations (SD) are also presented for each model. Parameter estimates
with the 95% confidence interval excluding 0 are in bold. ................................................ 51
vii
Liste des figures
Figure 1. Geographic location of Ithala Game Reserve, in KwaZulu-Natal, South Africa
and representation of its water network including the Pongola River, at the northern
boundary of the reserve. ................................................................................................... 21
Figure 2. Effect of sex and elephant density on the response of elephants to three
anthropogenic habitat features in dry and wet seasons in Ithala Game Reserve, South
Africa. RSF scores were calculated from the models presented in Table 1. Female selection
is represented by a dotted line whereas male selection is represented by a full line. A)
Relative probability of selection of infrastructures in dry season and B) in wet season; C)
Relative probability of selection with respect to roads in dry season and D) wet season for
males with 50 (M50) and 200 (M200) conspecifics and for females with 50 (F50) and 200
(F200) conspecifics; E) Relative probability of selection of fences in dry season and F) wet
season for males and females also at two population densities. ......................................... 26
Figure 3. Distribution of GPS collar data over time for each elephant studied in Ithala
Game Reserve, KwaZulu-Natal, South Africa. Female data distribution is represented by
red dotted lines whereas male data distribution is represented by blue full lines. .............. 46
Figure 4. A) Distribution of the 26 land cover types and B) Distribution of the 9 land cover
types derived from the detailed vegetation map in A) in Ithala Game Reserve, KwaZulu-
Natal, South Africa. ......................................................................................................... 48
Figure 5. Digital elevation model (20 m) in Ithala Game Reserve, KwaZulu-Natal, South
Africa. ............................................................................................................................. 49
Figure 6. Roads and topography in Ithala Game Reserve, KwaZulu-Natal, South Africa. 49
Figure 7. Exponential population growth model of the elephant population in Ithala Game
Reserve, South Africa. Circles represent observed population estimates, whereas triangles
are predictions from the adjusted model. .......................................................................... 50
viii
« Tombe sept fois, relève-toi huit fois. »
-Proverbe japonais
ixRemerciements
Plusieurs personnes méritent des remerciements pour leur soutien tout au long de cette
drôle de montagne russe qu’est la maîtrise. Tout d’abord, un merci spécial à mon directeur
de recherche Stéphane Boudreau de m’avoir donné la très grande opportunité de réaliser un
projet sur les éléphants et de récolter des données directement en Afrique du Sud et d’avoir
su être toujours disponible pour répondre à mes petites questions ici et là et ce, même avec
son nouveau poste de directeur du département par intérim. Merci également à Adrian
Shrader, mon superviseur en Afrique du Sud, qui m’a accueilli dans son université, a su
récolter l’information nécessaire pour me concocter une nouvelle question de recherche
lorsque la dernière est tombée à l’eau (eh oui, un mois après mon arrivée), a encadré mon
terrain et a même fait 10 h de route pour venir me porter un nouveau véhicule sur le terrain
lorsque l’autre s’est brisé. Il faut spécifier que le second a failli nous lâcher également….
Hahaha ! Un gros merci à Daniel Fortin pour sa participation à mon comité d’encadrement,
mais également pour son rôle de co-directeur, rôle qu’il a endossé en milieu de parcours en
raison de son aide indispensable dont il faisait déjà preuve au niveau des analyses. Je tiens
également à remercier David Ward, professeur à l’Université de Kent, qui m’a donné de
précieux conseils sur ma récolte de données et sur mes analyses. Finalement, merci à Jean-
Pierre Tremblay qui a accepté de siéger sur mon comité d’encadrement et qui a évalué ma
présentation de projet et oups, oublié mon séminaire de maîtrise. Sans rancune, tu dois quand
même passer à travers mon mémoire J
Mon terrain aurait été très différent sans l’aide ô combien souhaitée de Douglas
Makin, ancien étudiant au doctorat d’Adrian Shrader, qui a accepté à ma plus grande joie
d’être mon assistant de terrain pour les 3 semaines passées à la réserve faunique d’Ithala. Son
aide a été plus qu’appréciée et je ne me serais jamais vu réaliser ce terrain sans lui. Il a su
endurer la petite québécoise indécise et hors de sa zone de confort que j’étais. Merci
également à Rickert Van Der Westhuizen, l’écologiste d’Ezemvelo KZN Wildlife d’avoir su
m’encadrer sur le terrain et d’avoir répondu à mes nombreuses questions sur les données
disponibles à la réserve. Merci à Pete Ruinard, gestionnaire de la conservation à la réserve
faunique d’Ithala ainsi qu’aux autorités de la réserve de m’avoir permis de réaliser ce projet.
xMerci également aux rangers qui ont assuré notre sécurité. Je tiens également à remercier
Jane et Angus Flockhart, les gens qui m’ont hébergé en Afrique du Sud, pour leur accueil
plus que chaleureux et leur grande générosité. Merci à Ivana Cinková d’avoir été ma « seule
amie » sud-africaines, pas sud-africaine du tout !
Je souhaite également offrir mes remerciements aux membres passés et présents de
mon laboratoire pour leur soutien moral et leurs conseils. Un merci particulier à Catherine
Chagnon, Claudia Beaupré et Anna Gaspard pour avoir fait de notre bureau un endroit
agréable et sympathique à travailler ! Des questions de syntaxes aux conversations de fi-filles
(oh ouiiii !!), une chance qu’elles étaient là pour rendre le laboratoire plus vivant. Merci à
tous les gens qui m’ont aidé de près et de loin dans ma maîtrise que ce soit pour des questions
de statistiques, de boucles d’extraction dans R ou pour les fameuses projections de mes
shapefiles qui m’ont données tout un mal de tête. Pour n’en nommer que quelques-uns, merci
à Marie-Caroline Prima, Ricardo Simon, Sabrina Plante et Yannick Seyer pour leur précieuse
aide et leur disponibilité. Merci également aux responsables de travaux pratiques et de
recherche du département des sciences géomatiques Louis-Étienne Guimond et Suzie
Larrivée, à Marie-Andrée Drouin, technicienne en géomatique à la bibliothèque de
l’Université Laval et à François Girard, professeur à l’UdeM, pour avoir tenté de trouver le
problème avec mes projections. Merci à Laurence Desbois-Bédard du Service de
Consultation Statistique pour avoir révisé et peaufiné mon script R.
Sans oublier ma famille et mes amis qui m’ont apporté support, conseils et réconfort
dans les bons comme les mauvais moments qui ont parsemé mon parcours d’études graduées.
Que ce soit au niveau académique qu’au niveau personnel, ils ont écouté sans broncher (ou
presque...) mes découragements et plaintes et ont toujours essayé de me remonter le moral et
de me pousser à rester positive et à avancer. Un merci particulier à mes parents qui ont vécu
les hauts et les bas avec moi, à mes Biogirls qui m’ont permis de me changer les idées et
m’ont épaulée et aux irréductibles gens des 5 à 7 de biologie qui ont mis un peu de vie dans
le département de biologie. Petit clin d’œil également aux membres du laboratoire Gauthier
qui m’ont accueilli à plusieurs reprises lorsque mes collègues de laboratoire étaient parties
trotter dans le nord en me laissant toute seule à mes mammifères du sud.
xiFinalement, il est important de souligner la contribution de plusieurs partenaires
financiers dans la réalisation de ce projet de maîtrise tel que le Conseil de recherches en
sciences naturelles et en génie du Canada (CRSNG) et le Fonds de recherche du Québec –
Nature et technologies (FRQNT) qui ont contribué à la majeure partie de mon salaire de
maîtrise. Le supplément pour études à l’étranger Micheal-Smith du CRSNG, la bourse
« Stage hors Québec » du Bureau international ainsi que la bourse de « Les Offices jeunesse
internationaux du Québec » (LOJIQ) m’ont permis de réaliser mon terrain en Afrique du Sud.
Un merci également à Stéphane Boudreau d’avoir contribué financièrement aux sessions
supplémentaires.
xiiAvant-propos
Mon projet a comme objectif d’évaluer la sélection d’habitat par l’éléphant au sein de
la réserve faunique d’Ithala en Afrique du Sud. À l’aide de colliers GPS et de modèles de
sélection des ressources, nous avons démontré qu’omettre de considérer les impacts du sexe
des individus et de la densité de la population sur la sélection des variables anthropiques peut
entraîner des conclusions erronées sur la sélection des ressources. La première section de ce
mémoire correspond à une introduction générale comprenant les notions théoriques et le
contexte de cette étude. Le chapitre 1, le cœur de ce mémoire, est un article scientifique
rédigé en anglais présentant les résultats de cette étude. Finalement, la dernière section de
mon mémoire est une conclusion revenant sur les résultats obtenus et sur les répercussions
potentielles de ce projet.
L’article scientifique sera éventuellement soumis à une revue scientifique (à être
déterminée ultérieurement). L’ordre des auteurs de l’article ainsi que leur rôle est présenté
ci-dessous. Stéphane Boudreau, mon directeur de recherche, a participé à la conception du
projet avec Adrian Shrader et Dave Ward et m’a accompagné à toutes les étapes de celui-ci.
Daniel Fortin, mon co-directeur, s’est joint en cours de parcours et m’a guidé dans les
analyses statistiques, la rédaction et la révision du mémoire. Les 4 co-auteurs mentionnés
précédemment vont tous participer à la révision de mon article scientifique. Pour ma part, je
suis premier auteur de l’article et j’ai participé à ce projet du début à la fin, soit de la
conception à la révision.
Liste des auteurs de l’article en ordre :
(1) Audrey-Jade Bérubé : étudiante à la maîtrise au Département de biologie de l’Université
Laval ;
(2) Daniel Fortin : codirecteur de recherche et professeur titulaire au Département de
biologie de l’Université Laval;
(3) Adrian Shrader : codirecteur de recherche et professeur titulaire au Département de
zoologie et entomologie de l’Université de Pretoria, Afrique du Sud;
(4) David Ward : professeur titulaire au Département des sciences biologiques de
l’Université de Kent, États-Unis;
(5) Stéphane Boudreau : directeur de recherche et professeur titulaire au Département de
biologie de l’Université Laval.
xiiiIntroduction
Expansion des populations humaines et perte d’habitat
La biodiversité des écosystèmes terrestres est grandement menacée par la perte et la
fragmentation de l’habitat (Wiens 1996, Primack 2001, Kerr et Deguise 2004), deux
processus associés à l’augmentation de la population humaine mondiale (Costa et al. 2005).
On estimerait à plus de 32 000 le nombre d’espèces menacées d’extinction, soit 27% des
espèces étudiées mondialement. De ce nombre, on retrouve près du quart des espèces de
mammifères, 41% des amphibiens et 14% des oiseaux (Brashares 2003, Hoffmann et al.
2010, Imre et Derbowka 2011, IUCN 2020). Les activités humaines, telles que la conversion
des environnements naturels en terres agricoles ou l’étalement urbain (Forman et al. 2003)
amènent une perte de biodiversité qui peut menacer l’intégrité écologique des écosystèmes
(Canale et al. 2012; Galetti et Dirzo 2013; Dirzo et al. 2014). Le taux d’extinction actuel est
d’ailleurs ca. 1000 fois plus élevé que le taux inféré à partir des registres fossiles, i.e., avant
que l’humain n’ait un impact majeur sur celui-ci (De Vos et al. 2014). Par conséquent,
l’humain est maintenant devenu l’un des principaux agents responsables de l’extinction des
espèces.
Les activités anthropiques entraînent une diminution de la taille des fragments
d’habitat résiduels et contribuent à augmenter la distance les séparant (Fahrig 2003), deux
phénomènes se traduisant par une perte de biodiversité selon la théorie de la biogéographie
insulaire (MacArthur et Wilson 1963,1967). De plus, la taille réduite de certains fragments
n’est pas suffisante pour assurer la réalisation du cycle vital de nombreuses espèces alors
que des distances accrues peuvent nuire aux espèces nécessitant de grandes aires pour leurs
besoins, comme c’est le cas de nombreuses espèces de grands mammifères herbivores ou
carnivores (Chiarello 2000, Cardillo et al. 2004, Bartlett et al. 2016).
Conservation des grands mammifères
Avec l’augmentation des activités anthropiques, la conservation des mammifères de
grande taille représente un véritable casse-tête (Kerley et al. 2003). En effet, la conservation
1des espèces s'effectue principalement dans des aires protégées aux multiples objectifs :
protection des habitats naturels, restauration des habitats dégradés, conservation des espèces
animales, réduction des conflits humains-animaux, développement de l’industrie
écotouristique, etc. (Garai et al. 2004). Cependant, en raison des différentes activités
humaines, les endroits propices à la mise en place de nouvelles aires de conservation
couvrent de faibles superficies, ce qui limite le nombre et le type d'espèces pouvant y être
conservées efficacement. Morcatty et al. (2013) ont d’ailleurs observé que la diversité de
mammifères était proportionnelle à la superficie des aires protégées de la région du
Quadrilatero Ferrifero, au sud-est du Brésil. De plus, aucune des aires protégées répertoriées
ne possédait la superficie minimale requise pour converser l’ensemble des espèces menacées
ou vulnérables de mammifères de moyenne et grande taille recensées dans ces aires
protégées. Finalement, les petites réserves seraient moins appropriées pour prévenir
l’extinction locale d'espèces, particulièrement si d'autres vecteurs de perturbations
(braconnage, maladies) menacent de réduire les populations (Di Minin et al. 2013).
La conservation des herbivores de grande taille dans de petites réserves pose donc
des défis particuliers en gestion des ressources fauniques. À l’inverse de l’extinction, une
protection accrue face aux activités anthropiques illégales (ex. braconnage), l'absence de
carnivores supérieurs (nécessitant de plus grandes superficies; Mattson et al. 1996) et
l’érection de clôtures pour limiter les conflits humains-animaux (Berger et Cunningham
1994, Campbell et Hofel 1995), favorisent la croissance démographique des grands
herbivores dans les petites aires protégées (Owen-Smith 1983). Confinés à l’intérieur de ces
petites aires protégées et augmentant en nombre, ces herbivores peuvent dégrader leur
habitat (ex. O’Connor et al. 2007, Guldemond et van Aarde 2008) en induisant une pression
accrue sur les espèces végétales consommées (de Beer et van Aarde 2008, Shannon et al.
2006a). Les clôtures ont donc un effet amplificateur sur l’impact des herbivores sur la
végétation (Guldemond et van Aarde 2008). En plus de menacer l’intégrité écologique des
petites aires protégées (ex. Wiseman et al. 2004), cette altération exacerbée de l’habitat peut
par la suite se répercuter sur la population de l’herbivore.
2Sélection d’habitat : environnement
Évaluer la sélection d’habitat d’un animal nécessite une bonne compréhension des
différentes variables pouvant influencer ses choix. Plusieurs études se sont attardées aux
variables environnementales façonnant la répartition des animaux au sein d’une aire. Parmi
les variables ayant un impact sur l’utilisation, on retrouve le type de végétation
(Forchhammer 1995; Bugalho et al. 2001; Fortin et al. 2009; Bjørneraas et al. 2011; Bliss et
Weckerly 2016), la disponibilité (Jiang et al. 2010; Laurian et al. 2012, Massé et Côté 2012)
et la qualité de la nourriture (Herfindal et al. 2009; Bjørneraas et al. 2012) ainsi que la
disponibilité du couvert de protection (Herfindal et al. 2009; Jiang et al. 2010; Bjørneraas et
al. 2012). La présence de routes modifie également le comportement de l’animal. Elles sont
parfois évitées (Laurian et al. 2012, Paton et al. 2017; Rowland et al. 2018) en raison du
risque qu’elles apportent, parfois sélectionnées (Fortin et al. 2009; Laurian et al. 2012)
lorsque leur utilisation procure des bénéfices sur le plan énergétique. La topographie
(Laurian et al. 2012; Paton et al. 2017, Rowland et al. 2018) et la proximité des cours d’eau
(Traill 2004; Mandlate et al. 2019) influencent également la sélection d’habitat par un
animal. D’autres variables, de types intrinsèques ou temporels, peuvent moduler la sélection
des variables environnementales. Ainsi, un individu ne réagira pas nécessairement de la
même manière à son environnement durant les différentes périodes de sa vie. Explorer
l’impact de ces différentes variables sur la sélection de l’habitat permet d’avoir une meilleure
compréhension de la façon dont les animaux utilisent leur environnement.
Sélection d’habitat : saison
Parmi les variables influençant la sélection de l’habitat, on retrouve les saisons. En
effet, plusieurs études confirment des variations de sélection selon la période de l’année
(Forchhammer 1995; Fortin et al. 2009; Bjørneraas et al. 2011; Laurian et al. 2012; Paton et
al. 2017), ce qui démontre l’importance de considérer l’effet de cette variable. Par exemple,
Forchhammer (1995) mentionne que les prairies sont plus fortement utilisées par les bœufs
musqués (Ovibos moschatus) en saison de croissance végétale, alors que ces derniers
fréquentent davantage les landes sèches buissonneuses basses et les toundras lors des autres
périodes de l’année. De même, les orignaux (Alces alces) utilisent fortement les jeunes
peuplements forestiers du printemps à l’automne puisqu’ils fournissent des ressources
3alimentaires en abondance. Après cette période, ils utilisent davantage de vieilles forêts
puisque celles-ci présentent alors une végétation de meilleure qualité que les autres
environnements (Bjørneraas et al. 2011). Elles offrent aussi un couvert forestier qui limite
l’accumulation de neige, ce qui facilite les déplacements des orignaux (Bjørneraas et al.
2011). Finalement, Fortin et al. (2009) ont trouvé que les bisons (Bison bison) évitent les
cours d’eau en été, alors qu’ils les sélectionnent en hiver.
Sélection d’habitat : sexe
Le sexe de l’individu est également une variable qui peut influencer l’utilisation de
l’environnement par un animal (Dussault et al. 2005) en raison des besoins différentiels des
deux sexes (ex. Herfindal et al. 2009). Des études ont en effet observé que les mâles et les
femelles de plusieurs espèces de grands herbivores ne sélectionnent pas leur habitat de la
même façon : orignal : Bjørneraas et al. 2011, 2012; Laurian et al. 2012; mouflon
méditerranéen (Ovis gmelini musimon x Ovis sp.) : Marchand et al. 2015; cerf rouge (Cervus
elaphus) : Paton et al. 2017. Par exemple, Bjørneraas et al. (2011) ont démontré que les
orignaux mâles sélectionnent fortement les types de milieux présentant une végétation de
grande qualité alors que les femelles sélectionnaient plutôt des types de milieux présentant
un couvert plus fermé afin d’assurer la protection des jeunes. Des conclusions similaires ont
été établies pour les mouflons méditerranéens (Marchand et al. 2015). À l’opposé, Demment
et Van Soest (1985) proposent que les animaux de plus grande taille aient la possibilité de
se nourrir d’une végétation de qualité moindre en raison de leur plus grande capacité
digestive. Par conséquent, chez les animaux sexuellement dimorphiques, les mâles, plus
gros, pourraient utiliser des types d’habitats et de ressources de moins bonne qualité,
comparativement aux femelles (ex. cerf rouge d’Espagne [Cervus elaphus, sous-espèce
hispanicus] : Lazo et al. 1994).
Le type de végétation et la qualité de celle-ci ne sont pas les deux seules variables
qui peuvent être sélectionnées différemment entre les sexes. Laurian et al. (2012) ont trouvé
que les femelles orignaux sélectionnaient les endroits avec une pente plus faible, et ce, toute
l’année, alors que les mâles les sélectionnaient seulement à l’automne. L’évitement des
pentes abruptes est un comportement permettant de réduire ses dépenses énergétiques (Wall
4et al. 2006). Une autre étude a démontré que les femelles de cerfs rouges utilisaient
davantage des secteurs pentus pendant les jours d’automne, potentiellement en réaction à la
présence de chasseurs alors que les mâles n’affichaient pas cette stratégie d’évitement (Paton
et al. 2017). Pour ces deux études, les femelles ont démontré des comportements
d’évitement des routes moindre que les mâles (Laurian et al. 2012; Paton et al. 2017).
Puisque cet évitement chez les mâles était encore plus prononcé à l’automne dans l’étude de
Paton et al. (2017), les auteurs suggèrent qu’il serait en réaction à la pression de chasse plus
élevée pour ceux-ci. À l’automne. Force est de constater que lorsqu’on ne tient pas compte
du sexe et de la saison dans les modèles de sélection d’habitat, il est possible que l’on tire
des conclusions erronées sur la sélection d’habitat d’une espèce ou d’une population. Une
approche de gestion tenant compte de ces aspects est bénéfique pour assurer une
conservation adéquate des animaux et de leur habitat.
Sélection d’habitat : densité
La densité de la population peut avoir un effet tout aussi important sur la sélection
d’habitat que la saison et le sexe des individus. La théorie sur la sélection d’habitat
dépendante de la densité stipule que les animaux devraient utiliser davantage des milieux de
moindre qualité lorsque la densité de la population augmente (Rosenzweig 1991). Ce patron
de sélection découlerait d’une compétition intraspécifique accrue pour les ressources et serait
étroitement associé au fait que les animaux devraient se répartir sur le territoire en fonction
de la profitabilité de l’habitat (Fretwell et Lucas 1969: théorie de la distribution idéale libre;
Rosenzweig 1981, 1991; Morris 2003a). Conséquemment, on devrait observer une
diminution de la sélection d’habitat avec une utilisation plus homogène des différents types
d’habitats quand la densité de la population est élevée (Rosenzweig 1981, 1991; Morris
2003a). Les résultats de plusieurs études convergent en ce sens (ex. Fortin et al. 2008; van
Beest et al. 2014ab, 2016; Rozen-Rechels et al. 2015; McLoughlin et al. 2016; Tomassini et
al. 2019) et confirment la nécessité de tenir compte de l’effet de la densité des individus sur
la sélection d’habitat. Par exemple, van Beest et al. (2014a) ont trouvé que les wapitis
(Cervus canadensis manitobensis) et les orignaux sélectionnent davantage les forêts mixtes,
habitat primaire de grande qualité, en hiver quand leur densité est faible, alors que cette
sélection diminue à plus forte densité. Les habitats de moindre qualité en raison de la menace
5reliée à la chasse et de la plus grande possibilité de détection par les prédateurs sont
également davantage sélectionnés par les deux espèces lorsque la densité de conspécifiques
augmente. Plus précisément, l’augmentation de la densité d’individus a entraîné une plus
grande sélection des champs cultivés par les wapitis et des zones d’infrastructures humaines
par les orignaux. De même, pour les chevaux sauvages (Equus ferus caballus), des densités
élevées près des cours d’eau les obligent à utiliser des habitats de moindre qualité alors que
ce patron de sélection n’est pas observé lorsqu’ils s’éloignent des cours d’eau où la densité
de conspécifiques est plus faible (Rozen-Rechels et al. 2015, see also van Beest et al. 2014b).
Finalement, Fortin et al. (2008) concluent qu’une tactique spécialiste d’utilisation de
l’habitat et des ressources se solde par un meilleur succès reproducteur pour le cerf rouge
sauf lorsque leur densité est très élevée. Dans une telle situation, une tactique généraliste
semble plus optimale pour garantir un meilleur succès reproducteur.
Ne pas considérer l’effet de la densité sur la sélection d’habitat peut entraîner une
interprétation erronée des préférences de l’animal en plus de biaiser l’évaluation de la qualité
des habitats puisqu’une utilisation élevée est souvent associée à une qualité élevée (van Beest
et al. 2014b). Malgré tous ces travaux, encore peu d’études tiennent compte de l’effet de la
densité sur la sélection de l’habitat par les espèces fauniques puisque l’échelle temporelle
considérée est souvent trop courte pour permettre de grandes fluctuations de densité
(McLoughlin et al. 2010). Qui plus est, encore moins d’études s’intéressent aux différences
de comportements entre les mâles et les femelles face à une augmentation de la densité de
conspécifiques. Comme les besoins des mâles diffèrent parfois de ceux des femelles, il est
fort probable que la sélection des différentes composantes de leur environnement ne fluctue
pas de la même façon avec l’augmentation de la population. Les quelques études qui ont
évalué cette potentielle différence dans la sélection dépendante de la densité entre les deux
sexes ont obtenu des variations significatives (e.g., Clutton-Brock et al. 1987, Stewart et al.
2015 [cerf rouge, Cervus elaphus]; Kie et Bowyer 1999 [cerf de Virginie, Odocoileus
virginianus]). Ne pas tenir compte de l’effet de telles variables sur la sélection d’habitat, ni
de l’interaction entre ces variables, peut entraîner une mauvaise compréhension de
l’utilisation de l’habitat d’une espèce. Ces conclusions erronées peuvent quant à elles mener
à des décisions de gestion inadéquates ou inefficaces en appliquant une mesure de gestion
6sur l’ensemble de la population alors que ce ne sont que quelques individus spécifiques qui
sont problématiques par exemple. Afin de mieux comprendre l’effet combiné du sexe et de
la densité de population sur la sélection d’habitat, mon étude utilisera le cas du plus grand
herbivore terrestre : l’éléphant africain (Loxodonta africana, Blumenbach).
Conservation des éléphants
En raison de la grande superficie de son domaine vital, de ses exigences alimentaires
élevées et de son comportement d’alimentation plutôt destructeur, l’éléphant africain est l'un
des herbivores dont la gestion et la conservation sont particulièrement difficiles, et ce
phénomène est exacerbé dans les réserves de petites superficies. L’éléphant est en effet un
animal migrateur qui utilise plusieurs types de milieux afin de suivre les fluctuations
d’abondance et de qualité de la végétation pour satisfaire ses besoins nutritifs (Loarie et al.
2009). Animal emblématique pour le développement de l’industrie écotouristique (Garai et
al. 2004), il a été réintroduit dans plusieurs petites aires protégées au cours des dernières
décennies. Longtemps chassé par les humains en raison de conflits entre les deux espèces, il
est maintenant classé comme espèce vulnérable (Blanc, 2008). Toutefois, il est fréquent
d’observer d’importantes augmentations de populations dans les petites réserves clôturées
de l’Afrique du Sud puisqu’il bénéficie d'une protection accrue et profite de la création de
points d’eau artificiels (Slotow et al. 2005; van Aarde et Jackson 2007). Cependant, comme
tout autre herbivore de grande taille, sa conservation dans de telles réserves peut avoir des
conséquences néfastes sur les communautés végétales en réduisant significativement
l’abondance des espèces qu'il consomme (ex. Wiseman et al. 2004, Guldemond et van Aarde
2008, Young et al. 2009, Shrader et al. 2012).
Pour ajouter à la complexité de la conservation de cette espèce emblématique, la
viabilité des efforts de réintroduction d'éléphants dans des aires protégées de faible superficie
a largement été basée sur des prévisions de croissance de populations observées en milieux
naturels (Slotow et al. 2005). Or, il s’avère que ces dernières ont un taux de croissance plus
faible que les populations retrouvées dans de petites aires protégées. Par exemple, Slotow et
al. (2005) ont calculé un taux annuel de croissance de 15% pour des petites populations
confinées d’éléphants, taux de loin supérieur aux taux maximums viables prédits par Hanks
et McIntosh (4%; 1973) et Calef (7%; 1988). Une autre étude, réalisée en 2006 dans huit
7réserves de petites superficies en Afrique du Sud, a aussi montré que la croissance annuelle
moyenne des populations d’éléphants réintroduites est plus importante que prévue (Mackey
et al. 2006). La croissance est telle que certaines populations de petites réserves clôturées
pourraient devenir en situation de surabondance dans un horizon d’une ou deux décennies
en l'absence de mesures de réduction des populations et ainsi entraîner une dégradation de
l’habitat sur le long terme et une perte de biodiversité (Mackey et al. 2006). Pour évaluer la
viabilité de maintenir une population d’éléphants dans des aires protégées clôturées de faible
superficie, il importe donc de comprendre ce qui influence l’utilisation et la sélection de
l’habitat par celui-ci.
Sélection d’habitat par l’éléphant
La sélection de l’habitat par l’éléphant est influencée par de nombreux facteurs: la
disponibilité des ressources (Venter et al. 2015), la productivité de la végétation (Bastille-
Rousseau et al. 2019), la répartition des sources d’eau (Smit et al. 2007, Di Minin et al.
2013), la présence humaine (Wall et al. 2013), la topographie (Ashiagbor and Danquah
2017), le sexe (Leggett 2006) et la densité de la population (Robson et van Aarde 2018). En
raison de ses demandes énergétiques élevées, l’éléphant réagit davantage aux variations
d’abondance de végétation que d’autres espèces (Harris et al. 2008; de Beer et van Aarde,
2008). Des études se sont également intéressées à la variation intra-annuelle (entre les
saisons) de la sélection des types de formations végétales par l’éléphant. Selon Loarie et al.
(2009), les éléphants sélectionnent des secteurs à qualité/abondance végétale variable (boisés
ouverts, prairies) en saison humide alors qu’en saison sèche, ils sélectionnent des secteurs
donc la qualité et l’abondance végétales varient moins annuellement et qui présentent une
couverture végétale supérieure à celle des secteurs avoisinants (ex. boisés fermés). Cette
sélection différentielle selon la saison leur permettrait de sélectionner les secteurs les plus
productifs (i.e., plus « verts ») de leur environnement à un moment donné (Loarie et al. 2009;
Orrick 2018). La sélection de la qualité et l’abondance de la végétation semble légèrement
plus forte en saison humide quand l’eau est abondante et peu limitante (Loarie et al. 2009;
Wall et al. 2013).
La sélection des types de végétation par les éléphants peut varier selon le sexe. Par
exemple, Smit et al. (2007) ont trouvé une sélection plus marquée du couvert végétal par les
8troupeaux mixtes que par les groupes de mâles dans une étude comprenant sept populations
d’éléphants. Inversement, cette sélection était plus marquée pour les mâles dans une autre
étude réalisée au Parc national Kruger (PNK; de Knegt et al. 2011). On a également observé
qu’une augmentation de la densité d’éléphants au PNK a engendré un changement dans la
sélection de l’habitat par les femelles en saison sèche, celles-ci utilisant de moins en moins
les secteurs de meilleure qualité que constituent les boisés denses (Robson et van Aarde
2018).
L’impact des humains sur l’utilisation de l’habitat par l’éléphant a été évalué à
maintes reprises en considérant différentes variables anthropiques. En général, l’éléphant a
tendance à éviter les routes et à ne pas s’approcher des humains (Pinter-Wollman 2009;
Roever et al. 2012). Toutefois, comparativement aux femelles, les mâles s’approchent plus
fréquemment et plus près des villages (Cook et al. 2015). Ce patron est particulièrement
observé en saison sèche. De plus, les clôtures ont souvent un effet répulsif pour les éléphants,
ces derniers sélectionnant davantage la portion centrale de l’aire protégée. Les clôtures
engendrent donc un important effet de bordure (Vanak et al. 2010). Ce patron est toutefois
observé même lorsque l’aire protégée est non-clôturée. Kiffner et al. (2013) ont en effet
remarqué une plus grande utilisation des habitats éloignés de la limite de l’aire protégée, un
patron d’utilisation probablement associé à la chasse illégale.
La topographie influence également la sélection de l’habitat des éléphants en raison
des dépenses énergétiques plus importantes qu’elle peut engendrer. Ainsi, les secteurs pentus
peuvent être moins fréquentés par les éléphants (Wall et al. 2006; Roever et al. 2012). Au
Parc national Mole, l’altitude est la variable influençant le plus la répartition des éléphants
(Ashiagbor et Danquah 2017). Toutefois, la topographie peut également engendrer des
différences au niveau de la qualité de la végétation. Par exemple, l’utilisation de secteurs de
faible élévation en saison sèche (Shannon et al. 2006a; Marshal et al. 2011) peut être
expliquée par l’abondance de nutriments et la capacité de rétention d’eau des sols des
secteurs situés en bas de pente (Shannon et al. 2006a).
9Vous pouvez aussi lire

















































