L'acidose chez la chèvre laitière et l'usage du carbonate de potassium (K2CO3) : Impact sur les composantes du lait et la production - Corpus UL
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
L'acidose chez la chèvre laitière et l'usage du carbonate de potassium (K2CO3) : Impact sur les composantes du lait et la production Mémoire Stéphanie Dion Maîtrise en sciences animales - avec mémoire Maître ès sciences (M. Sc.) Québec, Canada © Stéphanie Dion, 2021
L’acidose chez la chèvre laitière et l’usage du carbonate de potassium (K2CO3) : impact sur les composantes du lait et la production Mémoire Stéphanie Dion Sous la direction de : Yvan Chouinard, directeur de recherche Daniel Rico, codirecteur de recherche
Résumé Le carbonate de potassium (K2CO3) est un additif alimentaire disponible sur le marché souvent recommandé dans l’alimentation des bovins laitiers pour limiter l’acidité du rumen à la suite d’une ingestion importante de glucides rapidement fermentescibles et prévenir la baisse du taux de matière grasse du lait qui en résulte. Chez la chèvre laitière, les effets du K2CO3 sont méconnus. L’objectif de ce projet de recherche vise à évaluer l’utilisation du K2CO3 comme traitements préventif et/ou curatif, pouvant empêcher la chute des matières grasses du lait chez des chèvres laitières en début de lactation recevant une ration riche en concentrés. Chez des chèvres de race Alpine, l’apport d’une ration riche en concentrés en début de lactation (rapport fourrage:concentrés de 45:55) a eu à long terme, des répercussions sur les performances et a provoqué une inversion des composantes du lait, c.-à-d. que le taux de matière grasse était inférieur au taux de protéines au terme des 56 jours de traitement. Dans les conditions de l'expérience actuelle, l’ajout du K2CO3 comme traitement préventif et/ou curatif n’a pas permis d’éviter le risque d’une chute des matières grasses laitières chez des chèvres en début de lactation recevant une ration riche en concentrés. ii
Abstract The objective of this study was to verify the use of potassium carbonate (K2CO3) as a preventive and/or a recovery treatment to prevent a low milk fat production when early-lactating dairy goats are receiving a diet rich in concentrates. A commercially available supplement of K2CO3 is often recommended for dairy cattle in early lactation to limit the rumen acidity following the intake of diets high in rapidly fermentable carbohydrates. In dairy goats, the effects of K2CO3 are unknown. In this experiment conducted in Alpine dairy goats, high intake in concentrates (forage:concentrate ratio 45:55) had long-term effects on lactation performances, and caused a reversal in the proportion of major components of the milk, i.e., the fat content was lower than the protein content after 56 days of treatment. Under the conditions of this current experiment, the addition of K2CO3 as a preventive and/or a recovery treatment did not affect milk fat production when early lactating dairy goats were receiving a high-concentrate diet. iii
Table des matières Résumé .................................................................................................................................................................ii Abstract ................................................................................................................................................................ iii Table des matières ...............................................................................................................................................iv Liste des figures ...................................................................................................................................................vi Liste des tableaux ............................................................................................................................................... vii Liste des abréviations en français ...................................................................................................................... viii Liste des abréviations en anglais .........................................................................................................................ix Remerciements ................................................................................................................................................... xii Avant-propos .......................................................................................................................................................xv Introduction ........................................................................................................................................................... 1 Chapitre 1 Revue des travaux antérieurs ............................................................................................................. 3 1.1 Caractéristiques générales des matières grasses laitières chez la chèvre ........................................... 3 1.1.1 Anatomie et physiologie de la mamelle caprine ............................................................................. 3 1.1.2 Composition et structure des matières grasses ............................................................................. 5 1.1.3 Profil en acide gras du lait .............................................................................................................. 6 1.2 Métabolisme des matières grasses alimentaires du rumen au prélèvement par la glande mammaire . 7 1.2.1 Lipolyse des matières grasses alimentaires................................................................................... 8 1.2.2 Biohydrogénation des acides gras insaturés.................................................................................. 9 1.2.3 Biosynthèse des acides gras microbiens ..................................................................................... 11 1.2.4 Digestion et absorption des matières grasses alimentaires et microbiennes .............................. 12 1.3 Lipogenèse de la glande mammaire .................................................................................................... 13 1.3.1 Synthèse des acides gras de novo par les cellules de la glande mammaire ............................... 14 1.3.2 Prélèvement des acides gras préformés par la glande mammaire .............................................. 14 1.4 Facteurs influençant la teneur en matière grasse laitière chez la chèvre ............................................ 15 1.4.1 Facteurs intrinsèques ................................................................................................................... 16 1.4.1.1 Sélection génétique ............................................................................................................. 16 1.4.1.2 Stade de lactation et parité .................................................................................................. 17 1.4.1.3 Santé du pis ......................................................................................................................... 18 1.4.2 Facteurs extrinsèques .................................................................................................................. 19 1.4.2.1 Photopériode ....................................................................................................................... 19 1.4.2.2 Saison .................................................................................................................................. 20 iv
1.4.2.2.1 Stress de chaleur .......................................................................................................... 20 1.4.2.3 Rapport fourrage:concentrés / Pourcentage de concentrés ................................................ 21 1.4.2.3.1 Acidose ruminale .......................................................................................................... 22 1.4.2.3.2 Théorie de la chute de la matière grasse du lait ........................................................... 24 1.5 Additifs alimentaires pour moduler le pH ruminal ................................................................................ 27 1.5.1 L’apport en substance tampon ..................................................................................................... 27 1.5.1.1 Apport de K2CO3 sur l’environnement ruminal et la synthèse de la matière grasse du lait . 27 1.6 Objectif de ce mémoire ........................................................................................................................ 32 1.7 Références ........................................................................................................................................... 33 Potassium carbonate as a supplement to improve milk fat concentration and yield in early- lactating dairy goats fed a high-starch, low-fiber diet ......................................................................................... 42 2.1 Résumé ................................................................................................................................................ 43 2.2 Interpretative summary ........................................................................................................................ 44 2.3 Abstract ................................................................................................................................................ 45 2.4 Introduction .......................................................................................................................................... 46 2.5 Materials and methods ......................................................................................................................... 46 2.5.1 Goats, Experimental Design, Diet and Treatments ...................................................................... 46 2.5.2 Experimental Measurements, Samplings and Analyses .............................................................. 47 2.5.2.1 Body Weight and Feed Intake. ............................................................................................ 47 2.5.2.2 Milk. ..................................................................................................................................... 48 2.5.2.3 Ruminal Fluid. ...................................................................................................................... 48 2.5.2.4 Blood Samples. ................................................................................................................... 49 2.5.3 Statistical Analysis ........................................................................................................................ 49 2.6 Results and Discussion ........................................................................................................................ 50 2.6.1 Treatment Assessment ................................................................................................................ 50 2.6.2 DMI, BW and Milk Yield ................................................................................................................ 51 2.6.3 Ruminal Fermentation .................................................................................................................. 52 2.6.4 Blood Parameters ......................................................................................................................... 53 2.6.5 Major Constituents, FA Profile and Minerals of Milk..................................................................... 53 2.7 Conclusions ......................................................................................................................................... 54 2.8 Acknowledgements .............................................................................................................................. 54 2.9 References ........................................................................................................................................... 56 Conclusion générale ........................................................................................................................................... 74 Bibliographie ....................................................................................................................................................... 77 v
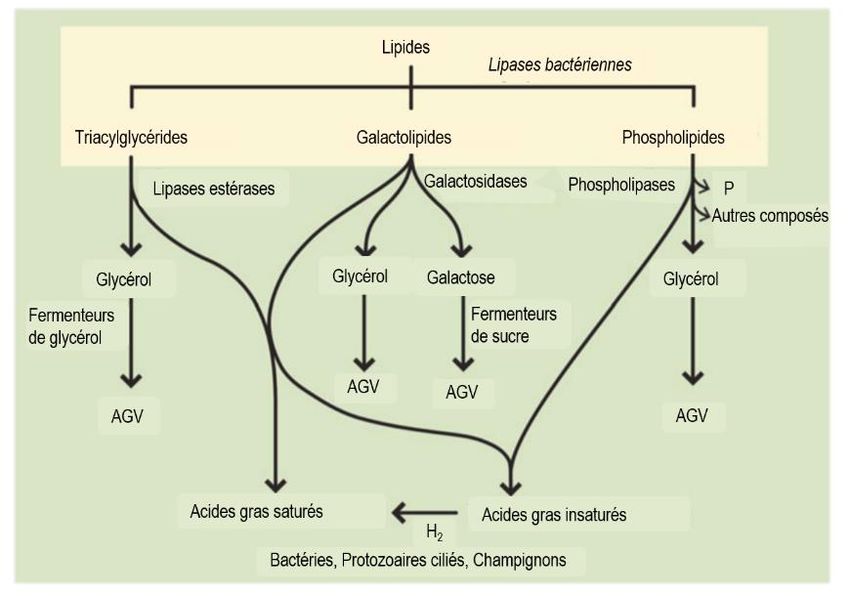
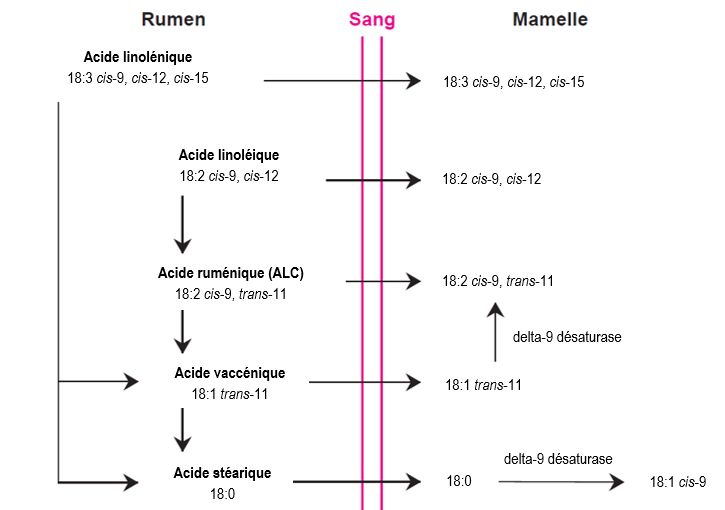
Liste des figures Figure 1.1 Structure de la mamelle d’une chèvre laitière montrant les parties sécrétrices et citernales des glandes (a) ainsi que l’organisation des alvéoles (b). Tirée de l’Institut de l’élevage (2006). .............................. 3 Figure 1.2 Lait de chèvre centrifugé. (1) trois globules de gras contiennent du matériel cytoplasmique d'un côté (x 4800). (2) À plus grand grossissement (x 28 000) du matériel cytoplasmique des globules de gras, on peut voir une mitochondrie, des ribosomes et des citernes du réticulum endoplasmique rugueux. Tirée de Wooding et al. (1970). .......................................................................................................................................... 4 Figure 1.3 Schématisation de l’hydrolyse des matières grasses alimentaires dans le rumen. AGV = Acides gras volatils. Adaptée de Nagaraja (2016). .......................................................................................................... 9 Figure 1.4 Les voies de biohydrogénation dans des conditions ruminales normales (trans-11; flèches vertes) et alternatives (trans-10; flèches rouges; lors de perturbations ruminales) des acides -linolénique et linoléique. Adaptée de Ferlay et al., 2017 et Fougère, 2018. ............................................................................ 10 Figure 1.5 Synthèse des acides gras par les microorganismes du rumen. L’astérisque (*) représente l’absorption des acides gras alimentaires par les bactéries du rumen et les astérisques (**) représentent l’α- oxydation des acides gras à chaîne paire. Adaptée de Vlaeminck et al. (2006)................................................ 12 Figure 1.6 Principales voies de production des acides gras trans et des acides linoléiques conjugués du lait. Adaptée de Chilliard et al. (2001). ...................................................................................................................... 15 Figure 1.7 Évolution de la teneur en matière grasse et de la production de lait en fonction du stade de lactation chez la chèvre. Adaptée de Ciappesoni et al. (2004). ......................................................................... 18 Figure 1.8 Effet de la température ambiante sur la consommation alimentaire des chèvres de race Alpine nourries avec un régime fourrager à 100 % ou un régime avec un rapport fourrage:concentrés de 50:50. Adaptée de Lu et Richard (1986), citée par Lu (1989). ...................................................................................... 21 Figure 1.9 Conséquences métaboliques d’un apport important de glucides rapidement fermentescibles dans la ration sur le pH ruminal et les populations microbiennes du rumen. Adaptée de Schwartzkopf-Genswein et al. (2003). ........................................................................................................................................................... 23 Figure 2.1 Milk yield (A), milk fat concentration (B) and yield (C), milk fat-to-protein ratio (D), ruminal pH (E), milk fat trans-11 18:1 (F) and trans-10 18:1 (G) concentrations, trans-11-to-trans-10 18:1 ratio (H), de novo milk fatty acid concentration (I) and daily secretion (J), as well as preformed milk fatty acid concentration (K) and daily secretion (L) in goats fed a diet with a forage-to-concentrate ratio of 55:45 (Baseline period; 27 ± 4 DIM) or 45:55 (Period 1; 55 ± 4 DIM) on a DM basis. Dots = Least squares means (n = 20). P-values are presented for the confounded effects of time and diet. ...................................................................................... 73 vi
Liste des tableaux Tableau 1.1 Teneurs moyennes et leurs coefficients de variation (CV) des classes de lipides retrouvées dans les matières grasses laitières des vaches et des chèvres. .................................................................................. 6 Tableau 1.2 Le profil en acides gras du lait de vache, de brebis et de chèvre.................................................... 7 Tableau 1.3 Variation de la composition du lait de différentes races caprines selon le pays............................ 16 Tableau 1.4 Production et composition du lait de chèvres présentant des comptes de cellules somatiques bas, moyens et élevés à 200 jours de lactation. ........................................................................................................ 19 Table 2.1 Composition of the experimental TMR............................................................................................... 60 Table 2.2 Nutrient composition of experimental TMR ........................................................................................ 61 Table 2.3 Intake, body weight, milk yield, feed efficiency, and refusal composition of early-lactation dairy goats fed a high-concentrate diet without K2CO3 for two periods (control treatment), with K2CO3 for two periods (preventive treatment), and with K2CO3 only in the second period (recovery treatment). .................................. 63 Table 2.4 Rumen pH and concentrations of NH3-N and VFA in early-lactation dairy goats fed a high- concentrate diet without K2CO3 for two periods (control treatment), with K2CO3 for two periods (preventive treatment), and with K2CO3 only in the second period (recovery treatment). ..................................................... 65 Table 2.5 Blood parameters of early-lactation dairy goats fed a high-concentrate diet without K2CO3 for two periods (control treatment), with K2CO3 for two periods (preventive treatment), and with K2CO3 only in the second period (recovery treatment). .................................................................................................................. 66 Table 2.6 Concentration and yield of major milk constituents, and mineral profile of milk in early-lactation dairy goats fed a high-concentrate diet without K2CO3 for two periods (control treatment), with K2CO3 for two periods (preventive treatment), and with K2CO3 only in the second period (recovery treatment). .................................. 67 Table 2.7 Milk fat composition of early-lactation dairy goats fed a high-concentrate diet without K2CO3 for two periods (control treatment), with K2CO3 for two periods (preventive treatment), and with K2CO3 only in the second period (a recovery treatment). ............................................................................................................... 69 vii
Liste des abréviations en français AGV Acide gras volatil ALC Acide linoléique conjugué CCS Comptage de cellules somatiques CoA Coenzyme A DACA Différence alimentaire cations-anions MAPAQ Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec MS Matière sèche mÉq Milliéquivalent VLDL Lipoprotéines de très faible densité viii
Liste des abréviations en anglais ADF Acide detergent fiber BW Body weight CP Crude protein CON Control treatment DCAD Dietary cation-anion difference DHIA Dairy herd improvement association DIM Days in milk DM Dry matter DMI Dry matter intake FA Fatty acid FAME Fatty acid methyl esters FCM Fat-corrected milk N Number of samples NDF Neutral detergent fiber OM Organic matter PREV Preventive treatment PUFA Polyunsaturated fatty acids R2 Coefficient of determination REC Recovery treatment SCC Somatic cell count SCS Somatic cell score SEM Standard error of the mean TMR Total mixed ration VFA Volatile fatty acid ix
A ma fille Élisabeth et mon loup Alex, x
“Si la chèvre avait la queue plus longue, elle pourrait balayer les étoiles.” Proverbe tchèque xi
Remerciements Le présent mémoire fait l’objet d’un travail d’équipe en son entier. Sans l’aide des collaborateurs, je n’aurais pu mener à bien ce projet de recherche. Tout d’abord, j’aimerais remercier mon directeur de maîtrise, Yvan. Merci pour ton support, ta rigueur et ta révision tout au long de ce projet. Tu m’as transmis sans équivoque ta passion pour la recherche et pour la finesse du détail. J’ai toujours adoré discuter de nouvelles idées de projets avec toi. Les compétences que j’ai acquises à tes côtés m’ont assurément fait évoluer en tant que jeune agronome et je me considère très choyé d’avoir pu travailler au sein de ton excellente équipe. Merci à Rachel et Marie-Ève d’avoir cru en mon habilité à réaliser ce beau projet. Vous avez été comme des mères dans cette belle aventure. Vous avez toujours été disponibles, empathiques et rassurantes lorsque j’avais des questions ou des entêtements. Merci de m’avoir encouragée dans toutes les opportunités qui se sont présentées au fil de mon parcours, clin d’œil au Zoo. À chacune votre façon, vous avez été déterminantes dans mon cheminement personnel et professionnel, je vous en serai toujours reconnaissante. Merci également pour tous vos conseils de « Mamans », ils ont été énormément précieux. Je vous aime! Merci, Micheline et Yolaine pour votre support technique et votre surveillance au laboratoire. Merci à tous les étudiant.es et stagiaires du Département en sciences animales ainsi que Geneviève Bégin d’Agriculture et Agroalimentaire Canada, qui ont contribué à la réalisation de mes analyses au laboratoire. Merci également à Annie et Annick pour avoir réalisé les prises de sang des chèvres lors de la phase expérimentale. Merci au Centre de recherche en sciences animales de Deschambault (CRSAD) de m’avoir accueilli comme l’une des vôtres au sein de l’équipe et de m’avoir permis de réaliser ce projet de recherche grâce à la ferme expérimentale caprine. Un merci tout spécial à Janie de m’avoir pris par la main à mon retour de France et de m’avoir transmis son expertise et sa rigueur afin de réaliser une phase expérimentale impeccable. Je n’oublierai jamais nos doux matins en voiture entourés d’un bon café McDo réconfortant pour débuter une folle journée de collecte. Merci à Daniel, mon codirecteur de maîtrise, pour son expertise et son soutien scientifiques. Je suis toujours émerveillée devant ton désir de comprendre les principes fondamentaux des productions animales. Je remercie également le personnel agricole du CRSAD, Michael, Luc et Paul. Merci de m’avoir aidée avec la phase expérimentale de mon projet. Vous avez enrichi mon expérience à la ferme avec chacune de vos personnalités bien distinctes. J’aimerais également remercier les partenaires financiers, le Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec et Agriculture et Agroalimentaire Canada, qui ont permis de concrétiser ce projet par le biais du Programme Innov’Action Agroalimentaire dans le cadre de l’entente Cultivons L’Avenir 2. xii
Merci à Chantal Lemieux et feu Alain Fournier du Ministère de l'Agriculture, des Pêcheries et de l'Alimentation du Québec pour leur aide dans la conceptualisation et la réalisation de ce projet. Merci également au local 4206 du Département des sciences animales et à tous les étudiants aux études graduées qui ont été sur mon passage, vous avez été ma vie sociale pendant toutes ces belles années. J’aimerais remercier tout particulièrement des personnes avec qui j’ai définitivement développé des affinités avec les années et avec qui, un jour qui c’est, je travaillerai. Merci, Maxime, ta personnalité singulière m’a convaincu de devenir membre de l’ACCESA ainsi que du Comité santé et recherche du CEPOQ. Ces engagements ont enrichi mon expérience aux études graduées et ils ont définitivement été contributoires à mon parcours professionnel en tant qu’agronome, je t’en suis reconnaissante. Marguerite, une athlète hors pair, brillante et à la fois sensible et familiale. Tu as été une voisine de bureau exemplaire, je ne me suis jamais sentie jugée et tu as toujours été volontaire à rire de mes bêtises ou juste à m’écouter. Merci pour les échanges à la fois scientifiques ou justes sur la vie en général. Ta présence à ma gauche a définitivement embelli mes journées de travail. Anne-Marie, « ma partner in crime », tu es une personne inspirante, inarrêtable, sportive, intelligente et une maman dévouée. Notre différence d’âge n’a jamais paru et c’est la beauté de notre amitié. Toujours remplie d’histoires et de péripéties, tu ne te laisses jamais abattre. Pour moi, tu es un modèle de force et une source d’inspiration. Merci d’avoir été une confidente tout au long de mon parcours universitaire. Véronique, la personne la plus empathique, travaillante, dévouée et déterminée que je connaisse. Nos débuts ont commencé dans un des plus grands malaises, mais il a certainement valu la peine. Tu es un modèle d’excellence pour moi et tu as certainement eu une influence déterminante sur mon parcours aux études graduées. J’ai adoré et j’adore partager avec toi des cafés, des dîners, des marches, des courses, des cours de TRX et de natation, une journée de raquette (je continue à croire que c’est ma faute si nous sommes restées prises dans la neige lorsque tu conduisais), des congrès, l’enseignement, des moments difficiles, des moments de gloire, des discussions loufoques, des « Bonnes journées » presque tous les jours malgré la distance, des rêves de voyages improbables et des rénovations. Je devais faire une légère énumération pour mettre en perspective nos dernières années, mais également notre amitié. Quétaine, mais sincère, je me considère très chanceuse d’avoir une amie d’exception comme toi. Je nous souhaite encore plein d’années de fous rires et une retraite bien méritée. Merci d’être là. Merci à ma grande famille (Papa, Alex, Chantall, Laurence, Christian, Sarah-Jeanne, Isarose, Carole, François, Lucas, Milène et Michel) de m’avoir encouragée tout au long de mes études. C’est définitivement la fin. Enfin, j’aimerais remercier mes premiers amours. Alex, tu m’as encouragée et tu as été à mes côtés du début jusqu’à la fin de mon parcours universitaire sans jamais douter de mes capacités. Merci de me pousser xiii
à être la meilleure version de moi-même, en tant qu’étudiante, conjointe et nouvellement maman. Merci de toujours croire en moi et de me rassurer dans les moments où je me remets le plus question...Dieu c’est que ça arrive. Je t’aime mon Loup. Je suis fébrile de connaître la suite de notre histoire à l’amour est dans le pré. Élisabeth, ma fille, tu es ma plus belle surprise et c’est du haut de tes 15 mois que tu m’encourages à terminer cette belle et grande expérience. J’adore te regarder t’émerveiller devant les petites choses de la vie. Je me dépêche d’écrire ses mots pour aller jouer et découvrir le monde avec toi. Je t’aime! Maman. xiv
Avant-propos Le deuxième chapitre du présent mémoire contient un manuscrit d’article scientifique rédigé entièrement en anglais. Je suis l’auteure principale de cet article et j’ai réalisé majoritairement tous les travaux de recherches (collecte de données, analyses en laboratoire, statistiques et rédaction) toujours sous la supervision de mon directeur Yvan Chouinard, de mon codirecteur Daniel Rico et de mes superviseures Rachel Gervais, Marie-Ève Brassard et Janie Lévesque. L’article scientifique a été soumis et accepté dans le Journal of Dairy Science sous le titre suivant avec ses auteurs : Potassium carbonate as a supplement to improve milk fat concentration and yield in early-lactating dairy goats fed a high-starch, low-fiber diet S. Dion1, M.E. Brassard1, J. Levesque2, D. E. Rico2, G. F. Tremblay3, R. Gervais1 and P. Y. Chouinard1 1Département des Sciences Animales, Université Laval, Québec, QC, Canada G1V 0A6 2Centre de recherche en sciences animales de Deschambault, Deschambault, QC, Canada G0A 1S0 3Québec Research and Development Centre, Agriculture and Agri-Food Canada, Québec, QC, Canada, G1V 2J3 xv
Introduction Selon le Portrait-diagnostic sectoriel de l’industrie caprine au Québec réalisé en 2017 par le Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ), la production canadienne de lait de chèvre se partage à 98 % entre la province de l’Ontario et celle du Québec. Au cours de la dernière décennie, les parts ontariennes du marché du lait de chèvre ont passé de 72 % en 2010 à 79 % en 2015 contre 28 % à 21 % pour celles de la province de Québec pour ces mêmes années. Cette baisse de 7 % des parts de marché de la province de Québec ne s’est toutefois pas traduite par une diminution de la quantité de lait produit. En moyenne entre 2010 et 2015, la production de lait de chèvre est restée stable se traduisant par un volume total de 11,0 millions de litres par année et par des recettes monétaires s’approchant de 11,4 millions de dollars (MAPAQ, 2017). À l’instar du nombre d’entreprises et du cheptel caprin, le prix du lait de chèvre est aussi resté relativement stable (104,3 $/hl en 2019). Cette stagnation est la conséquence d’une compétitivité féroce avec l’Ontario où les prix sont plus bas en raison d’un surplus de lait sur le marché. La consommation de lait chèvre au Québec est encore très marginale puisqu’elle occupe moins de 1 % des parts de marché de l’ensemble du lait de consommation vendu. Pourtant, un engouement pour les produits transformés à base de lait de chèvre comme le lait de consommation, le yogourt et le beurre ont été observés entre 2010 et 2016 alors qu’une augmentation de 10 % a été rapportée (MAPAQ, 2017). La transformation fromagère demeure cependant le principal marché (80 à 85 %; MAPAQ, 2017). D’ailleurs, en 2016, on estimait cette production à 895 tonnes (MAPAQ, 2018) comptabilisant plus de 200 fromages de chèvre québécois (MAPAQ, 2017). Selon le programme de contrôle laitier caprin de Lactanet en 2016, la production moyenne d’une chèvre était de 817,5 litres par lactation ce qui représente une amélioration de la productivité de 3,8 % par rapport à 2010 (MAPAQ, 2018). Récemment, les producteurs québécois ont eu des conflits avec les transformateurs fromagers pour des raisons reliées principalement à un prix sous compétitif et à la qualité du lait (Jung, 2018). Pour ces raisons, certains transformateurs ont délaissé le marché québécois pour celui de l’Ontario (Samson, 2019). La matière grasse du lait peut être affectée par plusieurs éléments intrinsèques à l’animal (génétique, stade de lactation, santé du pis) et à l’environnement (photopériode, saison, traite), mais l’alimentation est souvent l’élément prédominant (Bauman et Griinari, 2001). En 2015, 63 % des troupeaux caprins sous la supervision de Lactanet avaient un contrôle laitier ou plus pendant l’année où l’on pouvait observer des inversions de taux des composantes, c.-à-d. où le taux de matière grasse était inférieur au taux de protéines (Brunelle, communication personnelle, 2016). La qualité de certains fromages est négativement affectée par une inversion de composants, car la texture des produits obtenus est qualifiée de granuleuse et le goût recherché de « chèvre » est manquant ou moins prononcé (Morand-Fehr et al., 2007). L’inversion de 1
composantes du lait peut être liée à des conditions d’acidose ruminale subclinique (Sandrucci et al., 2019). L’acidose subclinique peut être induite par des régimes alimentaires riches en concentrés en début de lactation visant à combler les besoins élevés des animaux et à engendrer une baisse du rapport matières grasses:protéines du lait (Giger-Riverdin et al., 2014). Cette maladie nutritionnelle est causée par une acidification importante du rumen affectant sa flore et sa faune et influençant ultimement la dégradation des nutriments. Dans de telles conditions, l’hydrogénation des acides gras alimentaires par les bactéries (la biohydrogénation) du rumen peut être altérée et un déplacement vers des voies alternatives de biohydrogénation est privilégié produisant des intermédiaires appartenant à la famille des acides gras trans-10 (18:2 trans-10, cis-12 et 18:1 trans-10). Ces acides gras trans-10 sont connus comme ayant des effets inhibiteurs directs ou indirects sur la synthèse des matières grasses laitière (Shingfield et al., 2010). Des études récentes chez la vache laitière ont montré qu'une supplémentation en carbonate de potassium (K2CO3) pourrait permettre de mieux contrôler le pH du rumen et la teneur en matières grasses du lait (Harrison et al., 2012; Jenkins et al., 2014). À notre connaissance chez la chèvre laitière, les effets potentiels du K2CO3 sont méconnus. Ainsi, l’objectif des travaux présentés dans ce mémoire était de valider l’utilisation du K2CO3 comme traitement préventif ou curatif contre l’acidité du pH ruminal et l’inversion des composantes du lait chez des chèvres recevant une ration riche en concentrés en début de lactation. Ce mémoire se décline en deux chapitres. En premier lieu, le chapitre 1 consiste en une revue des travaux antérieurs portant sur les caractéristiques de la matière grasse du lait caprin, le métabolisme des gras alimentaires et les facteurs pouvant influencer la synthèse de gras du lait chez la chèvre. De surcroît, des moyens pouvant limiter l’acidité du rumen ainsi que les connaissances actuelles sur la supplémentation en K2CO3 dans l’alimentation de la vache laitière seront présentées. En deuxième lieu, le Chapitre 2 présente, sous forme d’article scientifique, l’expérience que nous avons menée et les résultats de recherche y étant associés. En somme, l’objectif poursuivi dans ce mémoire est de démystifier les effets de l’utilisation du K2CO3 comme supplément alimentaire en début de lactation chez des chèvres recevant une ration riche en concentrés. . 2
Chapitre 1 Revue des travaux antérieurs 1.1 Caractéristiques générales des matières grasses laitières chez la chèvre La chèvre se distingue des autres ruminants laitiers, entre autres, par l’anatomie de sa glande mammaire et par la composition de son lait. 1.1.1 Anatomie et physiologie de la mamelle caprine La mamelle caprine est composée de deux glandes indépendantes (Figure 1.1a; Institut de l’élevage, 2006). Chacune contient une partie supérieure responsable de sécréter le lait (le tissu sécrétoire) ainsi qu’une partie inférieure responsable d’emmagasiner le lait dans un réservoir (la citerne). Le tissu sécrétoire est composé de milliards d’alvéoles. L’alvéole est le lieu dans lequel le lait est sécrété. Elle est recouverte d’une couche de cellules épithéliales accolées (lactocytes) qui synthétisent le lait à partir des éléments du sang irriguant les glandes mammaires (Figure 1.1b). L’expulsion du lait des lactocytes se fait par les cellules myoépithéliales (cellules musculaires qui entourent l’alvéole) qui sous l’effet de l’ocytocine (une hormone) se contractent et relâchent le lait dans la lumière des alvéoles. Par la suite, le lait est drainé du milieu alvéolaire vers la citerne de la glande et du trayon par l’intermédiaire de canaux galactotrophes/lactifères. Un même canal galactotrophe qui rassemble un groupe de 10 à 100 alvéoles se nomme un lobule et plusieurs lobules assemblés forment un lobe. Sur le plan histologique, la glande mammaire caprine ressemble beaucoup à celle des bovins et des ovins (Lemelin et al., 2009; Institut de l’élevage, 2006). Figure 1.1 Structure de la mamelle d’une chèvre laitière montrant les parties sécrétrices et citernales des glandes (a) ainsi que l’organisation des alvéoles (b). Tirée de l’Institut de l’élevage (2006). 3
La sécrétion du lait découle de deux fonctions essentielles chez les mammifères : 1) une fonction de filtration du sang des vaisseaux qui entourent les alvéoles (eau, minéraux, protéines sanguineset l’azote non protéique); et 2) une fonction de synthèse des cellules sécrétrices à partir des métabolites sanguins (lactose, matières grasses, protéines coagulables (les caséines) et les protéines solubles (présentent dans le lactosérum) (Pradal, 2012). De façon successive lors de la traite, trois fractions de lait sont sécrétées : le lait citernal, le lait alvéolaire et le lait d’égouttage. Les citernes des glandes mammaires des petits ruminants sont proportionnellement plus grandes que celles des vaches laitières. Elles y représentent entre 40 à 80 % du volume total (Marnet et Mckusick, 2001). Ainsi, le lait récolté à la traite chez la chèvre provient à environ 70 à 80 % de la citerne et 20 à 30 % des alvéoles (Institut de l’élevage, 2006). Cette caractéristique physique permet aux chèvres d’être traites dans un temps moyen de trois minutes, ce qui est très rapide (de Crémoux et al., 2016); et puisque la majorité du lait se situe dans la citerne, il n’est pas nécessaire de stimuler la mamelle avant la traite (Institut de l’élevage, 2006). Toutefois, la stimulation par la trayeuse est essentielle, car elle déclenche une décharge d’ocytocine qui est responsable de l’expulsion des matières graisses laitières qui se trouvent à 75 % dans la fraction alvéolaire (Labussière, 1988). Par ailleurs chez les caprins, la sécrétion du lait est de type apocrine (Dulin et al., 1982). L’existence de ce type de sécrétion a été identifiée chez la chèvre par Wooding et al. (1970) lors d’une étude par microscopie électronique du matériel cellulaire dans des globules de gras (Figure 1.2). La sécrétion apocrine est le détachement par pincement d’un bout de la cellule sécrétrice au moment du relâchement des éléments synthétisés au sein de la cellule. Des fragments de différentes tailles de la cellule aussi nommés débris cellulaires contenant des organelles (sauf le noyau) se retrouvent dans le lait (Neveu et al., 2002). En raison de ce mécanisme de sécrétion, la forte teneur en cellules anucléées rend le comptage de cellules somatiques moins précis par certaines méthodes d’analyse au laboratoire. Figure 1.2 Lait de chèvre centrifugé. (1) trois globules de gras contiennent du matériel cytoplasmique d'un côté (x 4800). (2) À plus grand grossissement (x 28 000) du matériel cytoplasmique des globules de gras, on peut voir une mitochondrie, des ribosomes et des citernes du réticulum endoplasmique rugueux. Tirée de Wooding et al. (1970). 4
1.1.2 Composition et structure des matières grasses Sous forme de petits globules ou de gouttelettes de gras, les matières grasses laitières représentent entre 3,0 à 7,2 % du lait sécrété par les cellules épithéliales de la glande mammaire chez les chèvres (Gantner et al., 2015). Le globule de gras est formé d’un noyau et d’une membrane complexe permettant de se stabiliser et de se protéger contre la lipolyse et l’oxydation dans la phase aqueuse du lait (Couvreur et Hurtaud, 2007; Bernard et al., 2018). En moyenne, les globules de gras du lait de chèvre ont un diamètre de 3,5 µm (Park et al., 2006). Cette petite taille, comparativement aux globules de gras du lait de vache (4,9 µm) et du lait des humains (4,5 µm; Yao et al., 2016), leur confère une meilleure dispersion en solution et permet une homogénéisation naturelle. D’un point de vue nutritionnel, cette caractéristique physique des matières grasses du lait de chèvre permet une meilleure digestion des gras, car l’action des lipases pancréatiques a lieu sur une plus grande surface de contact (Gantner et al., 2015; Park, 2017). Par ailleurs, leur taille et l’absence d’agglutinine, une enzyme responsable d’agglomérer les globules de gras, sont associées à une faible capacité de former de la crème à la surface du lait (Gantner et al., 2015; Park, 2017). Le globule de gras contient différents composants classifiés selon leur polarité; les lipides neutres et les lipides polaires (Rodríguez-Alcalá et Fontecha, 2010). Parmi les lipides neutres, on retrouve majoritairement des triacylglycérols (Tableau 1.1). Ces derniers sont formés de trois acides gras estérifiés à un glycérol et ils sont particulièrement présents dans le noyau du globule de gras. Quant aux lipides polaires, ceux-ci sont principalement composés de phospholipides et on les retrouve plus précisément dans la membrane du globule puisqu’ils sont d’excellents agents émulsifiants. Outre ces effets protecteurs, la membrane est responsable d’empêcher l’agrégation et la coalescence des globules (Gantner et al., 2015). Au cours des dernières années, la membrane des globules de gras a été un élément de recherche important en raison de ses composantes connues pour leurs propriétés bioactives et leurs bienfaits potentiels sur la santé humaine. Selon une revue réalisée par Spitsberg (2005), certains phospholipides ont été associés à des effets inhibiteurs du cancer du côlon, de pathogènes gastro-intestinaux, de la maladie d'Alzheimer, de la dépression et du stress. Par la taille réduite de leurs globules, les matières grasses du lait de chèvre présentent des teneurs en lipides polaires plus élevées que celles du lait de vache (0,65 contre 0,36, respectivement). 5
Tableau 1.1 Teneurs moyennes et leurs coefficients de variation (CV) des classes de lipides retrouvées dans les matières grasses laitières des vaches et des chèvres. Vache Chèvre Composant lipidique Moyenne CV Moyenne CV Lipide neutre (%) 99,64 0,01 99,35 0,02 Lipide polaire (%) 0,36a 3,94 0,65b 2,66 Lipide neutre (% de lipides totaux) Ester de cholestérol 0,04 2,70 0,04 2,46 Triacylglycéride 97,75 0,10 97,32 0,06 Cholestérol/Diglycéride/acide gras libre 1,81 6,32 1,89 2,93 Monoglycéride 0,04 2,99 0,10 9,96 Phospholipide (% des lipides polaires) Lactosylcéramide 5,10a 13,42 7,57b 6,32 Phosphatidyléthanolamine 36,58a 0,57 29,17b 8,99 Phosphatidylinositol 6,18 4,86 5,77 3,69 Phosphatidylsérine 7,28 5,71 7,65 8,65 Phosphatidylcholine 24,60 a 4,30 26,25b 3,69 Sphingomyéline 20,25 a 6,44 23,24b 6,51 a, b, c Une lettre différente est attribuée à des groupes significativement différents (p < 0,05). Adapté de Rodríguez-Alcalá et Fontecha (2010). 1.1.3 Profil en acide gras du lait Il est connu que le lait de chèvre contient des teneurs en acides gras à chaînes courte et moyenne (6:0 à 14:0) plus élevées comparativement au lait de vache et de brebis (Tableau 1.2). Cette principale différence réside dans les proportions plus grandes des acides caproïque (6:0), caprylique (8:0), caprique (10:0) et laurique (12:0; Jandal et al., 1996). Les acides gras 6:0, 8:0 et 10:0 ont ainsi été nommés d’après les caprins en raison de leur teneur importante dans leur lait (Haenlein, 2004). De plus, ils sont fortement corrélés au goût typique de « chèvre » (Park et al., 2017). Ces acides gras peuvent former entre 15-18 % de la matière grasse laitière totale par rapport à 5-9 % chez la vache et ils contribuent à sa meilleure digestibilité (Bernard et al., 2018). Selon Tziboula-Clarke (2003), ces acides gras à courte et moyenne chaîne proviendraient d’une différence dans la polymérisation de l’acétate, un acide gras volatil produit lors de la fermentation par les bactéries dans le rumen. Quant aux acides gras à chaîne impaire et ramifiée (iso et anteiso), ils se retrouvent en proportions similaires dans le lait de chèvre et de vache (Park et al., 2017). Il en est de même pour les acides gras saturés et mono-insaturés. Les acides gras saturés et mono-insaturés composent en quasi-totalité à plus ou moins 96 % la matière grasse du lait des ruminants. Par l’action de la population microbienne de leur estomac, les ruminants ont la capacité d’hydrogéner les acides gras insaturés. Ce processus sera décrit ultérieurement dans le présent chapitre. Quant à la proportion des acides gras polyinsaturés (18:2-18:3), elle est légèrement plus élevée chez les caprins de 25 % par rapport aux vaches (Castro-Gómez et al., 2014). Selon une revue 6
scientifique rédigée par Bernard et al. (2018), le lait de chèvre contient entre 59 et 74 % d’acides gras saturés et de 2,5 à 7,3 % d’acides gras polyinsaturés. Tableau 1.2 Le profil en acides gras du lait de vache, de brebis et de chèvre. Acide gras (AG), % Vache Brebis Chèvre 4:0 3,20 ± 0,23 2,47 ± 0,3 2,01 ± 0,4 6:0 2,08 ± 0,17 1,97 ± 0,19 2,39 ± 0,47 8:0 1,21 ± 0,14 1,78 ± 0,23 2,70 ± 0,4 10:0 3,05 ± 0,47 6,00 ± 0,99 9,71 ± 1,48 10:1 0,84 ± 0,16 0,52 ± 0,09 0,69 ± 0,14 12:0 3,28 ± 0,53 3,15 ± 0,53 4,26 ± 0,63 14:0 12,15 ± 0,99 11,21 ± 1 9,63 ± 0,72 14:1 1,02 ± 0,15 0,17 ± 0,03 0,14 ± 0,02 15:0 1,09 ± 0,17 0,85 ± 0,11 0,67 ± 0,1 16:0 32,19 ± 1,58 29,32 ± 2,02 27,69 ± 1,7 16:1 1,38 ± 0,15 1,04 ± 0,07 0,70 ± 0,08 17:0 0,57 ± 0,06 0,55 ± 0,2 0,51 ± 0,07 17:1 0,32 ± 0,05 0,31 ± 0,07 0,27 ± 0,05 18:0 8,93 ± 1,81 11,75 ± 1,2 9,66 ± 1,94 18:1 trans total 2,59 ± 0,81 2,39 ± 0,55 2,57 ± 0,43 18:1 cis total 22,21 ± 1,91 22,45 ± 2,57 21,7 ± 2,32 18:2 cis-9, cis-12 3,00 ± 0,91 2,73 ± 0,39 3,74 ± 0,7 18:2 cis-9, trans-11 0,38 ± 0,18 0,52 ± 0,17 0,51 ± 0,13 18:3 0,24 ± 0,09 0,53 ± 0,41 0,28 ± 0,06 20:0 0,27 ± 0,49 0,29 ± 0,09 0,17 ± 0,04 Somme AG saturés 68,02 ± 2,56 69,35 ± 2,87 69,40 ± 2,60 AG mono-insaturés 28,36 ± 2,34 26,87 ± 2,53 26,07 ± 2,36 AG polyinsaturés 3,61 ± 0,98 3,78 ± 0,76 4,53 ± 0,76 AG saturés / AG insaturés 2,15 ± 0,26 2,29 ± 0,32 2,29 ± 0,28 Selon la méthode Folch modifiée par Iverson et al. (2001). Adapté de Castro-Gómez et al. (2014). 1.2 Métabolisme des matières grasses alimentaires du rumen au prélèvement par la glande mammaire Étant un herbivore et faisant partie de la famille des ruminants, la chèvre laitière se nourrit principalement de fourrages. Sur une base de matière sèche (MS), la teneur en lipides des fourrages se situe entre 6 et 8 % sous forme de galactolipides et de phospholipides localisés principalement dans les feuilles (Harfoot, 1978). Une grande partie des acides gras retrouvés dans les plantes fourragères sont insaturés et majoritairement constitués d’acides linolénique (18:3 cis-9, cis-12, cis-15), linoléique (18:2 cis-9, cis-12) et 7
oléique (18:1 cis-9; Harfoot et Hazlewood, 1997). Le stade de maturité et l’espèce sont des éléments pouvant faire varier la teneur en lipides et le profil en acides gras des fourrages (Boufaïed et al., 2003). D’autres ingrédients importants incorporés dans les rations des chèvres laitières sont les concentrés. Cette catégorie d’aliments comprend les céréales (maïs, blé, orge, avoine, etc.), les protéagineuses (pois, fèverole), ainsi que les oléagineuses (soya, canola) et leurs dérivés (tourteaux). Les producteurs ont souvent recours aux céréales et aux oléagineuses pour augmenter la densité énergétique de la ration et aux protéagineux et tourteaux d’oléagineuses pour augmenter la teneur en protéines. Les proportions calculées de ces différents ingrédients (fourrages et concentrés) dans les rations dépendent du stade physiologique de l’animal et de ses besoins nutritionnels. Les lipides des concentrés sont différents de ceux des fourrages, étant constitués à 98 % de triacylglycérols. Dépendamment de la nature du concentré, la teneur en lipides varie. Les oléagineuses peuvent atteindre des concentrations allant jusqu’à 50 % de matière grasse (p. ex. tournesol). Les protéagineuses et les tourteaux d’oléagineuses sont composés de 1 à 10 % de matière grasse, tandis que les céréales en contiennent de plus faibles proportions, soit de 1 à 6 %. Mis à part quelques exceptions, ces ingrédients contiennent majoritairement des acides gras insaturés comme les acides linoléique, oléique, palmitique et -linolénique (Cuvelier et al., 2005). Ainsi, malgré une alimentation contenant une importante quantité d’acides gras insaturés, le lait de chèvre est plutôt riche en acides gras saturés. Ce phénomène s’explique par deux réactions biochimiques très importantes dans le rumen. D’une part l’hydrolyse des lipides alimentaires (lipolyse) et l’hydrogénation des acides gras et d’autre part, la synthèse d’acides gras par les microorganismes du rumen. 1.2.1 Lipolyse des matières grasses alimentaires Les lipides alimentaires (galactolipides, phospholipides et triacylglycérols) sont rapidement hydrolysés après leur ingestion par des lipases et des estérases produites par les bactéries et les protozoaires de la flore ruminale. Cette étape permet de libérer presque la totalité du glycérol, du galactose, du phosphore et des acides gras de ces molécules complexes (Figure 1.3; Jenkins, 1993). Les bactéries lipolytiques les plus connues sont Anaerovibrio lipolytica et Butyrivibrio fibrisolvens. Il a été rapporté que plus de 85 % des triacylglycérols sont habituellement hydrolysés, mais que plusieurs facteurs peuvent avoir des répercussions sur ce taux (Bauman et Lock, 2006). Par exemple, la lipolyse est plus élevée lorsque des rations enrichies en lipides sont offertes aux animaux et elle est plus faible quand le pH ruminal est très acide parce que l’activité et la croissance des bactéries sont réduites dans ces conditions. Après l’hydrolyse des liaisons ester, la fraction glucidique (glycérol et galactose) est rapidement fermentée en acides gras volatils (propionate et butyrate) par les bactéries. Ces derniers sont absorbés rapidement par les parois du rumen et contribuent à combler les besoins énergétiques de l’animal (NRC, 2007). 8
Vous pouvez aussi lire