Rôle des mitochondries dans le développement d'un stress oxydant dans le muscle squelettique au cours du vieillissement - Inra
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
INRA Prod. Anim.,
2006, 19 (4), 305-318 Rôle des mitochondries
dans le développement d’un stress
oxydant dans le muscle squelettique
au cours du vieillissement
F. CAPEL1, L. DEMAISON2, B. MORIO1, V. RIMBERT1, P. PATUREAU MIRAND1, L. MOSONI1
1 INRA, Université Clermont I, UMR1019 Nutrition Humaine, F-63122 Saint-Genès Champanelle, France
2 INSERM, E221, Université J. Fourier, Laboratoire de Bioénergétique Fondamentale et Appliquée,
BP 53, F-38041 Grenoble, France
Courriel : mosoni@clermont.inra.fr
La fonte musculaire observée au cours du vieillissement est une préoccupation majeure en
terme de santé publique car la population âgée ne cesse de croître dans les pays développés.
Maintenir la masse musculaire est en effet un facteur clé pour permettre une vie autonome,
active et en bonne santé pour les personnes âgées. L’implication du stress oxydant, en
particulier d’origine mitochondrial, dans ce phénomène a été analysée.
La proportion de personnes âgées ne situations d'agressions d'ordre nutri- Le stress oxydant est ainsi considéré
cesse de croître dans la population fran- tionnel, infectieux ou traumatique comme l’un des principaux facteurs
çaise et même mondiale. En 2000, la (Young et al 1989). Ceci se traduit par responsables des perturbations métabo-
France comptait 16 % de personnes de un état de santé de plus en plus précai- liques à l'origine du vieillissement. Il
plus de 65 ans, et les prévisions de re avec l'âge et par un accroissement résulte d'un déséquilibre entre l'action
l'Institut National de la Statistique et progressif de la mortalité, générant des souvent délétère et toxique des EAO
des Etudes Economiques (INSEE) pour dépenses de santé croissantes et un présentes et générées au sein de l'orga-
les années suivantes montrent que cette mal-être pour de nombreuses personnes nisme et les activités antioxydantes ou
proportion va augmenter et atteindre de plus en plus dépendantes. Il est donc de réparation. La mitochondrie serait
20 % en 2020 et 35 % en 2050. Ce phé- important de comprendre les mécanis- au cœur de ce processus car elle consti-
nomène est lié à l'amélioration des mes de la fonte musculaire de façon à tue une source importante d'EAO
conditions de vie (alimentation, hygiè- tenter de la ralentir. (Chance et al 1979). De plus, il a été
ne) et aux progrès de la médecine (mor- montré que la quantité d'EAO produites
talité infantile…). Ainsi, en 1900, Les variations de la masse des protéi- par les mitochondries était inversement
l'espérance de vie moyenne en France nes musculaires dépendent essentielle- corrélée avec la longévité des espèces
était de 47 ans. Cette espérance de vie ment de la balance entre leur synthèse et (Beckman et Ames 1998). Ces EAO
est de nos jours de l'ordre de 78 ans leur dégradation continuelle. Au cours peuvent induire des dommages oxyda-
(pour l'année 1998). Le vieillissement du vieillissement, il est possible que des tifs au sein même des structures mito-
touche donc une population de plus en altérations dans les activités de synthèse chondriales (lipides membranaires,
plus importante. Or, ce vieillissement et de dégradation des protéines muscu- Acide DésoxyriboNucléique mitochon-
se caractérise par une perte progressive laires se produisent sous l'action des drial (ADNmt), protéines de la chaîne
des fonctions physiologiques et respiratoire ou enzymes de la matrice).
psychologiques, une réduction des Espèces Actives de l'Oxygène (EAO).
capacités à maintenir l'homéostasie. On En effet, il a été montré dans de nom- Une altération des structures mito-
constate en particulier l'apparition de breux tissus et organes, dont le muscle, chondriales contribue à perturber le
façon relativement précoce d'une l'existence d'une accumulation de pro- fonctionnement de la chaîne respiratoi-
réduction de la masse des protéines téines altérées, principalement du fait re, risquant de provoquer une limitation
musculaires (sarcopénie). En moyenne, d'oxydations radicalaires, mais aussi à de la production d'énergie dans la cellu-
l'homme perd 11 kg de masse muscu- la suite de la fixation non enzymatique le et une aggravation du stress oxydant.
laire corporelle entre 20-29 ans et 70- de sucres (glycations), ou autres modifi- Ces interactions entre stress oxydant et
79 ans (Cohn et al 1980). Ce déclin cations post-traductionnelles. Ceci peut fonctionnement mitochondrial pour-
contribue à limiter la mobilité des per- s'accompagner à la fois d'une altération raient avoir des conséquences particu-
sonnes âgées, mais il aggrave aussi leur de la fonctionnalité des protéines oxy- lièrement importantes dans des tissus à
vulnérabilité. En effet, le muscle cons- dées et d'une modification du potentiel faible renouvellement cellulaire
titue un véritable tissu de secours dans protéolytique musculaire et jouer un comme le muscle, qui a pourtant été
lequel l'organisme va puiser dans des rôle dans la fonte musculaire. très peu étudié.
INRA Productions Animales, Octobre 2006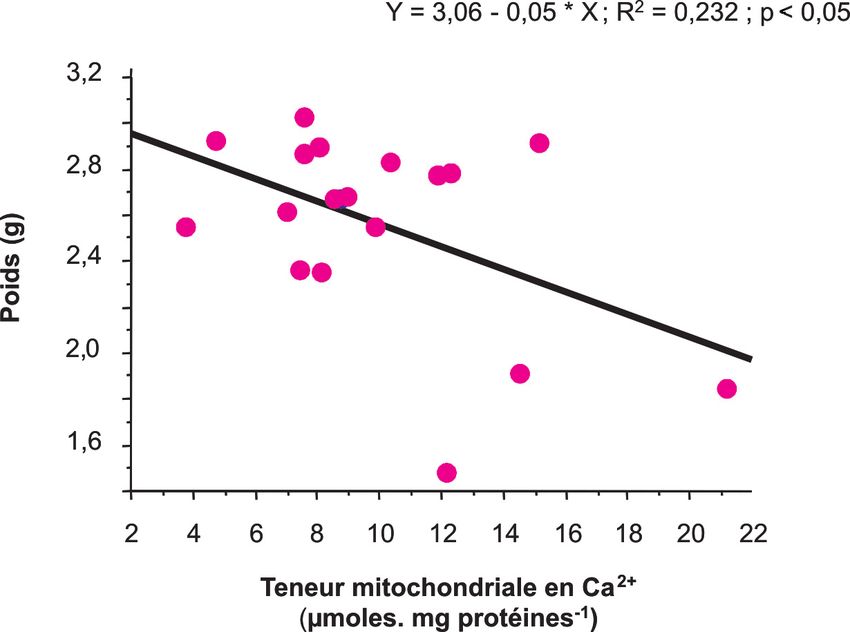
306 / F. CAPEL et al
Ces travaux ont donc été entrepris biopsie du vastus lateralis sous anes- se), et l’autre utilisée comme index de
pour déterminer si un défaut de contrô- thésie locale chez l’homme, ou sur le la teneur en calcium mitochondrial
le du stress oxydant et notamment de la muscle entier chez le rat. La détermina- (activité isocitrate deshydrogénase).
production mitochondriale d'EAO exis- tion d'une capacité de production
te dans le muscle squelettique au cours d'EAO représentative des mitochon- La chaîne mitochondriale de
du vieillissement. Cela devrait permet- dries intégrées dans la structure muscu- transport d'électrons génère de l'anion
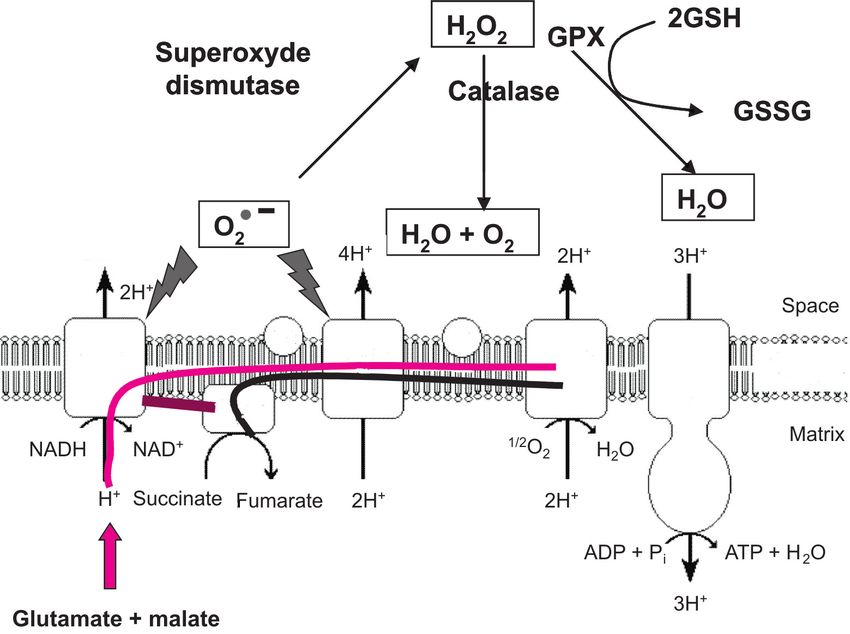
tre de déterminer si ce phénomène laire impose d'obtenir en moins de superoxyde O2•- (figure 1) qui peut être
intervient dans le développement de la deux heures des mitochondries dont les converti en H2O2 (peroxyde d’hydrogè-
sarcopénie, et quels sont les mécanis- membranes sont aussi intactes que pos- ne) grâce à l’enzyme antioxydante
mes impliqués. sible. Ainsi une technique permettant appelée superoxyde dismutase et pré-
sente dans la mitochondrie (forme Mn
d'obtenir des mitochondries présentant SOD) ou dans le cytosol (forme Cu-Zn
de très bonnes capacités respiratoires SOD). A l'inverse de l'O2•-, l'H2O2 est
1 / Comment évaluer l’ori- (Rasmussen et al 1997) a été adaptée à plus stable et diffuse à travers les mem-
notre étude. Outre la respiration, nous
gine, l’intensité et les consé- avons analysé le fonctionnement des
branes mitochondriales. Nous avons
mesuré la production d'H2O2 grâce à
quences du stress oxydant mitochondries isolées par la détermina- une sonde dont la fluorescence varie en
dans le muscle ? tion d’activités enzymatiques spéci- fonction de son état d’oxydation après
fiques. Nous avons mesuré d’une part réaction avec l’H2O2 présent après
des activités enzymatiques impliquées addition de peroxydase. Après avoir
dans le transport des électrons le long utilisé la scopolétine dans une première
L’étude de l'effet du vieillissement de la chaîne respiratoire lors de la respi- étude, nous avons retenu le DCFH-DA
sur le fonctionnement mitochondrial et ration (somme des activités des com- par la suite. L'échantillon de mitochon-
le stress oxydant dans le muscle a été plexes II et III, activité cytochrome c dries, source d'H2O2, est incubé sous
réalisée à la fois chez le rat et chez oxydase - complexe IV), deux enzymes agitation permanente, à température
l’homme, en comparant des sujets du cycle de Krebs, l’une souvent utili- constante en présence de substrats du
expérimentaux sains, adultes ou âgés. sée comme marqueur de la densité métabolisme mitochondrial (glutamate,
Les mesures ont été effectuées après mitochondriale (activité citrate syntha- malate, succinate) dans un état de respi-
Figure 1. Représentation schématique des flux d’électrons à travers les complexes de la chaîne respiratoire mitochondriale, de la
production d’anion superoxyde, et des défenses antioxydantes cellulaires.
CoQ Complex Cyt Complex
Complex III C
I IV Inner
Complex Mitochondrial
V Menbrane
Complex
II
A partir de l’oxydation des substrats du complexe I (glutamate ou malate, ––) ou du complexe II (succinate, ––, les électrons parcourent les diffé-
rents complexes de la chaîne respiratoire mitochondriale, créant un gradient de protons. Ce gradient est utilisé pour synthétiser de l’ATP(Adénosine
triphosphate) à partir d’ADP(Adénosine DiPhosphate) au niveau du complexe V. On parle de couplage entre le gradient de proton et la synthèse
d’ATP. Il existe également un flux inverse d’électron du complexe II vers le complexe I ––. Lors de ces transferts d’électrons, des fuites se produi-
sent, aboutissant à la formation d’anion superoxyde O2•-. Cet anion dismute spontanément ou sous l’action de la superoxyde dismutase en H2O2.
Cette forme oxydante est alors prise en charge par la glutathion peroxydase (GPX) à l’aide de glutathion réduit (GSH) qui est alors oxydé (GSSG)
ou par la catalase, et est détoxifiée en eau.
INRA Productions Animales, Octobre 2006Rôle des mitochondries dans le développement d’un stress oxydant dans le muscle squelettique... / 307
ration basale en l’absence d’ADP (état qu'elle est convertie spontanément ou exhaustive des mitochondries muscu-
IV), pendant lequel la production grâce à la superoxyde dismutase (SOD) laires malgré le traitement par la subti-
d'EAO est maximale (Barja 1999). en H2O2 qui peut diffuser hors de la lisine.
Puisque certains couples redox inter- mitochondrie et conduire à la formation
agissent avec le système de détection, d'EAO plus néfastes (radical hydroxy- Chez les rats de 4,5 mois, les mito-
nous avons choisi et adapté un protoco- le) par réaction avec d'autres molécules chondries du muscle tibialis anterior
le de mesure de l'H2O2 produit par les telles que le fer. L'évolution avec l'âge avaient tendance à produire plus
mitochondries après les avoir éliminées de la production mitochondriale d'EAO d'H2O2 que les mitochondries du mus-
à la fin de l'incubation (Staniek et Nohl dans le muscle squelettique n'a été que cle soleus (+ 155 %, p = 0,08). Il n’y
1999). Cette technique permet de limi- très peu étudiée et les résultats contra- avait plus de différence à 24 mois. En
ter les interactions non spécifiques et de dictoires (Bejma et Ji 1999, Drew et al effet, dans le soleus, la production
détecter la libération nette d’H2O2 hors 2003) qui ont été rapportés, ne permet- mitochondriale d'H2O2 augmentait
de la mitochondrie. Des gammes d'éta- tent pas de savoir quel est l'effet du significativement avec l'âge (+ 208 %
lonnage d'H2O2 ont été préparées dans vieillissement et s'il est différent en entre 4,5 et 24 mois) et rejoignait le
des conditions identiques à celles des fonction du type musculaire. Cette niveau observé dans le tibialis anterior
échantillons (absence de mitochon- étude a donc été entreprise pour compa- qui ne variait pas avec l’âge
dries, mêmes tampons et substrats). rer l'évolution au cours du vieillisse- (figure 2A). L'addition d'antimycine A,
ment de la production mitochondriale inhibiteur du flux d’électron au niveau
Nous avons analysé l’intensité des d'H2O2 et de ses mécanismes en fonc- du complexe III stimulait la production
dommages oxydatifs présents au tion du type musculaire. Ses consé- d'H2O2, les valeurs obtenues étant iden-
niveau mitochondrial en mesurant la quences sur diverses activités du méta- tiques chez les animaux jeunes et âgés
teneur en groupements carbonyles des bolisme mitochondrial et sur le stress (figure 2B). Cette stimulation par l’an-
protéines mitochondriales à l'aide de oxydant ont été recherchées en exami- timycine A est le signe d’un rôle du
tétrahydroborure de sodium tritié nant l'évolution de la teneur en dérivés complexe III dans la production mito-
(NaB3H4). carbonylés des protéines mitochondria- chondriale d’EAO. Ce rôle semblait
les et de l'activité des enzymes antioxy- inchangé avec l’âge.
Enfin, nous avons complété notre dantes majeures (SOD, catalase, GPX
étude du stress oxydant musculaire par et GRed). Les travaux ont été réalisés Lorsque les électrons provenaient du
la mesure des activités des enzymes avec des rats mâles Wistar jeunes complexe II (en présence du substrat
antioxydantes (figure 1). Nous avons (4,5 mois) et âgés (24 mois), élevés à adéquat, le succinate) et étaient orien-
ainsi déterminé 1) les activités SOD l'animalerie présente sur le site INRA tées uniquement vers le flux normal
cytosoliques (Cu-Zn SOD) et mito- de Theix (Unité Expérimentale de d’électrons vers le complexe III (en
chondriales (Mn SOD), capables de Nutrition Comparée). Les muscles étu- présence d’un inhibiteur qui bloque le
dismuter l'anion superoxyde O2•- pro- diés étaient 2 muscles de la patte arriè- flux inverse d’électrons possible du
duit en H2O2 ; 2) l’activité glutathion re ayant des propriétés métaboliques complexe II vers le complexe I, la roté-
réductase (GRed), qui permet de rédui- différentes : le soleus, un muscle oxy- none), il n’y avait également aucun
re le glutathion oxydé et donc de res- datif et le tibialis anterior, un muscle effet de l’âge sur la production mito-
taurer l’antioxydant majeur qu’est le mixte à prédominance glycolytique. chondriale d'H2O2 (figure 3).
glutathion ; 3) l’activité glutathion per- L’addition d’antimycine A stimulait
oxydase (GPX) qui, en utilisant La masse du tibialis anterior était très fortement celle-ci aux deux âges,
comme co-facteur le glutathion, permet significativement réduite avec l'âge confirmant le rôle du complexe III dans
de détoxifier l’ H2O2 en eau ; 4) l’acti- (- 23 %), alors que celle du soleus était la production mitochondriale d’EAO.
vité catalase qui détoxifie également stable, confirmant l'atrophie plus Nous avons également vérifié que cette
l’H2O2. importante des muscles glycolytiques production ne se situait pas au niveau
au cours du vieillissement. du complexe II (l'ajout de TTFA, inhi-
2 / L’évolution de la pro- biteur spécifique du complexe II indui-
Les paramètres respiratoires mesurés sait une diminution de la quantité
duction mitochondriale sur les mitochondries isolées déno- d'H2O2 produite par les mitochondries
d'H2O2 dans les muscles taient un bon état de fonctionnement, et en présence de succinate).
n'étaient pas affectés par le vieillisse-
oxydatifs et glycolytiques ment. De même, aucune variation Face à cette production mitochon-
chez le rat (Capel et al 2004) significative des rendements d'extrac- driale d’EAO, les taux d'oxydation des
tion (mg de protéines mitochondriales protéines mitochondriales n'étaient pas
par g de tissu) n'a été constatée en fonc- différents aux 2 âges dans les 2 mus-
Lors du transfert normal des tion de l'âge ou du muscle, et l’activité cles. En revanche, ils tendaient
électrons au sein des complexes de la cytochrome C oxydase était maintenue (p < 0,065) à être supérieurs dans le
chaîne respiratoire mitochondriale au dans le tibialis anterior. En revanche, tibialis anterior par rapport au soleus,
cours de la respiration (figure 1), une l'activité de la citrate synthase était plus quel que soit l'âge, indiquant donc un
fuite d’électrons se produit aux niveaux faible chez les rats de 24 mois que chez niveau d'oxydation de cette fraction
des complexes I et III (Barja 1999). Le ceux de 4,5 mois. Ce dernier résultat protéique plus important dans les mus-
même phénomène a lieu lors du flux pourrait signifier que le nombre de cles glycolytiques que dans les muscles
inverse d’électrons qui se produit éga- mitochondries baisse dans le muscle oxydatifs. Nous avons aussi constaté
lement entre les complexes II et I (Liu avec l'âge. L'importance de cette baisse que les activités Cu-Zn SOD (forme
et al 2002, St-Pierre et al 2002). Cette (- 16 %) n'était probablement pas cytosolique) et catalase étaient plus éle-
fuite provoque la capture d’un électron suffisante pour avoir un effet détectable vées dans le tibialis anterior des rats de
par l’oxygène O2 qui devient ainsi une sur le rendement de l'isolement des 24 mois que chez ceux de 4,5 mois
EAO : O2•-. Cette forme radicalaire a mitochondries. De plus, la méthode (figure 4). A l'inverse, chez les animaux
une demi-vie extrêmement brève, puis- retenue ne permet pas une extraction de 24 mois, l'activité GPX était plus
INRA Productions Animales, Octobre 2006308 / F. CAPEL et al
Figure 2. Effet du type musculaire et du vieillissement chez le rat sur la production faible (figure 5), tandis que l'activité de
mitochondriale d’H2O2 en présence de glutamate / malate avec (B) ou sans (A) antimy- la forme mitochondriale des SOD (Mn
cine. SOD) restait stable (figure 4).
Soleus Ainsi, ce travail a permis de montrer
Tibialis que les muscles oxydatifs et glycoly-
tiques sont affectés différemment par le
stress oxydant au cours du vieillisse-
ment. Chez le jeune animal, les mito-
chondries produisent plus d'H2O2 dans
les muscles glycolytiques que dans les
muscles oxydatifs, ce qui est cohérent
avec la tendance à observer plus de pro-
duits de l'oxydation des protéines mito-
chondriales dans ce type musculaire et
avec la sensibilité des muscles glycoly-
tiques à la sarcopénie. C'est également
cohérent avec le fait que les muscles
glycolytiques constituent un environne-
ment chimiquement plus réduit que les
muscles oxydatifs.
En revanche, même si la production
d'H2O2 augmentait avec l'âge dans le
muscle soleus, il semblerait que cela ait
un impact négligeable sur la sarcopé-
Les valeurs indiquées sont significativement différentes des valeurs obtenues dans le soleus des
jeunes rats : * PRôle des mitochondries dans le développement d’un stress oxydant dans le muscle squelettique... / 309
Figure 5. Effet du vieillissement sur les activités glutathion peroxydase (GPX) et gluta- dans le muscle squelettique (Liu et al
thion réductase (GRed) dans le muscle tibialis anterior chez le rat. 2002, Servais et al 2003), nous avons
choisi comme deuxième condition
expérimentale le même mélange de sub-
strats, additionné de roténone. Ainsi,
dans nos conditions expérimentales,
une inhibition de la production mito-
chondriale d'EAO par la roténone serait
une indication d'une production majori-
tairement liée au flux inverse, une sti-
mU/mg protein
mU/mg protein
mulation serait le signe d'une produc-
tion liée au flux normal et une absence
d'effet ne permettrait pas de conclure
sur les mécanismes de production.
Puisque l'activité physique et l'état
nutritionnel sont des paramètres qui
peuvent influencer la fonction mito-
chondriale, les volontaires, répartis en
deux groupes âgés de 23,5 ans et de
67 ans (n = 6 par groupe) ont été sélec-
tionnés de manière à ce qu'ils aient un
* : Valeur significativement différente de la valeur obtenue chez les jeunes rats (P310 / F. CAPEL et al
Tableau 2. Effet du vieillissement sur les activités enzymatiques mitochondriales mus- Cette augmentation de la libération
culaires chez l'homme. mitochondriale d'H2O2 pourrait égale-
ment être un signal d'adaptation pour la
cellule ou la mitochondrie. Les EAO
régulent en effet plusieurs fonctions
cellulaires, dont le métabolisme du cal-
cium ou la réponse au stress.
4 / La lidocaïne peut-elle
interférer avec les mesures
Abréviations : CS, citrate synthase ; COX, cytochrome c oxidase ; NS, non significatif.
de la production d’EAO sur
les biopsies humaines ?
Le vieillissement n'affectait pas la portance de ce flux est accrue avec l'âge
capacité des mitochondries à oxyder les et ne s'oppose pas aux données obte-
acides gras, ni les activités des com- nues chez l'homme ou l'animal. La réalisation d’une biopsie muscu-
plexes II à IV de la chaîne respiratoire laire chez l’homme nécessite une anes-
et de la citrate synthase (tableau 2), Pourtant, en accord avec d’autres étu- thésie locale. Dans notre étude, nous
mais il induisait une augmentation des réalisées chez l’homme avons utilisé une injection sous-cutanée
importante (+ 175 %) de la production (Rasmussen et al 2003, Rimbert et al de lidocaïne (Capel et al 2005a). Or, ce
d'H2O2 (figure 6). En présence de roté- 2004), les capacités oxydatives muscu- produit est capable de perturber la
none, cette production était inhibée laires semblaient préservées chez les respiration mitochondriale au même
d'environ 70 % dans les 2 groupes et il sujets âgés. Les conséquences de la sti- titre que les inhibiteurs que nous utili-
n'y avait plus de différence significati- mulation de production d'H2O2 liée au sons ; il a des propriétés découplantes
ve entre les 2 groupes d'âge, malgré une flux inverse d'électrons restent donc à (Dabadie et al 1987, Sztark et al 1998).
production qui tendait à demeurer préciser. Il n'est pas évident que les Il est donc susceptible d’interférer avec
supérieure dans le groupe de volontai- mitochondries subissent des altérations les mesures réalisées au niveau mito-
res âgés (+ 130 %, p = 0,11) (figure 6). significatives, malgré la proximité de la chondrial. Pour tester ceci, nous avons
source d'EAO. Les altérations pour- répété chez le rat les mesures réalisées
Ainsi, ces résultats indiquent que la raient être spécifiques, touchant certai- chez l’homme. Pour tester l’effet de la
production d'H2O2 augmente avec l'âge nes molécules particulièrement sensi- lidocaïne, nous avons injecté ce produit
dans le muscle vastus lateralis humain. bles. En outre, il est possible que des en intramusculaire au niveau du muscle
Cette augmentation est vraisemblable- effets délétères apparaissent seulement gastrocnemius de la patte droite de
ment due au flux inverse d'électrons, chez des individus encore plus âgés que l'animal (1mL d'une solution de lidocaï-
puisque la roténone réduisait la libération ceux qui étaient inclus dans ces tra- ne 2 %), et nous avons comparé la
d'H2O2 et masquait l'effet du vieillisse- vaux. D'autres études seront nécessai- respiration mitochondriale et la produc-
ment. En effet, il a été montré dans le res pour vérifier cette hypothèse. tion d’H2O2 des mitochondries isolées
muscle : 1) que la roténone inhibe la pro-
duction d'EAO par des mitochondries Figure 6. Effet du vieillissement sur la production mitochondriale d’H2O2 dans le mus-
respirant en présence de succinate cle vastus lateralis humain en présence de glutamate/malate/succinate avec ou sans
(Servais et al 2003) grâce à l'inhibition roténone.
du flux inverse d'électrons ; 2) que la
production mitochondriale d'EAO en
présence de substrats du complexe I est
stimulée par la roténone (St-Pierre et al
2002). L'effet inhibiteur de la roténone
est clairement prédominant par rapport à
l'effet stimulateur dans nos conditions.
La grande majorité de l'H2O2 produit en
l'absence de roténone était donc due au
flux inverse d'électrons.
Une seule autre étude a décrit la pro-
duction mitochondriale d'EAO dans le
muscle chez l'homme âgé avec des sub-
strats du complexe I. Elle n'a pas mis en
évidence d'effet du vieillissement
(Tonkonogi et al 2003). Chez l'animal,
l'analyse de l'effet du vieillissement
dans le muscle sur cette production a
été réalisée avec des substrats des com-
plexes I ou II (en présence de roténo-
ne). L'ensemble des travaux antérieurs
ne permettait donc pas d'étudier le rôle
du flux inverse. Notre étude met en évi- * Significativement différent des jeunes (P < 0,05).
dence pour la première fois que l'im- # Significativement différent du basal (P < 0,05).
INRA Productions Animales, Octobre 2006Rôle des mitochondries dans le développement d’un stress oxydant dans le muscle squelettique... / 311
à partir des muscles droit (avec lidocaï- Tableau 3. Contribution des différents composants cellulaires à la production totale
ne) et gauche (sans lidocaïne). Il s’agit d'H2O2, dans le foie. MAO : Monoamines oxydases (Chance et al 1979).
bien sûr d’un modèle un peu extrême
car le volume injecté est important,
l’injection se fait en intramusculaire,
alors que chez l’homme nous avons
veillé à réaliser plusieurs petites injec-
tions en sous-cutané dans la zone pré-
vue de biopsie.
Les résultats obtenus ont montré
qu’en l’absence d’ADP (état IV), l'in-
jection de lidocaïne stimulait la res-
piration en présence de succinate. Elle avec le même objectif, mais en nous Les rats, des mâles Wistar âgés de
inhibait l'augmentation de la consom- intéressant cette fois-ci à la production 6 mois ou de 22 mois provenaient de
mation mitochondriale d'oxygène nor- globale musculaire d'EAO, et en parti- l'Unité Expérimentale de Nutrition
malement provoquée par la roténone en culier à celle qui n'est pas d'origine Comparée de l'INRA de Theix. Le mus-
présence de succinate dans le muscle mitochondriale. Il existe très peu de cle gastrocnemius a été prélevé et utili-
témoin. En présence d’ADP (état III), données concernant la répartition de la sé immédiatement pour les mesures de
aucun effet significatif de la lidocaïne production cellulaire d'EAO dans le production d'H2O2. Nous avons souhai-
n'était détecté, à part une tendance de la muscle squelettique et l'effet du té obtenir un extrait de muscle total,
consommation d'oxygène à diminuer vieillissement sur cette répartition n'a dépourvu des gros débris cellulaires et
en présence de glutamate et de malate jamais été étudié. Même si l'on admet des protéines myofibrillaires et un
(p = 0,06). Cette injection provoquait couramment que les mitochondries extrait de muscle appauvri en mito-
également une réduction de la produc- sont la source principale d'EAO de la chondries. Nous avons procédé comme
tion d'H2O2 en présence des 3 substrats cellule, il est possible que cela ne soit pour l'isolement des mitochondries par
et interagissait significativement avec pas vrai dans tous les tissus et/ou espè- centrifugation différentielle. Nous
l'effet de la roténone : dans le groupe ces animales. En effet, dans la principa- avons mesuré la production d'H2O2 par
le revue bibliographique (Chance et al les surnageants obtenus lors des centri-
témoin, la roténone inhibait la produc-
1979) qui est généralement citée pour fugations à 1000 g (fraction totale) et
tion mitochondriale d'H2O2 de 48 %, 4400 g (fraction appauvrie en mito-
alors que dans le groupe traité, elle n'a- étayer l'affirmation selon laquelle les
mitochondries sont la source principale chondries). Nous avons également
vait plus d'effet significatif. Cet effet mesuré la teneur en groupements car-
pourrait être la conséquence de l’effet d'EAO, il est indiqué que, dans le foie,
les mitochondries représenteraient seu- bonyles des protéines dans ces surna-
découplant de la lidocaïne, capable de geants 1000 g et 4400 g.
réduire le potentiel membranaire au lement 15 % de la production totale
niveau mitochondrial. d'H2O2 (tableau 3). Les peroxysomes
produisent de l'H2O2 lors de la β-oxy- Comme précédemment, la masse des
dation des acides gras et possèdent des muscles tibialis anterior et gastrocne-
Ainsi, nous avons montré que la lido- mius était significativement plus faible
caïne peut effectivement interférer avec oxydases comme sources potentielles.
Les microsomes peuvent oxyder le chez les rats de 22 mois que chez ceux
nos mesures de production d’ H2O2 en de 6 mois. En revanche le poids du
masquant l’effet inhibiteur de la roténo- NADPH grâce à la NADPH-cytochro- muscle soleus (métabolisme oxydatif)
ne. Cependant, l'inhibition marquée de me c reductase et possèdent des cyto- n'était pas significativement différent
la production mitochondriale d'H2O2 chromes P450, intervenant dans la aux 2 âges.
par la roténone que nous avons obtenue détoxication des xénobiotiques. Il exis-
lors de l’expérience réalisée chez te en plus des sources présentées dans Dans le gastrocnemius, la production
l’homme indique bien que la lidocaïne ce tableau une production d'EAO occa- d'H2O2 (figure 7) exprimée en nmol.
n'a pas interféré. Cela est certainement sionnelle, par exemple par des cellules min-1. mg prot-1 augmentait significati-
lié aux précautions prises lors de l’in- phagocytaires qui sont des sources vement avec l'âge dans le surnageant
jection en veillant à réaliser de multi- importantes d'EAO et qui peuvent total (+ 102 %, p = 0,05) et dans le sur-
ples injections sous-cutanées. La lido- s’être infiltrées dans le tissu musculaire nageant appauvri en mitochondries
caïne est donc un élément capable dans certaines situations physiopatho- (+ 170 %, p < 0,05). Lorsque cette pro-
d’interférer avec les mesures, mais ce logiques. Nous avons montré précé- duction est exprimée par gramme de
problème peut être évité. demment que les activités de la Cu-Zn muscle, l'augmentation liée à l'âge restait
SOD et de la catalase étaient plus sti- significative pour le surnageant appauvri
5 / L’augmentation avec mulées avec l'âge dans le muscle tibia- en mitochondries. Du fait d'une variabili-
lis anterior chez le rat que celle de la té importante et d'une tendance à une
l'âge de la production Mn SOD mitochondriale. Ceci pourrait baisse avec l'âge de la teneur en protéi-
non mitochondriale d'H2O2 être la conséquence d'un accroissement nes, ce n'était plus le cas dans le surna-
au cours du vieillissement de la libéra- geant total. Cela suggère que la contribu-
dans le muscle gastrocne- tion d'EAO dans le cytoplasme par tion des mitochondries à l'augmentation
mius chez le rat d'autres organites cellulaires que les de la production tissulaire d'EAO en
mitochondries. fonction de l'âge est moins importante
que celle des autres sources.
Dans nos expériences précédentes, Nous avons donc comparé la produc-
nous avons analysé l'évolution avec tion d'H2O2 par un extrait musculaire La teneur en dérivés carbonylés sur
l'âge de la production mitochondriale total ou par un extrait appauvri en mito- les protéines, marqueurs de leur oxyda-
d'EAO dans le cadre du vieillissement chondries obtenu chez des rats adultes tion, n'évoluait pas avec l'âge quelle
musculaire. Cette étude a été initiée ou âgés. que soit la fraction considérée.
INRA Productions Animales, Octobre 2006312 / F. CAPEL et al
Figure 7. Effet du vieillissement sur la production d'H2O2 dans un extrait musculaire (gastrocnemius) total (surnageant 1000 g) ou
appauvri en mitochondries (surnageant 4400 g).
* : Effet âge significatif à P < 0,05.
Nos résultats montrent donc que la particulières soient oxydées au cours ment du stress oxydant dans une situa-
production d'H2O2 augmente avec l'âge du vieillissement comme cela a été ob- tion particulière comme le vieillisse-
dans les 2 fractions étudiées. Ceci est servé dans le plasma de rats et de sou- ment et la fonte musculaire. Parmi eux,
en accord avec une étude antérieure ris (Jana et al 2002) ou chez la droso- le stress calcique semble pouvoir jouer
réalisée chez le rat dans un homogénat phile (Yan et al 1997). un rôle puisque des travaux indiquent
complet du muscle vastus lateralis que le calcium favorise la production
(Bejma et Ji 1999) bien que les condi- Cette augmentation de la production mitochondriale d'EAO (Kowaltowski
tions d'incubation soient différentes des non mitochondriale d'H2O2 est cohé- et al 2001). Au cours du vieillissement,
nôtres. Puisque nous observons une rente avec les niveaux d'activités de la les altérations rapportées au niveau des
augmentation avec l'âge de la produc- Cu-Zn SOD et de la catalase plus éle- pompes SERCA du Réticulum
tion d'H2O2 dans le surnageant appau- vés dans les muscles de rats âgés par
Sarcoplasmique (RS) (Viner et al 1999,
vri en mitochondries, et comme elle est rapport à ceux trouvés dans les muscles
de jeunes adultes. Il semble donc Carmeli et al 2002) pourraient favori-
plus marquée que dans la fraction tota- ser l'accumulation de Ca2+ dans le cyto-
le, il est vraisemblable que la produc- nécessaire de prendre en compte cette
observation lors des études d'interven- plasme. Les mitochondries sont alors
tion d'H2O2 soit augmentée au cours du susceptibles d'accroître leur prélève-
vieillissement dans les peroxysomes ou tion visant à réduire le niveau de stress
oxydant dans le muscle squelettique. ment de Ca2+ pour rétablir l'équilibre
les microsomes. Ces 2 types d'organites des concentrations en Ca2+ dans les dif-
contribuent majoritairement, avec les férents compartiments. Ce travail a
mitochondries, à la production cellulai- Ce protocole ne permettait pas de
comparer précisément les productions donc été réalisé pour déterminer si la
re d'H2O2 dans le foie (Chance et al
1979). Cependant, il n'existe pas, à d'H2O2 dans les deux surnageants car relation entre les variations des concen-
notre connaissance, de données dans la leurs compositions étaient différentes. trations mitochondriales en Ca2+ et leur
littérature se rapportant au muscle ou à Toutefois, au vu des résultats obtenus, il production d'EAO existe in vivo dans le
l'effet de l'âge. semble que les mitochondries ne soient muscle squelettique et si une augmen-
pas nécessairement la source cellulaire tation de la teneur mitochondriale en
Malgré l'augmentation de la produc- majoritaire d'H2O2 dans le muscle Ca2+ pourrait favoriser la sarcopénie
tion d'H2O2, on n'observe pas en paral- comme cela est communément admis. avec l'âge (Capel et al 2005b). Pour
lèle une augmentation de la proportion cela, nous avons utilisé un modèle ani-
de protéines carbonylées. Ces résultats 6 / La stimulation de la mal basé sur le traitement de rats avec
sont en accord avec la stabilité au cours un ionophore calcique, le A23187.
du vieillissement du niveau d'oxydation production mitochondriale Cette molécule permet l'entrée massive
des protéines totales observée chez le d'H2O2 in vivo par le Ca2+ de Ca2+ dans les cellules et induit une
rat (Mosoni et al 2004). Pourtant, d'au- dans le muscle gastrocne- entrée de Ca2+ dans les mitochondries.
tres études ont mis en évidence une
augmentation avec l'âge du niveau mius chez le rat âgé Cette étude a été conduite en 2 par-
d'oxydation dans la fraction soluble des ties. Dans chaque cas, 2 muscles ont été
protéines musculaires chez le rat étudiés, le gastrocnemius (métabolisme
(Reznick et Packer 1994). Il est possi- Il est important d’identifier les méca- mixte) et le soleus (métabolisme oxy-
ble que seulement certaines protéines nismes pouvant relier le développe- datif).
INRA Productions Animales, Octobre 2006Rôle des mitochondries dans le développement d’un stress oxydant dans le muscle squelettique... / 313
La première partie de notre travail du A23187 chez les animaux âgés uni- tration matricielle en Ca2+ (Denton et
devait permettre de vérifier l'effet du quement, alors que le vieillissement al 1978). Le dosage a été possible uni-
traitement au A23187 injecté par voie restait sans effet. En outre, une corréla- quement pour le muscle gastrocnemius,
intrapéritonéale chez des rats mâles tion inverse entre la teneur mitochon- en raison de la trop faible quantité de
Wistar âgés de 6 et 23 mois, d'une part, driale en Ca2+ et la masse musculaire mitochondries extraites du soleus.
sur les teneurs mitochondriales en Ca2+ était observée pour les 2 muscles (figu- L'activation de 40 % de l'activité ICDH
et, d'autre part, sur la masse musculai- re 8). Ceci suggère un lien possible observée dans les mitochondries des
re. L'incorporation mitochondriale de entre masse musculaire et teneur mito- animaux traités par rapport aux témoins
Ca2+ a été évaluée à l'aide d'une métho- chondriale en Ca2+. indique que la concentration mitochon-
de radioactive (Capel et al 2005b). driale en Ca2+ a bien été augmentée
Chez les animaux traités et par compa- Dans la deuxième partie de ce travail, suite à l'injection de A23187. Cette
raison aux témoins, il s'est avéré que 48 nous avons reproduit le modèle chez augmentation de l'activité ICDH n'était
heures après l'injection, les teneurs des animaux âgés de 21 mois avec pour pas liée à un accroissement global du
mitochondriales en Ca2+ étaient in- objectif d'analyser les effets du A23187 métabolisme mitochondrial car les acti-
changées dans le muscle gastrocnemius sur la production mitochondriale vités citrate synthase et COX sont res-
chez rats de 6 mois, et tendaient à aug- d'EAO avant l'apparition de la fonte tées stables dans la même fraction
menter chez les rats de 23 mois. Dans musculaire. Pour cela, les mesures ont mitochondriale.
le muscle soleus, ces teneurs étaient été effectuées 24 h après l'injection du
inchangées pour les rats de 6 mois et A23187 avec une dose réduite de 20 % Dans les conditions fixées pour l'in-
réduites chez les rats de 23 mois. Ainsi, afin d'éviter toute mortalité. Les mus- cubation des mitochondries isolées, la
une interaction significative a été cles soleus et gastrocnemius ont été production mitochondriale d'H2O2
détectée entre les effets de l'âge et du prélevés, les mitochondries isolées, et augmentait dans le groupe traité par
traitement : avec l'âge, la teneur mito- leur production d'EAO a été évaluée en rapport au groupe témoin dans le gas-
chondriale en Ca2+ augmentait chez les dosant l'H2O2 produit en présence de trocnemius et restait stable dans le
rats témoins, mais était réduite chez les glutamate + malate comme substrats de soleus (figure 9). Ce résultat qui a été
rats traités. En outre, les teneurs mito- la chaîne respiratoire. Des mesures de observé en l'absence ou en présence
chondriales en Ca2+ étaient plus éle- consommation d'oxygène par oxygra- d'antimycine A (inhibiteur du complexe
vées dans le muscle soleus que dans le phie ont été réalisées pour contrôler les III), est cohérent avec la stimulation de
gastrocnemius. capacités respiratoires des mitochon- l'activité de la Mn SOD totale dans le
dries. Aucun effet du traitement sur le gastrocnemius, et avec la baisse de l'ac-
En ce qui concerne l'effet du A23187, rapport de contrôle respiratoire et sur le tivité de la GPX qui témoigne de la
sur les masses musculaires, il a été ob- rapport ADP/O n'a été trouvé. Il a seu- réduction des capacités mitochondria-
servé que les rats de 23 mois étaient lement été observé que les capacités les à éliminer l'H2O2. Les activités de la
plus sensibles au traitement au A23187 respiratoires maximales étaient plus catalase et de la Cu-Zn SOD étaient
que les rats de 6 mois. Le poids du mus- importantes dans le gastrocnemius. inchangées suite au traitement.
cle gastrocnemius (exprimé en pour-
centage du poids corporel) était signifi- L'activité isocitrate déshydrogénase L'estimation des teneurs en protéines
cativement réduit au cours du (ICDH) a été mesurée en guise de mar- carbonylées, n'a pas mis en évidence
vieillissement, mais n'était pas affecté queur de l'évolution de la concentration d'influence de l'augmentation de la pro-
par le traitement quel que soit l'âge. A mitochondriale en Ca2+ dans la fraction duction d'H2O2 sur le niveau d'oxyda-
l'inverse, le soleus subissait une atro- de mitochondries isolées car elle est sti- tion des protéines mitochondriales.
phie significative de 17 % sous l'effet mulée par l'augmentation de la concen- Le nombre de mitochondries a été
réduit sous l'effet du traitement, puis-
Figure 8. Relation entre la teneur mitochondriale en Ca2+ et la masse du gastrocne- qu'une diminution importante (- 50 %)
mius chez le rat de 6 ou 23 mois, traité ou non avec le A23187. de l'activité citrate synthase était obser-
vée dans l'homogénat musculaire utili-
sé pour l'extraction des mitochondries
(Capel et al 2005b). Cette observation
n'était pas confirmée par la comparai-
son des rendements d'extraction mito-
chondriale (2,40 ± 0,36 et 2,52 ± 0,33
mg de protéines mitochondriales
extraites par gramme de muscle dans
les groupes témoins et A23187 respec-
tivement). Ce dernier paramètre n'est
cependant pas le meilleur index de la
densité mitochondriale puisque moins
de 50 % de la population mitochondria-
le est isolée par les techniques couran-
tes d'extraction.
Plusieurs arguments bibliographiques
et les données issues de cette étude sug-
gèrent que l'augmentation de la concen-
tration mitochondriale en Ca2+ sous
l'effet du A23187 a été plus forte dans
le soleus que dans le gastrocnemius.
INRA Productions Animales, Octobre 2006314 / F. CAPEL et al
Figure 9. Effet de l’injection in vivo du ionophore A23187 sur la production mitochon- variation de la production mitochon-
driale d’H2O2 dans les muscles soleus et gastrocnemius chez des rats âgés. driale d'H2O2 n'a été constatée dans le
soleus. Une augmentation précoce de la
concentration mitochondriale en Ca2+ a
pu initier un processus de transition de
perméabilité membranaire de sorte que
des évènements apoptotiques ou nécro-
tiques ont pu survenir dans ce muscle
(Lemasters et al 1998), éliminant ainsi
les mitochondries endommagées.
Dans le gastrocnemius, les mito-
chondries sont moins riches en Ca2+ et
sont théoriquement moins sollicitées
en cas d'une élévation du Ca2+ cyto-
plasmique grâce à l'efficacité du RS.
L'élévation de la concentration du
cation dans la matrice n'a vraisembla-
blement pas suffi pour déclencher
l'apoptose, mais a pu induire l'auto-
phagie des mitochondries les plus sus-
ceptibles au stress calcique (Lemasters
et al 1998), ce qui tend à être confirmé
par la baisse d'activité de la citrate
synthase dans ce muscle. Néanmoins,
Gastrocnemius Soleus elle a induit une stimulation de la pro-
duction d'H2O2 pour toutes les mito-
chondries présentes. Le traitement a
également provoqué une diminution
de l'activité GPX, directement impli-
quée dans le contrôle des concentra-
tions en H2O2 (Antunes et al 2002).
Les mécanismes expliquant cet effet
restent à déterminer. Par ailleurs, l'ef-
fet de la surcharge calcique sur le
stress oxydant est spécifiquement
situé au niveau mitochondrial, puisque
les activités antioxydantes non mito-
chondriales (dans le muscle) n'étaient
pas modifiées par le traitement.
La première conclusion que l'on
peut tirer de ce travail est que les rats
âgés sont plus vulnérables au stress
calcique que les rats adultes pour les-
quels l'injection de A23187 n'a eu
aucun effet. Nos résultats démontrent
également que l'augmentation des
concentrations calciques mitochon-
driales a la capacité de favoriser la
production d'H2O2 par la chaîne respi-
ratoire et qu'il y a une relation inverse
entre la masse du muscle et la teneur
Gastrocnemius Soleus mitochondriale en calcium. Ces résul-
tats permettent de penser que des alté-
* : Significativement différent du groupe témoin (P < 0,05). rations de l'homéostasie calcique
# Significativement différent du groupe correspondant sans antimycine A (P < 0,05). pourraient jouer un rôle dans le déve-
loppement de la sarcopénie. Pourtant,
alors que nos résultats suggèrent que
Nous montrons que les mitochondries beaucoup plus faible (Narayanan et al les muscles oxydatifs sont plus sensi-
du soleus sont plus riches en Ca2+ que 1996). Ce phénomène pourrait être bles à un stress calcique que les mus-
celles du gastrocnemius. En cas de sur- amplifié lors du vieillissement cles glycolytiques, on sait que les
charge calcique, la captation mitochon- (Carmeli et al 2002). Une altération de muscles oxydatifs sont moins affectés
driale de Ca2+ pourrait être plus impor- l'homéostasie calcique pourrait donc par la sarcopénie. D'autres études
tante dans les muscles oxydatifs que induire des perturbations plus impor- seront nécessaires pour mieux com-
dans les muscles glycolytiques tantes au niveau mitochondrial dans les prendre ces relations complexes.
(Sembrowich et al 1985) à cause d'une muscles oxydatifs que dans les muscles
activité de transport du Ca2+ vers le RS glycolytiques. Cependant, aucune
INRA Productions Animales, Octobre 2006Rôle des mitochondries dans le développement d’un stress oxydant dans le muscle squelettique... / 315
7 / Synthèse et perspectives quences réelles de ce stress oxydant susciter devront être mises en oeuvre.
d'origine mitochondriale sur le vieillis- Plusieurs facteurs physiologiques ont
sement musculaire. déjà été proposés, notamment la restric-
L'objectif de ce travail a été de déter- tion énergétique, peut être en liaison
L'hypothèse selon laquelle le niveau avec le statut insulinique (Lambert et
miner si une évolution de la production de production mitochondriale d'H2O2
mitochondriale d'EAO au cours du Merry 2004), l'exercice, en partie en
peut jouer un rôle dans la fonte muscu- rapport avec le métabolisme calcique,
vieillissement pouvait provoquer un laire semble néanmoins confirmée. En
stress oxydant dans le muscle. En effet, l'état inflammatoire et le grand âge. Or
effet, la production mitochondriale ces facteurs sont connus pour interférer
l'analyse de la littérature montre que le d’H2O2 et les niveaux d’oxydation sont
stress oxydant pourrait être l’un des avec les phénomènes du vieillissement
plus élevés dès l’âge adulte dans les musculaire.
éléments responsables de la sarcopénie.
Il est important d'en comprendre les muscles glycolytiques que dans les
muscles oxydatifs. Or ils sont plus De nouvelles questions sont aussi
mécanismes, car préserver la masse apparues, qui concernent les mécanis-
musculaire, c'est préserver la capacité affectés par la sarcopénie.
mes mitochondriaux impliqués, d'au-
de restauration d'un état de bonne santé Le mécanisme exact de ce phénomè- tres origines possibles pour le stress
des personnes âgées et favoriser leur ne reste cependant inconnu. En effet, oxydant et ses conséquences métabo-
autonomie et leur qualité de vie. alors qu'une aggravation du stress oxy- liques.
Ainsi, nous avons évalué les méca- dant d'origine mitochondriale devrait
en premier lieu affecter les mitochon- Ainsi, parmi les mécanismes de cette
nismes et l'évolution avec l'âge de la production mitochondriale d'EAO et de
production d'H2O2 par les mitochon- dries, il est difficile de mettre en évi-
dence une évolution de ces altérations sa régulation, le rôle du flux inverse
dries isolées de muscles oxydatifs ou d'électrons dans l'augmentation de la
glycolytiques. Lorsque cela a été possi- en fonction de l'âge. Ainsi, ni le degré
d'oxydation des protéines mitochon- production mitochondriale d'EAO en
ble, nous avons étudié en parallèle fonction de l'âge devra être précisé,
l'activité des principales enzymes anti- driales, ni l'activité des complexes de la
chaîne respiratoire n'ont varié, que ce notamment dans les muscles oxydatifs,
oxydantes. En effet, le stress oxydant chez d'autres espèces et dans diverses
est dépendant des taux de production soit chez l'homme ou chez le rat.
situations physiologiques. En effet, il a
d'EAO et de l'activité des systèmes été très rarement étudié et nos observa-
antioxydants qui les neutralisent. Cette absence de parallélisme dans
l'évolution de ces paramètres et de la tions mettent en évidence son implica-
Chez le rat, dans un muscle glycoly- production mitochondriale d'EAO en tion dans les phénomènes associés au
tique, la production mitochondriale fonction de l'âge suggère que l'augmen- vieillissement.
d'H2O2 évaluée avec des substrats des tation de cette dernière n'avait qu'un
complexes I ou II n'évoluait pas entre impact limité au moment des observa- Ce travail était focalisé sur la produc-
4,5 et 24 mois. Dans un muscle oxyda- tions. Elle n'aurait donc pas d'effet tion d'EAO par les mitochondries, mais
tif, cette production, évaluée avec des majeur à court terme sur le vieillisse- on sait que d'autres sources d'EAO peu-
substrats du complexe I était plus éle- ment musculaire. En revanche, elle vent aussi être à l'origine d'un stress
vée chez des rats âgés que chez des rats pourrait contribuer à fragiliser le tissu oxydant dans la fibre musculaire. Un
jeunes. Chez l'homme, dans le muscle et se manifester plus particulièrement travail exploratoire original a d'ailleurs
vastus lateralis (métabolisme glycoly- lors d'un stress, ou dans le grand âge suggéré que la contribution de ces au-
tique), la production mitochondriale (au delà de 24 mois chez le rat ou de tres sources à l'augmentation de la pro-
d'H2O2 déterminée en présence d'un 75 ans chez l'homme) lorsqu'elle a pu duction musculaire d'EAO au cours du
mélange de substrats des complexes I et avoir une action prolongée. vieillissement, pourrait être importante.
II augmentait avec l'âge. Nous avons Ce point devra être éclairci et s'il est
montré que cette production était liée Cette conclusion est cohérente avec confirmé, il conviendra de poursuivre
au flux inverse d'électrons entre les les résultats obtenus lorsque les mus- l'analyse de l'activité de ces diverses
complexes II et I. cles de rat ont été soumis à un stress sources et, pour chacune, de leur régu-
calcique par l'injection du ionophore lation.
Ainsi, en résumé, on peut penser que A23187 car les effets qui pourraient
dans les muscles glycolytiques, la pro- résulter d'un stress oxydant d'origine Enfin, les effets sur le métabolisme
duction mitochondriale d'H2O2 induite mitochondriale ne se sont manifestés protéique musculaire des variations des
par le flux normal d'électrons le long de que chez les rats âgés (accroissement différentes formes de production
la chaîne respiratoire reste stable avec de la production mitochondriale d'EAO dans le muscle devront être pré-
l'âge. En revanche, le flux inverse du d'H2O2, de l'activité de la Mn SOD cisés car il est possible que leurs cibles
complexe II vers le complexe I semble dans le muscle gastrocnemius, atrophie et/ou leur impact soient plus ou moins
être impliqué dans un accroissement de du soleus). spécifiques, et qu'elles soient à l'ori-
cette production. La réduction de l'acti- gine :
vité de la GPX observée pourrait aggra- De l'ensemble des résultats obtenus – soit d'adaptations métaboliques
ver ce phénomène. Dans les muscles lors de ce travail émerge donc une diverses puisqu'un rôle signal a été
oxydatifs, la production mitochondriale réponse claire à la question initiale sur attribué à certaines EAO ou composés
d'H2O2 induite par le flux normal le rôle potentiel de la production mito- résultant de leur action.
d'électrons augmente avec l'âge (nous chondriale d'EAO dans le développe- – soit de perturbations à contrôler
n’avons pas testé le flux inverse dans ce ment de la sarcopénie associée au pour en limiter les effets délétères.
type de muscles). Ainsi, on peut vieillissement. De nouvelles approches
conclure que le taux de production physiologiques qui viseront à prolonger Une meilleure connaissance de ces
d'H2O2 par les mitochondries augmen- l'analyse des relations entre les varia- phénomènes devrait permettre de choi-
te avec l'âge dans tous les types muscu- tions de cette production et les pertur- sir les moyens (renforcement des
laires. Il reste à déterminer les consé- bations métaboliques qu'elles peuvent défenses antioxydantes, réduction plus
INRA Productions Animales, Octobre 2006Vous pouvez aussi lire

















































