Effet de la vitesse de filage sur la structure moléculaire et étude de la dynamique de différents acides aminés de soie d'araignée native et ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Effet de la vitesse de filage sur la structure moléculaire et étude de la dynamique de différents acides aminés de soie d’araignée native et supercontractée Mémoire Jane Gagné Maîtrise en chimie - avec mémoire Maître ès sciences (M. Sc.) Québec, Canada © Jane Gagné, 2020
Effet de la vitesse de filage sur la structure moléculaire et étude de la dynamique de différents acides aminés de soie d’araignée native et supercontractée Mémoire Jane Gagné Sous la direction de : Michèle Auger Normand Voyer
Résumé La soie d’araignée, notamment la soie produite par la glande ampullacée majeure (Am), est reconnue comme ayant une ténacité élevée, une bonne résistance mécanique tout en demeurant extensible. Sa structure moléculaire et ses propriétés sont influencées par des facteurs externes comme l’humidité relative et la vitesse de filage. La fibre, par exemple, se contracte de près de la moitié de sa longueur lorsqu’elle est exposée à une humidité élevée, un phénomène nommé supercontraction. Cette étude vise à mieux connaître les propriétés de la soie naturelle et à développer éventuellement une fibre synthétique équivalente. Nous avons caractérisé les fibres de soie Am par spectromicroscopie Raman en lumière polarisée et par résonance magnétique nucléaire (RMN) à l’état solide. La première technique permet de quantifier l’orientation des protéines de soie et de caractériser leur structure secondaire. La RMN permet de déterminer la structure secondaire et la dynamique de relaxation des acides aminés. Les fibres Am de deux espèces d’araignées, Nephila clavipes et Araneus diadematus, ont été obtenues par filage forcé à différentes vitesses (de 0,3 à 2,0 cm/s) et soumises à des taux d’humidité relative supérieurs à 90%. Deux groupes d’araignées ont été marqués isotopiquement avec des solutions de 1-13C-Gly et 1-13C-Ala afin d’analyser la structure et la dynamique d’acides aminés particulièrement importants. Les résultats montrent que l’amplitude de la supercontraction augmente avec la vitesse de filage. Les données indiquent qu’à 1,0 cm/s, l’orientation moléculaire est maximale et qu’elle diminue au-dessus et au-dessous de cette vitesse. Ceci suggère qu’il existe une vitesse de filage pour laquelle les propriétés sont optimales. Les temps de relaxation T1 et T1r des échantillons marqués concordent avec les résultats de spectromicroscopie Raman et suggèrent que les microcristaux de feuillets b de la soie subissent une contraction qui renforce les liaisons intermoléculaires en présence d’eau. ii

Abstract Spider silk, especially the silk produced by the major ampullate glands (Am), is known to have high tenacity, good resistance, and yet remains extensible. Its molecular structure and properties are influenced by external factors such as relative humidity and spinning speed. Fibers, for example, shrink by almost half their length when exposed to high humidity, a phenomenon called supercontraction. This study is intended to better understand the properties of natural spider silk and therefore aid the development of an equivalent synthetic fiber. We characterized Am silk fibers by polarized-light Raman spectromicroscopy and solid-state nuclear magnetic resonance (NMR). The first technique allows for quantification of the orientation of the protein chains and characterization of their secondary structure. Solid-state NMR is used to determine the secondary structure and dynamics of amino acids. The silk of two spider species, Nephila clavipes and Araneus diadematus, was obtained by forced spinning at different speeds (from 0.3 to 2.0 cm/s) and subjected to relative humidity levels greater than 90%. Two groups of spiders were isotopically labeled with 1-13C-Gly and 1-13C-Ala solutions in order to analyze the structure and dynamics of particularly important amino acids. The supercontraction results show that the amplitude of the supercontraction increases with the reeling speed. The data show that at a reeling speed of 1.0 cm/s, the molecular orientation is maximum and decreases above and below this value. This suggests that there is a spinning speed where the properties are optimal. The measurements of the relaxation times T1 and T1r of the labelled samples are consistent with Raman spectromicroscopy results and suggest that b-sheet microcrystals of silk undergo a contraction which strengthens the intermolecular bonds in the presence of water. iii
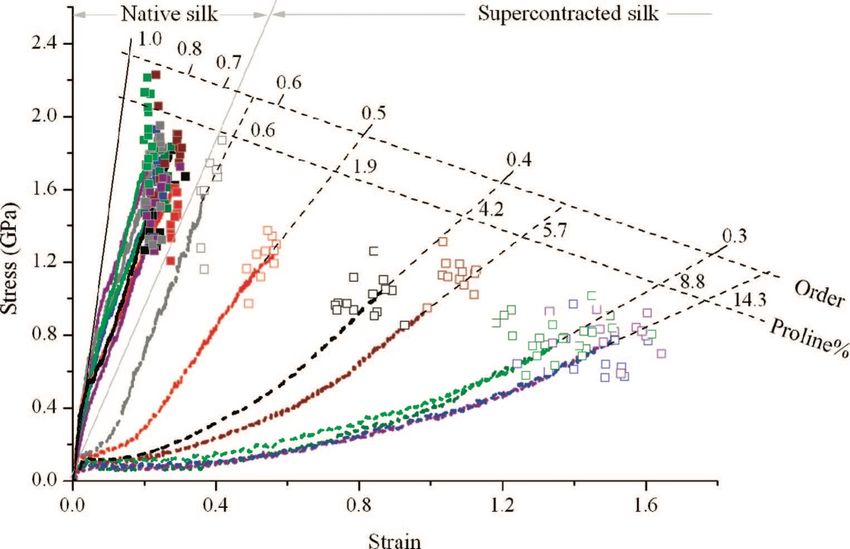
Table des matières Résumé................................................................................................................................ ii Abstract .............................................................................................................................. iii Table des matières ............................................................................................................ iv Liste des figures ................................................................................................................ vi Liste des tableaux ........................................................................................................... viii Liste des abréviations ....................................................................................................... ix Liste des symboles ............................................................................................................ x Remerciements ................................................................................................................. xii Introduction......................................................................................................................... 1 Propriétés de la soie ......................................................................................................... 1 Structure de la soie ........................................................................................................... 2 Supercontraction ............................................................................................................... 4 Processus de filage naturel des araignées ....................................................................... 5 Objectifs du projet ............................................................................................................. 7 Chapitre 1 Techniques utilisées........................................................................................ 9 1.1 Spectromicroscopie Raman ........................................................................................ 9 1.1.1 Effet Raman ......................................................................................................... 9 1.1.2 Spectromicroscopie Raman ............................................................................... 11 1.1.3 Déterminant moléculaire .................................................................................... 13 1.1.4 Fonction de distribution d’orientation.................................................................. 14 1.2 Résonance magnétique nucléaire des solides ......................................................... 16 1.2.1 Principes de base : Nucléaire............................................................................. 17 1.2.2 Principes de base : Magnétique ......................................................................... 18 1.2.3 Principes de base : Résonance.......................................................................... 20 1.2.4 Sensibilité ........................................................................................................... 20 1.2.5 Résonance magnétique nucléaire à l’état solide ................................................ 20 1.2.6 Transformée de Fourier et étude de la dynamique ............................................ 21 Chapitre 2 Matériels et méthodes ................................................................................... 27 2.1 Soie de Nephila clavipes (NC) et d’Araneus diadematus (AD) ................................. 27 2.2 Préparation des échantillons..................................................................................... 28 2.2.1 Échantillon pour la spectromicroscopie Raman ................................................. 28 2.2.2 Échantillon pour la RMN à l’état solide............................................................... 29 2.2.3 Hydrolyse de la soie d’araignée ......................................................................... 30 iv

2.3 Spectromètre Raman ................................................................................................ 31 2.4 RMN à l’état solide .................................................................................................... 32 2.4.1 Mesure de spectres en polarisation croisée à l’angle magique (CPMAS) ......... 32 2.4.2 Mesure des temps de relaxation ........................................................................ 32 Chapitre 3 Effet de la vitesse de filage sur l’orientation moléculaire .......................... 33 3.1 Effet de la vitesse de filage ....................................................................................... 33 3.2 Paramètre pour caractériser l’orientation moléculaire .............................................. 33 3.3 Résultats ................................................................................................................... 34 3.3.1 Effet de la vitesse de filage sur la supercontraction ........................................... 34 3.3.2 Analyse des spectres Raman à différentes vitesses de filage ........................... 35 3.3.3 Comparaison des spectres Raman de la soie native et supercontractée .......... 39 Chapitre 4 Étude de la dynamique des chaînes latérales de la soie ........................... 42 4.1 Choix des acides aminés à l’étude ........................................................................... 42 4.2 Résultats ................................................................................................................... 43 4.2.1 Mesure en CPMAS de la soie native marquée isotopiquement ......................... 43 4.2.2 Temps de corrélation et variations des temps de relaxation .............................. 43 4.2.3 Relaxation et dynamique moléculaire ................................................................ 45 Conclusion ........................................................................................................................ 49 Conclusions .................................................................................................................... 49 Perspectives ................................................................................................................... 49 Bibliographie..................................................................................................................... 51 v

Liste des figures Figure 1 Graphique schématisé de la contrainte en fonction de la déformation de quelques matériaux naturels et synthétiques (Figure modifiée à partir de Gosline et coll2.). ....... 1 Figure 2 Effet de la vitesse de filage sur les propriétés mécaniques de la soie de l’araignée Nephila pilipes de la glande ampullacée majeure (Am)3. .............................................. 2 Figure 3 Schéma d’une fibre de soie d'araignée native où les régions cristallines sont en rouge et les régions amorphes en vert. ......................................................................... 3 Figure 4 Courbes de contrainte-déformation représentatives des soies de la glande ampullacée majeure d’une gamme d’espèces d’araignées testées à l’état natif (trait plein) et à l’état supercontracté (trait pointillé). Le pourcentage de résidus proline dans la séquence des protéines est indiqué. Chaque espèce est identifiée par une couleur distincte, comme suit: gris, Cyrtophora citricola; rouge, Latrodectus hesperus; noir, Nephila edulis; brun, Nephila senegalensis; vert, Nuctenea sclopetaria; mauve, Argiope lobata; vert pâle, Argiope argentata; bleu, Araneus diadematus31. ................. 5 Figure 5 Illustration de la glande ampullacée majeure et ses diverses sections pour l’araignée Nephila clavipes37. ........................................................................................ 6 Figure 6 A.A à l'étude, à gauche l'Gly et à droite Ala ........................................................... 8 Figure 7 Diagramme d’énergie des transitions impliquées durant la diffusion Rayleigh et Raman (Stokes et anti-Stokes), où hu0 est l’énergie incidente, huvib l’énergie vibrationnelle et Du le déplacement Raman (Figure modifiée à partir de Baker et coll56.). ........................................................................................................................... 9 Figure 8 Montage optique du spectromicroscope Raman confocal HR800. ...................... 12 Figure 9 Illustration de la rotation du squelette peptidique selon les angles de rotations dièdres y et f1. ............................................................................................................ 13 Figure 10 Déplacement Raman (cm-1) en fonction des différentes structures secondaires des protéines pour la bande amide I et la bande amide III (Figure adaptée de Tu1.). 14 Figure 11 Systèmes de coordonnées xyz (référentiel de l’unité structurale) et XYZ (référentiel du laboratoire), et angles d’Euler (Figure adaptée de Lefèvre et coll.)62... 15 Figure 12 Orientation d’une fibre de soie d’araignée, d’un feuillet b, d’un groupement carbonyle et du tenseur Raman dans le système d’axes référentiel pour l’analyse en spectroscopie Raman.................................................................................................. 16 Figure 13 Effet de l’application d’un champ magnétique externe sur la dégénérescence des niveaux d’énergie de spin. .................................................................................... 18 Figure 14 Schématisation d’un noyau de spin non nul, du moment cinétique de la masse et du moment magnétique de la charge. ..................................................................... 18 Figure 15 Particules sans champ magnétique extérieur (à gauche) et avec champ magnétique extérieur (à droite) où la précession de Larmor est représentée............. 19 Figure 16 Application de la transformée de Fourier. .......................................................... 22 Figure 17 Orientation et précession d’un noyau de spin ½ dans un champ magnétique constant B0 qui définit l’axe z....................................................................................... 23 Figure 18 Schéma des vecteurs impliqués dans la mesure A) de T1 B) et de T1r64. .......... 25 vi

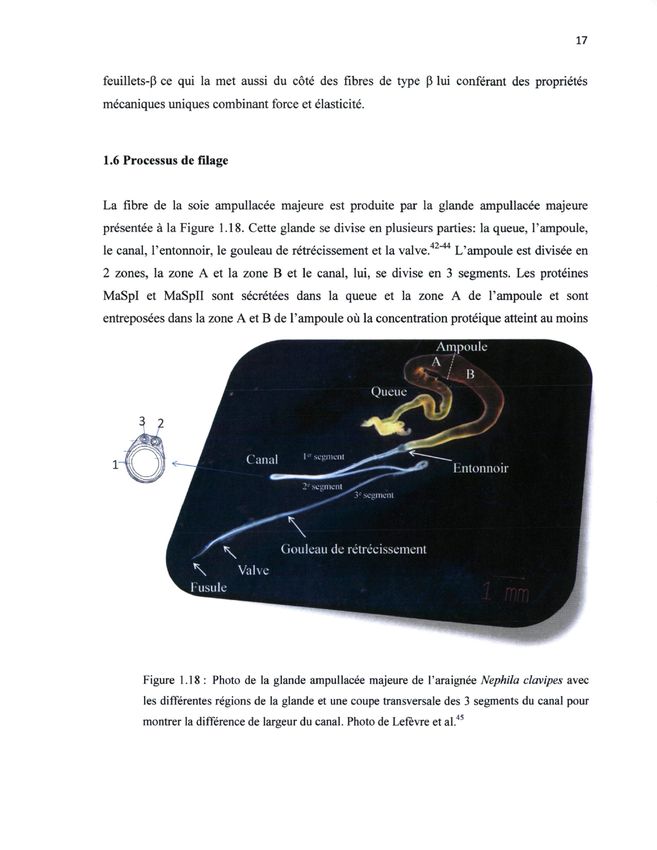
Figure 19 Filière et soie de la glande ampullacée majeure (fil de survie) vue sous binoculaire ................................................................................................................... 27 Figure 20 Montage pour le filage forcé des araignées ....................................................... 27 Figure 21 Préparation d'échantillon pour la spectromicroscopie Raman. En A, nous observons l’éprouvette avec une bobine de soie d’araignée Araneus diadematus où un papier collant permet d’avoir une fibre tendue. En B, nous voyons comment la soie est transférée sur une éprouvette pour les études en spectromicroscopie Raman. En C, nous observons une lamelle sur laquelle une fibre de soie d’araignée est déposée et fixée pour limiter les mouvements. .......................................................................... 28 Figure 22 Tube à hydrolyse utilisé pour la soie d’araignée ................................................ 30 Figure 23 Spectres Raman polarisés typiques en xx et zz d'une fibre de soie ampullacée majeure d’Araneus diadematus................................................................................... 34 Figure 24 Effet de la vitesse de filage sur l’amplitude de la supercontraction d’une fibre de soie d’araignée (AD).................................................................................................... 35 Figure 25 Spectres polarisée xx et zz de la soie de la glande ampullacée majeure native et supercontractée de l'araignée Araneus diadematus obtenue par filage forcé à une vitesse de 1,0 cm/s. Les spectres sont normalisés de sorte que le maximum du pic en Ixx soit égal pour la bande amide I.............................................................................. 36 Figure 26 Effet de la vitesse de filage sur la largeur de la bande amide I et sur sa position pour les fibres de soie ampullacée majeure native et supercontractée de l’araignée Araneus diadematus. Les spectres sont normalisés de sorte que le maximum du pic en Ixx soit égal pour la bande amide I......................................................................... 38 Figure 27 Graphique de la largeur à mi-hauteur de la bande amide I en fonction de la vitesse de filage et de la supercontraction pour la soie ampullacée majeure d’Araneus diadematus. ................................................................................................................. 40 Figure 28 Spectre RMN 13C d’une fibre de soie ampullacée majeure native de l’araignée Nephila clavipes sans marquage isotopique. En bleu, nous pouvons voir la décomposition spectrale.............................................................................................. 42 Figure 29 Dépendances des différents temps de relaxation au temps de corrélation tc (Figure adaptée de Baughman65.). .............................................................................. 44 Figure 30 A, Spectre 13C CPMAS de la soie marquée 1-13C-Ala native en bleu et supercontractée en rouge. B, schéma représentatif de la contraction des feuillets b suite à la supercontraction dans une fibre de soie d’araignée. ................................ 47 vii
Liste des tableaux Tableau 1 Nombres quantiques de l’électron dans le modèle de Hund-Mulliken............... 17 Tableau 2 Paramètres d'orientation R'' en fonction de la vitesse de filage pour la soie de la glande ampullacée majeure native et supercontractée de l’araignée Araneus diadematus déterminés à partir de l’intensité de la bande amide I. ............................ 37 Tableau 3 Effet de la vitesse de filage sur le déplacement Raman et la largeur à mi- hauteur de la bande amide I pour la soie de la glande ampullacée majeure de l'araignée Araneus diadematus. .................................................................................. 39 Tableau 4 Déplacements chimique observés dans la littérature pour diverses structures secondaires pour 1-13C-Ala et 1-13C-Gly ainsi que les déplacements chimiques expérimentaux de la présente étude pour la soie ampullacée majeure de l’araignée Nephila clavipes78, 79. ................................................................................................... 43 Tableau 5 Temps de relaxation 13C T1r (Gly) en fonction de la température pour la soie de la glande ampullacée majeure de l’araignée Nephila clavipes à l’état natif. ............... 45 Tableau 6 Temps de relaxation 1H T1 pour la soie native et supercontractée des deux acides aminés marqués dans la soie ampullacée majeure de Nephila clavipes......... 46 Tableau 7 Temps de relaxation 13C T1r pour la soie native et supercontractée des deux acides aminés marqués dans la soie ampullacée majeure de Nephila Clavipes. ....... 48 viii
Liste des abréviations 2D Deux dimensions A ou Ala Alanine AA Acide aminé AD Araneus diadematus ADF Fibroïne d’Araneus diadematus Am Ampullacée majeure FID Free Induction Decay G ou Gly Glycine L ou Leu Leucine MaSpI Spidroïne I de la glande ampullacée majeure de Nephila clavipes MaSpII Spidroïne II de la glande ampullacée majeure de Nephila clavipes NC Nephila clavipes P ou Pro Proline PC Polarisation croisée Q ou Gln Glutamine RF Radio fréquence RMN Résonance magnétique nucléaire S ou Ser Sérine SC Supercontractée ou supercontraction TI Fonction d’impulsion d'inversion TF Transformée de Fourier Y ou Tyr Tyrosine ix
Liste des symboles " Tenseur de polarisabilité "# Tenseur de polarisabilité à l’état d’équilibre Bz Champ magnétique externe appliqué B1 Champ magnétique appliqué B0 Champ magnétique externe Beff Champ magnétique effectif Bd Champ magnétique dipolaire σ Contrainte à la rupture E Module de Young %⃗ Champ électrique incident du laser %⃗# Champ électrique incident à l’état d’équilibre Déformation à la rupture ℎ Constante de Planck J Moment cinétique k Constante de Boltzmann l Nombre quantique azimutal M0 Magnétisation provenant de l’échantillon Mx Vecteur du moment magnétique dans l’axe x My Vecteur du moment magnétique dans l’axe y Mz Vecteur du moment magnétique dans l’axe z m Masse atomique ml Nombre quantique magnétique ms Nombre quantique magnétique de spins n Nombre quantique principale N(q, f, y) Fonction de distribution d’orientation (dépendant des angles d’Euler q, f et y) %⃗ Dipôle induit q Charge électrique atomique %⃗ Coordonnées du mode normal de vibration %⃗# Coordonnées à l’équilibre du mode normal de vibration t Durée d’application du champ B1 tc Temps de corrélation Moment magnétique de spin n Fréquence (Hz) Pulsation (rad/s) Rapport gyromagnétique / Angle d’inclinaison f Angle azimutal Y Angle de la rotation de précession xyz Référentiel de l’unité structurale d’une molécule XYZ Référentiel du laboratoire x
Je dédie ce mémoire à mes amis non scientifiques qui n’y comprendront rien lors de leur lecture promise. xi
Remerciements J’aimerais tout d’abord remercier ma directrice de recherche Michèle Auger. J’ai eu la chance d’étudier dans un laboratoire de recherche où nous sommes bien plus que des collègues de travail et où des amitiés se forment. J’ai eu l’honneur de côtoyer une grande scientifique, une femme exceptionnelle, attentionnée et passionnée. Je suis certaine qu’elle veille sur nous tous les jours. « Une grande âme sert tout le monde tout le temps. Une grande âme ne meurt jamais. Elle nous rassemble encore et encore. » -Maya Angelou Un gros merci à Normand Voyer et ses étudiants de nous avoir accueillis dans leur laboratoire à bras ouverts. Je tiens également à remercier notre professionnel de recherche Thierry Lefèvre. Tout ce travail est en partie grâce à toi. Tu as su prendre les rênes du laboratoire et assurer son bon fonctionnement. Tous tes conseils autant sur les araignées que sur la spectroscopie m’ont été extrêmement utiles. J’ai très hâte de continuer de travailler avec toi sur notre livre sur la soie d’araignée. J’ai créé de belles amitiés dans le groupe Auger notamment avec Benji, Gab et Koral. Je vous remercie des beaux moments que vous m’avez fait vivre à vos côtés. Les apéros au bureau vont me manquer ainsi que le fameux Caps. J’ai le regret de vous annoncer qu’il faut maintenant payer notre tricount. J’espère que cette amitié saura survive aux années et à la distance et que Benji nous invitera à NY l’année prochaine (même si je préfèrerais la Cali…). J’aimerais également remercier la grande famille du Rouge et Or et plus particulièrement la famille du Rouge et Or Basketball. Le sport universitaire m’a fait vivre de merveilleux moments et m’a fait grandir en tant que joueuse et en tant qu’adulte. J’ai eu la chance de rencontrer des personnes exceptionnelles et de tisser de forts liens d’amitié. Je tiens finalement à remercier ma famille qui m’a toujours soutenue et encouragée dans le sport ainsi que dans mes études. Je suis choyée d’avoir une si belle et grande famille. Merci Papa, Maman, Stef, Mymi et mon fiancé Marco. xii
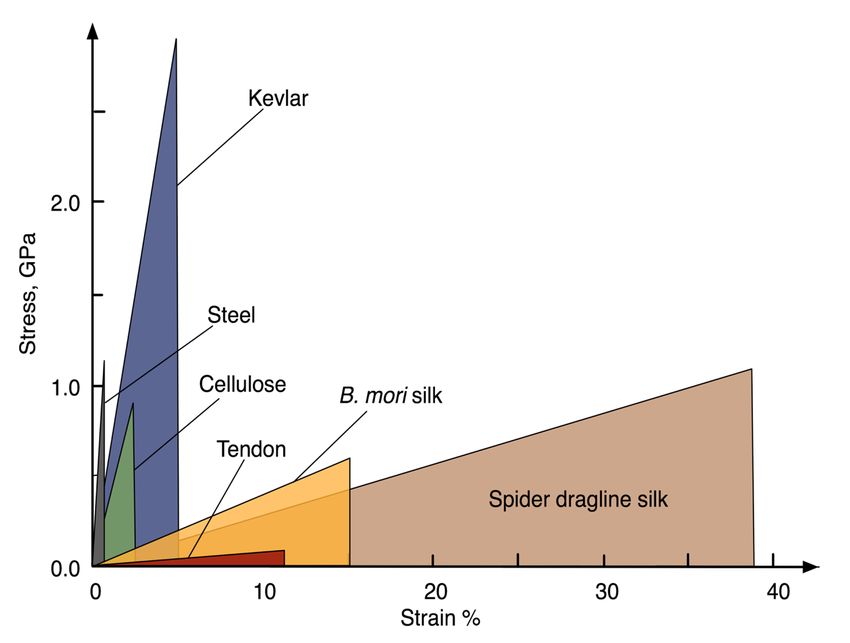
Introduction Propriétés de la soie La soie d’araignée est un ensemble de matériaux exceptionnels produits par la nature. Les propriétés mécaniques de la soie ampullacée majeure (Am), ou fil de survie, sont supérieurs à de nombreux matériaux, autant naturels que synthétiques. La soie d’araignée Am est reconnue comme étant une fibre avec une ténacité élevée, une bonne résistance mécanique tout en demeurant extensible. Kevlar Contrainte GPa Acier Cellulose Soie B. mori Tendon Soie d’araignée (fil de survie) Déformation (%) Figure 1 Graphique schématisé de la contrainte en fonction de la déformation de quelques matériaux naturels et synthétiques (Figure modifiée à partir de Gosline et coll2.). Pour caractériser les propriétés mécaniques d’étirement, on mesure la contrainte en fonction de l’élongation. Quelques matériaux sont représentés schématiquement à la Figure 1. La force nécessaire pour briser un matériau, plus précisément la tension nécessaire, se nomme la contrainte à la rupture (σ). C’est une mesure de force par unité de surface exprimée en N/m2. Ainsi, la force à appliquer pour briser la soie d’araignée est du même ordre que celle nécessaire pour briser l’acier2. La Figure 1 montre également la déformation à la rupture ( ), soit le pourcentage d’élongation par rapport à la longueur initiale. Pour la soie Am, elle est d’environ 37 % tandis que celle de l’acier s’approche de 1 %. L’aire sous la courbe représente l’énergie nécessaire pour briser le matériau, que l’on appelle résistance. Celle de la soie Am surpasse la plupart des matériaux synthétiques, dont le Kevlar, une fibre bien connue dans le domaine du textile pour ses propriétés mécaniques remarquables, et l’acier. 1
clearly shows the difference of ‘‘particle size’’ due to the fibril segments proposed mergence of the silk fibril segments at higher reeling speed. segment consists of seve A model for describing the corresponding nanostructure b-structures. This featur change with increasing reeling speed is illustrated in Fig. 7, dragline silk may play an d–f. An alternative explanation for the decrease of the high mechanical performa !1 intercrystallite distance above 10 mms may be obtained La soie étant produite au cours d’un processus complexe impliquant une traction exercée par l’araignée (voir from the relation between the nucleation rate and reeling section Processus de filage), speed.lesAccording propriétés mécaniques to Eq. 4, thede la soie sont density crystallite influencées par lathe within vitesse de filage The relation between (Figure 2). Plus la vitesse derises silk filagewith est élevée, plus la contrainte the increasing reelingàspeed. la rupture est grande, ce Accordingly, thequi indique une mechanical property intercrystallite distance becomes shorter as the reeling speed fibre plus solide. La déformation à la rupture tend, elle, à diminuer, montrant que la soie devient plus cassante. increases. The mechanical propert Le module de Young, qui correspond à la pente initiale de la courbe tension-élongation, The above results on the size measurement of silk fibril augmente avec la considerably with the reel vitesse de filage, ce quisegments indique queusing AFM la soie (40–80 devient de plusnm), together en plus rigide. with the results Finalement, curves start to deviate fro la limite d’élasticité of the crystallite size (2–7 nm) and intercrystallite distance point. The stress and str augmente également avec la vitesse de filage dénotant une fibre plus dure. ‘‘yield stress’’ and ‘‘yie slope of the stress-strain yield point is Young’s m be calculated from stress shown in Figs. 8 b and should be determined at t structural factors, such as Contrainte GPa Contrainte à la rupture Déformation (%) Figure 2 Effet de la vitesse de filage sur les propriétés mécaniques de la soie de l’araignée Nephila pilipes de la glande ampullacée majeure (Am)3. La supercontraction (SC) est un phénomène qui influence aussi les propriétés mécaniques de la soie, entre autres, en diminuant la rigidité et en augmentant l’extensibilité2, 4, 5. Plus amples détails seront donnés sur cette contraction provoquée par l’eau. Structure de la soie Toutes les araignées sont capables de produire et d’utiliser une variété de soies différentes. Cependant, les araignées orbitèles, qui produisent des toiles circulaires planes, sont celles qui en utilisent le plus. Ces araignées peuvent produire jusqu’à sept types de soies, toutes ayant des fonctions biologiques diverses. Chaque soie provient d’une glande précise située dans les filières, qui se trouvent sur l’abdomen de FIGURE 8 Comparison of silks drawn at different speeds from spider l’araignée2, 6. La soie estN. constituée pilipes. (a) de protéines curve. Stress-strain fibreuses The nommées curves startspidroïnes. La structure to deviate from the first primaire des linear part at the yield point. The stress and strain at the yield point are called spidroïnes, c’est-à-dire leur séquence d’acide aminé (AA), contrôle en partie l’arrangement moléculaire dans la ‘‘yield stress’’ and ‘‘yield strain’’. respectively. (b) The slope of the stress- fibre. Le processus de filage strainest l’autre curve in afacteur versusintervenant strain at lowdans la showing strain structurethe tridimensionnelle de la soie. yield point. (Inset) The calculated Young’s modulus (slope in the linear region of the stress- FIGURE 9 Effect of the reel strain curve) with reeling speed. spider dragline silk. (a) The effe 2
Le poids moléculaire des spidroïnes Am est d’environ 350 kDa7. La structure secondaire des protéines influence les propriétés mécaniques et biologiques de la soie. La soie Am est reconnue comme étant un biomatériau des plus résistants. L’araignée utilise ce type de soie pour le cadre et les rayons de sa toile, en plus de fil de survie pour échapper aux prédateurs. Il s’agit ainsi d’un matériau ayant une très bonne résistance à la traction et une excellente extensibilité8. La glande Am exprime en fait deux spidroïnes, MaSpI (Major ampullate spidroin I) et MaSpII (Major ampullate spidroin II) 9, qui constituent respectivement 81 % et 19 % de la soie de Nephila clavipes (NC)10. Le ratio entre MaSpI et MaSpII varie selon l’espèce d’araignée, ce qui influence les propriétés de la soie10, 11. Les deux spidroïnes possèdent de fortes ressemblances. Les régions situées aux extrémités, soit les séquences non répétitives N-terminale et C-terminale, sont hautement conservées pour les différents types de soie et les espèces d’araignées7, 12, 13. De plus, les deux protéines comportent un long domaine central répétitif de type copolymère bloc qui alterne des régions riches en glycine (Gly) et des régions riches en alanine (Ala)14. La spidroïne 1 (MaSpI) contient des motifs polyalanine (poly-A), GA et GGX où X peut-être Y (Tyr), L (leu) ou Q (Gln)12. Les séquences de poly-A peuvent enchainer de 3 à 7 résidus Ala et se trouvent à chaque bout de chaine12. Ainsi l’empilement des chaînes protéiques est favorable et il y a formation de nanocristaux de feuillets β. Ces feuillets b sont alignés dans l’axe de la fibre. Ils sont responsables de la rigidité de la fibre9, 15, la quantité de feuillets β dictant la cristallinité de la soie. Les motifs GGX ne permettent pas un repliement aussi efficace que pour les séquences poly-A16. Ainsi, on obtient une phase amorphe désordonnée, qui constitue la majeure partie de la soie, et qui disperse la phase cristalline (Figure 3)9. Les motifs GGX se retrouvent sous forme d’hélices 31, de coudes et de segments désordonnés dans la matrice amorphe. La plus grande différence entre MaSpI et MaSpII est la quantité de résidus proline (Pro). Pour NC, MaSpI ne contient pas de résidus Pro tandis que MaSpII en contient 15 %. On retrouve généralement la Pro dans les motifs GPGXX17. Celui-ci se replie sous forme de coudes β contribuant au domaine semi-amorphe et désordonné et procure de l’extensibilité et de la mobilité aux chaines polypeptidiques18, 19. Une importante corrélation existe entre l’abondance de résidus Pro dans la spidroïne MaSpII, l’extensibilité de la soie Am et l’amplitude de la SC5, 9 (voir section suivante). Figure 3 Schéma d’une fibre de soie d'araignée native où les régions cristallines sont en rouge et les régions amorphes en vert. 3
Guerette et coll.20 ont répertorié quatre fibroïnes (ADF-1, ADF-2, ADF-3, ADF-4) différentes pour l’espèce Araneus diadematus (AD). Dans le cas de la glande Am, les protéines qui la composent sont l’ADF-3 et l’ADF- 4 selon un ratio de 3:2. Contrairement à NC, ces deux spidroïnes sont riches en Pro. L’ADF-3 contient un bloc cristallin riche en Ala (ASAAAAAA) et plusieurs blocs riches en Gly et en Pro (GPGGQGPYGPG, GGYGPGS et GPGQQ) pour la phase amorphe. En ce qui concerne l’ADF-4, le bloc donnant lieu à un milieu cristallin est SSAAAAAAAA et ceux donnant lieu à des domaines amorphes sont GPGSQGPS et GPGGY20. En général, les deux spidroïnes ont une composition en Ala, Pro et Gly similaire tandis que leur composition en serine (Ser) et en Gln varient, ce qui influence des propriétés comme l’hydrophobicité et la solubilité21. Dans la plupart des cas, la composition en feuillets β constitue 29 % de la structure secondaire du fil de survie d’AD. On retrouve des coudes β à 12-15 % et des hélices 31 à 19 %22. Supercontraction Lorsque la soie est soumise à une humidité élevée ou est mouillée par l’eau, la fibre se contracte longitudinalement tandis que son diamètre augmente4. La contraction peut aller jusqu’à 50 % de la longueur initiale de la fibre4. Ce phénomène est connu sous le nom de supercontraction (SC). Son amplitude dépend de l’espèce d’araignée, du type de soie et de la vitesse de filage23, 24. Lors de ce phénomène, les molécules d’eau s’infiltrent dans la microstructure de la soie, perturbant ainsi le réseau de liaisons hydrogène, ce qui entraine une augmentation de la mobilité des chaines polypeptidiques. De ce fait, celles-ci se replient spontanément, amenant l’entropie à son maximum et cause la contraction et le gonflement de la fibre25. Les liaisons hydrogène dans la phase amorphe sont perturbées par l’insertion des molécules d’eau, car celles-ci ont un effet plastifiant, alors que les feuillets β ne semblent pas affectés26-30. Il a été démontré que l’amplitude de la SC est influencée par la composition chimique de la soie. En effet, la quantité de résidus Pro dans la structure affecte la contraction de la soie (Figure 4)23, 31, 32. La Pro se distingue des autres AA par sa chaîne latérale qui forme un cycle remontant sur son squelette. Cette structure affecte les angles de torsions et empêche l’adoption d’une configuration stable comme les feuillets b ou les hélices a. 4
Soie Soie SC Soie native SC Contrainte (Gpa) Déformation (%) Figure 4 Courbes de contrainte-déformation représentatives des soies de la glande ampullacée majeure d’une gamme d’espèces d’araignées testées à l’état natif (trait plein) et à l’état supercontracté (trait pointillé). Le pourcentage de résidus proline dans la séquence des protéines est indiqué. Chaque espèce est identifiée par une couleur distincte, comme suit: gris, Cyrtophora citricola; rouge, Latrodectus hesperus; noir, Nephila edulis; brun, Nephila senegalensis; vert, Nuctenea sclopetaria; mauve, Argiope lobata; vert pâle, Argiope argentata; bleu, Araneus diadematus31. De plus, la Pro n’a pas de groupes amide pour établir des liaisons hydrogène et stabiliser les structures dans lesquelles elle est impliquée. Pour NC, chaque Pro est associée à quatre autres groupes peptidiques dans la séquence GPGXX, de sorte que ces quatre groupes sont intrinsèquement « désordonnés » par la Pro33, 34. Par conséquent, cet AA désavantage la formation de structures en hélice α et feuillet β, mais tend à induire des coudes β et γ dans le squelette. Le motif GPGXX perturbe ainsi la structure cristalline de la soie d’araignée34 et augmente l’élasticité de la fibre35. Cependant, certaines espèces d’araignées qui ne possèdent pas de Pro ou en ont très peu subissent la SC23. Par conséquent, celle-ci ne s’explique pas uniquement par la présence de Pro23. Goseline et al ont proposé une hypothèse selon laquelle la SC pouvait également être provoquée par la présence d’AA polaires et chargés36. Processus de filage naturel des araignées Comme pour toutes les soies, la formation de la fibre Am a lieu dans une glande spécifique. La Figure 5 montre cette glande et ses différentes subdivisions anatomiques. Initialement, la soie est un liquide très 5
mécaniques uniques combinant force et élasticité. 1.6 Processus de filage La fibre de la soie ampullacée majeure est produite par la glande ampullacée majeure visqueux. C’est seulement après avoir traversé la glande Am que la soie se retrouve sous forme de fibre présentée à la Figure 1.18. Cette glande se divise en plusieurs parties: la queue, l'ampoule, insoluble. Cette transformation est générée par la traction mécanique que l’araignée exerce lorsqu’elle file, 42 44 le canal, l'entonnoir, le gouleau de rétrécissement et la valve. " L'ampoule est divisée en que ce soit2 lors de lala zone zones, construction de saB toile A et la zone et leou la production canal, d’unenfil3 de lui, se divise survie lorsqu’elle segments. se déplace ou Les protéines qu’elle chute. MaSpI et MaSpII sont sécrétées dans la queue et la zone A de l'ampoule et sont entreposées dans la zone A et B de l'ampoule où la concentration protéique atteint au moins Ampoule A / . / B Queue Canal I c v seemenl Entonnoir 2e segment Gouleau de rétrécissement Valve Fusule Figure 1.18 : Photo de la glande ampullacée majeure de l'araignée Nephila clavipes avec Figure 5 Illustration de la glande ampullacée majeure et ses diverses sections pour l’araignée Nephila les différentes régions de la glande et une coupe transversale des 3 segments du canal pour clavipes37. montrer la différence de largeur du canal. Photo de Lefèvre et al.45 À la Figure 5, on peut voir plusieurs parties de la glande Am, dont la queue, l’ampoule, le goulot de rétrécissement (ou entonnoir), le canal et la valve11. L’ampoule est séparée en deux sections, la section A et la section B tandis que le canal se divise en trois segments. Le filage commence au moment où les cellules épithéliales de la queue de l’ampoule (zone A) sécrètent les protéines MaSpI et MaSpII. Au départ, les protéines sont solubles dans l’eau et constituées de structures désordonnées et partiellement hélicoïdales empêchant ainsi la soie de se replier prématurément38. La solubilisation des protéines de soie à l’état liquide est critique, car elle doit à la fois empêcher la précipitation prématurée des protéines tout en permettant une formation rapide d’un fil solide insoluble à la fin du conduit39. En solution, les protéines sont sous forme de dimères et sont stabilisées par des ponts disulfures. Les protéines seront transformées en fibre composée de feuillets β et de domaines amorphes au fil de leur parcours dans la glande. Lorsque l’araignée a besoin de soie, les protéines emmagasinées dans l’ampoule se dirigent vers les autres sections de la glande. Les conditions du milieu évoluent au sein de la glande. Le pH varie ainsi de 6,7 dans l’entonnoir à 6,3 quand on se rapproche du canal. Tout au long de ce dernier, le pH 6
passe de 6,3 à 5,7 et la concentration en sel diminue40, 41. Le gradient de pH est maintenu grâce à la présence d’anhydrase carbonique. Des forces de cisaillements deviennent de plus en plus intenses au fur et à mesure que le diamètre du canal diminue, ce qui provoque l’alignement des chaines de protéine parallèlement à l’axe de la fibre27, 42, 43. Le domaine C-Terminal est alors déstabilisé ce qui permet aux séquences répétitives d’adopter une structure riche en feuillets β. À un pH d’environ 6, les groupements N-terminal favorisent la formation de la fibre. Cela peut être lié au développement de dimère stables44. C’est également à ce niveau que le NaCl a le plus grand impact, en effet, le sel influence la structure tertiaire45. Le pH diminue progressivement dans le canal, ce qui provoque la dimérisation du groupement N-terminal et la synthèse d’homodimères (pH
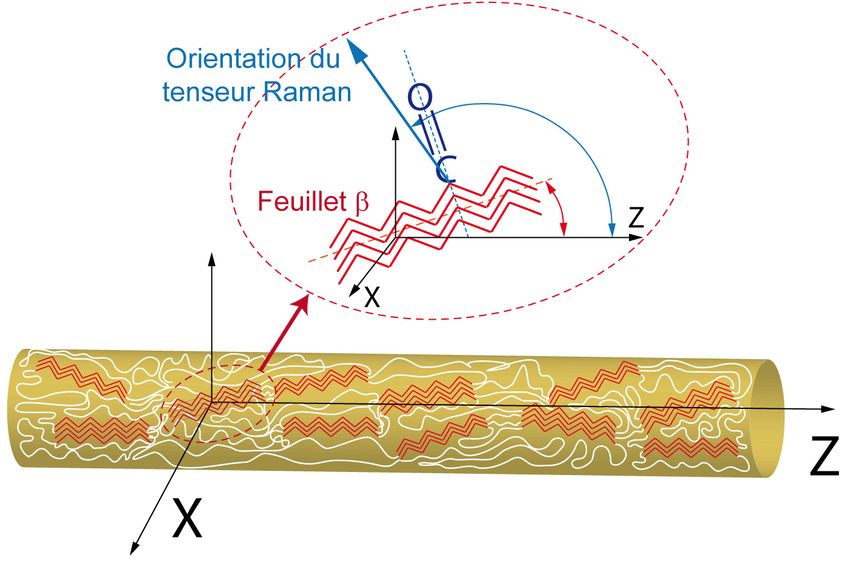
De ce fait, la première partie de ce manuscrit porte sur l’effet de la vitesse de filage et de la SC sur l’orientation moléculaire de fibres de soie d’araignée, une étude réalisée par spectromicroscopie Raman. La seconde partie porte sur l’étude de la dynamique des chaînes polypeptidiques via deux AA marqués isotopiquement, 1- 13C-Ala et 1-13C-Gly (Figure 6). Le choix de ces deux AA repose sur deux facteurs. Premièrement, la Gly et Ala sont deux AA présents en grande quantité dans la soie d’araignée. Deuxièmement, ces deux AA se retrouvent dans deux environnements différents au sein d’une fibre de soie d’araignée. La Gly est située principalement dans les régions amorphes de la soie. Quant à Ala, elle se retrouve majoritairement dans les feuillets b qui sont dans les régions cristallines de la soie. La mesure des temps de relaxation T1 et T1r par RMN à l’état solide sur les protons et les carbones respectivement, nous informent sur la rigidité et la mobilité des AA présents dans deux structures secondaires distinctes au sein de la soie. Glycine Alanine Figure 6 A.A à l'étude, à gauche l'Gly et à droite Ala 8
Chapitre 1 Techniques utilisées 1.1 Spectromicroscopie Raman 1.1.1 Effet Raman La spectroscopie Raman permet l’analyse des différents modes vibrationnels d’une molécule donnée. La diffusion Raman, ou l’effet Raman implique une diffusion inélastique de la lumière visible, impliquant un échange d’énergie entre le photon incident et la molécule analysée. Ce phénomène optique a été découvert en 1928 simultanément par les physiciens Chandrashekhara Raman et Leonid Mandelstam55. La Figure 7 schématise les transitions vibrationnelles en spectroscopie Raman. Figure 7 Diagramme d’énergie des transitions impliquées durant la diffusion Rayleigh et Raman (Stokes et anti-Stokes), où hu0 est l’énergie incidente, huvib l’énergie vibrationnelle et Du le déplacement Raman (Figure modifiée à partir de Baker et coll56.). Lorsqu’un échantillon est irradié par une source de lumière monochromatique dans le visible, une grande partie de la lumière est diffusée à la même longueur d’onde que celle correspondant au photon incident. Il n’y a dès lors pas d’échange d’énergie entre la molécule et le photon. On parle donc de diffusion élastique, aussi connue sous le nom de diffusion Rayleigh. À l’inverse, la diffusion Raman se produit lorsqu’il y a diffusion inélastique. Ceci implique un échange d’énergie entre le photon incident et la molécule étudiée. Un complexe photon-molécule est alors formé dans un niveau d’énergie virtuel. Étant donné que l’énergie d’excitation n’est pas suffisamment élevée pour atteindre un niveau électronique excité, un photon est réémis et retombe dans un niveau d’énergie plus faible ou plus élevé que le niveau fondamental. Ainsi, la lumière diffusée n’a pas la même longueur d’onde que la lumière incidente. Deux scénarios sont possibles lorsqu’on parle de diffusion Raman. Le premier se produit lorsque la lumière est décalée vers de plus grandes longueurs d’onde, donc à plus faible énergie, et se nomme la diffusion Raman Stokes. Le second se produit lorsque la lumière est 9
décalée vers de plus courtes longueurs d’onde, donc à plus forte énergie, et se nomme la diffusion Raman anti-Stokes. La différence d’énergie entre le photon incident et celui diffusé est reliée à la composition chimique de l’échantillon et à la structure moléculaire. De ce fait, il est possible d’analyser la composition chimique de l’échantillon selon sa capacité à diffuser la lumière. Puisque l’intensité du signal en Raman est directement liée au nombre de molécules dans les différents modes vibrationnels, les raies Stokes sont étudiées davantage. En effet, à température ambiante, les modes vibrationnels de basse énergie sont plus peuplés, ainsi les raies Stokes sont plus intenses que les raies anti- Stokes57. La répartition des deux états énergétiques suit la distribution de Boltzmann : ∆5 ./ (1) .0 = 367 , où N2/N1 est le rapport entre la population du premier niveau excité par rapport au niveau fondamental, DE est la différence d’énergie entre chaque niveau du système (J), T est la température en Kelvin (K) et k est la constante de Boltzmann (1,38×10-23 J.K-1). La variation de la polarisabilité électronique est très importante en spectroscopie Raman. Toutes les molécules possèdent une polarisabilité électronique a, qui peut être exprimée par une faible distorsion du nuage électronique. Au moment où l’échantillon est irradié par le laser, le champ électrique ( %⃗ ) incident interagit avec les électrons du nuage électronique de la molécule et induit un dipôle ( %⃗), ce qui se traduit par l’équation suivante : %⃗ = " ∙ %⃗ , (2) où " est un tenseur de rang 2 décrivant la polarisabilité électronique de la liaison étudiée. L’équation du champ électrique incident à un temps donné est représentée ci-dessous : %⃗ = %⃗# cos(2 # ), (3) où %⃗# décrit le champ électrique à l’état d’équilibre et # la fréquence de la radiation incidente. Lorsque l’amplitude de vibration est petite, la polarisabilité est une fonction linéaire du mode normal de %⃗ et peut s’exprimer comme : vibration " (4) " = "# + E G %⃗# + ⋯ , %⃗ # 10
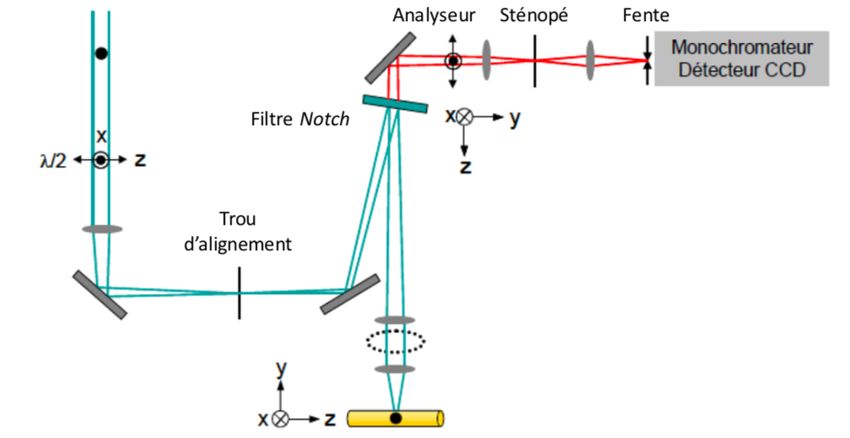
L JK où "# est la polarisabilité de la liaison à la position d’équilibre, IJM%⃗N est le taux de variation de " par rapport # %⃗ et à la variation de %⃗# est le vecteur représentant les coordonnées à l’équilibre du monde normal de %⃗ décrit le déplacement moléculaire et s’écrit : vibration. Si la liaison étudiée vibre à une fréquence , %⃗ = %⃗# cos (2 PQR ) (5) Une vibration sera active en Raman si la vibration entraîne un changement de polarisabilité électronique de la liaison tel que : " (6) E G ≠0 %⃗ # Lorsqu’on remplace, dans l’équation du dipôle induit %⃗, les paramètres de " et %⃗ on obtient : " (7) %⃗ = "# ∙ %⃗# (2 # ) + E G %⃗# ∙ %⃗# cos (2 # ) ∙ cos (2 PQR ) %⃗ # À l’aide de l’identité trigonométrique cos( + ) + cos ( − ) (8) cos cos = , 2 on trouve : ] JKL (9) %⃗ = "# ∙ %⃗# (2 # ) + ^ IJM%⃗ N %⃗# ∙ %⃗# cos(2 ( # − PQR ) ) # 1 " + E G %⃗# ∙ %⃗# cos (2 ( # + PQR ) ) 2 %⃗ # Les trois termes observés dans la dernière équation correspondent respectivement à la diffusion Rayleigh, la diffusion Raman Stokes et la diffusion Raman anti-Stokes. 1.1.2 Spectromicroscopie Raman Le spectromètre Raman utilisé dans cette étude est couplé à un microscope optique ce qui permet de focaliser le faisceau laser sur une petite surface et d’atteindre une résolution latérale d’environ 2-3 µm. Ainsi, cette technique est très efficace pour sonder des échantillons comme la soie d’araignée qui possèdent un diamètre pouvant varier entre quelques centaines de nanomètres et 10 µm58. Cette technique est très utile pour obtenir de l’information sur la conformation et sur l’orientation des protéines grâce à l’utilisation de lumière polarisée 11
Vous pouvez aussi lire

















































