Effets de l'hivernage en banques sur la survie et la qualité reproductive des reines d'abeilles mellifères (Apis mellifera L.) - Mémoire Mireille ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Effets de l'hivernage en banques sur la survie et la
qualité reproductive des reines d'abeilles mellifères
(Apis mellifera L.)
Mémoire
Mireille Levesque
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Mireille Levesque, 2022
Effets de l’hivernage en banques sur la survie et la
qualité reproductive des reines d’abeilles mellifères
(Apis mellifera L.)
Mémoire
Mireille Levesque
Sous la direction de :
Pierre Giovenazzo, directeur de recherche
Résumé
L’industrie apicole occupe une place cruciale au sein de l’économie canadienne et est en
expansion depuis plusieurs années. Les conditions climatiques du pays entraînent
cependant des pertes importantes de colonies et limitent l’élevage de reines tôt au
printemps. Les apiculteurs doivent donc importer des reines d’autres pays afin de remplacer
les colonies perdues, ce qui contribue à la dissémination d’agents pathogènes et de lignées
d’abeilles ayant des traits génétiques indésirables pour l’apiculture canadienne.
L’acquisition de reines locales au printemps est donc souhaitable et pourrait être facilitée
par l’hivernage de reines en banques. En élevant des reines à la fin de l’été et en les
regroupant dans une même colonie appelée banque durant la période hivernale, les
apiculteurs auraient accès à des reines locales tôt au printemps. Cette technique est récente
et les connaissances à ce sujet ont besoin d’être approfondies. Dans ce contexte, les
objectifs de mon projet étaient d’évaluer l’effet de deux densités de reines (40 et 80)
entreposées en banques sur leur survie hivernale et d’évaluer l’influence de l’hivernage des
reines en banques sur leur qualité reproductive et leur performance en colonie. Nos résultats
montrent que la survie hivernale des reines est supérieure à la densité la plus basse.
L’hivernage des reines en banques n’affecte pas leur fertilité, mais réduit leur taille, leur
poids et leur ponte durant leurs premières semaines en colonie au printemps. Néanmoins,
après leurs premières semaines en colonie, les reines hivernées en banques regagnent une
taille et un poids comparables à ceux de reines hivernées libres dans leur colonie respective.
Ce projet démontre le potentiel de l’entreposage des reines en banques pour conserver des
reines durant la période hivernale et ainsi assurer un approvisionnement en reines locales
tôt au printemps. Cette méthode favorise aussi le maintien de traits génétiques adaptés aux
besoins de l’industrie apicole canadienne.
ii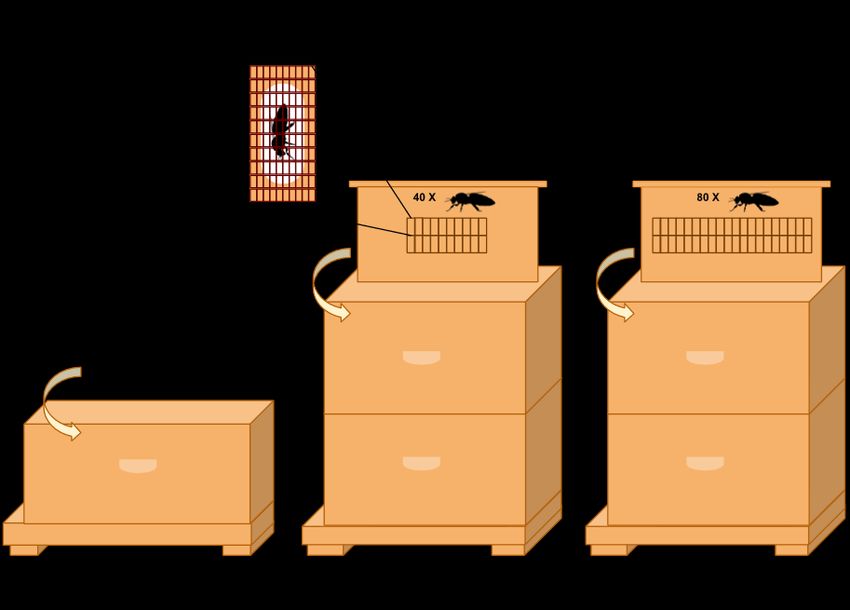
Abstract
The beekeeping industry is a vital part of Canadian economy and has been expanding for
several years. However, winter causes significant colony losses and limits the rearing of
queens in early spring. As a result, beekeepers must import queens from other countries to
replace lost colonies, which contributes to the spread of pathogens and undesirable genetic
traits. Developing the country’s autonomy towards queen imports in early spring is
therefore essential and could be facilitated by overwintering queens in a bank colony. By
rearing queens at the end of summer and grouping them in a colony called a bank during
the winter period, beekeepers would have access to local queens in early spring. This
method is recent and the knowledge on this subject needs to be broadened. In this context,
the objectives of my project were to assess the effect of two densities of queens (40 and 80)
stored in a bank colony on their winter survival and to evaluate the influence of
overwintering queens in a bank colony on their reproductive quality and colony
performance. Our results show that queen survival rate is higher at the lowest density.
Overwintering queens in a bank colony does not compromise their fertility, but reduces
their size, weight and oviposition during their first weeks in a colony in spring.
Nevertheless, after their first weeks in the colony, queens overwintered in bank colonies
regain a size and weight comparable to that of queens overwintered free in their respective
colony. This project demonstrates the potential of storing queens in a bank colony during
the winter period. This method ensures a supply of local queens early in the spring and
maintains genetic traits corresponding to the needs of the Canadian beekeeping industry.
iiiTable des matières
RÉSUMÉ ........................................................................................................................................... II
ABSTRACT ..................................................................................................................................... III
TABLE DES MATIÈRES .............................................................................................................. IV
LISTE DES TABLEAUX ............................................................................................................... VI
LISTE DES FIGURES ................................................................................................................. VII
LISTE DES ABRÉVIATIONS ...................................................................................................... IX
GLOSSAIRE .....................................................................................................................................X
REMERCIEMENTS..................................................................................................................... XII
AVANT-PROPOS ........................................................................................................................ XIV
INTRODUCTION ............................................................................................................................. 1
L’ABEILLE MELLIFÈRE .................................................................................................................................... 1
Historique ................................................................................................................................................. 1
La colonie d’abeilles ................................................................................................................................. 1
LA REINE ........................................................................................................................................................ 3
Vie et anatomie de la reine........................................................................................................................ 3
Qualité de la reine ..................................................................................................................................... 5
APICULTURE AU CANADA .............................................................................................................................. 9
L’hiver et ses défis .................................................................................................................................... 9
Les importations de reines ...................................................................................................................... 13
L’HIVERNAGE DES REINES EN BANQUES ....................................................................................................... 17
Origine et principes généraux ................................................................................................................. 17
Les conditions à respecter ....................................................................................................................... 19
Effets sur la qualité et la performance des reines .................................................................................... 23
OBJECTIFS ET HYPOTHÈSES .......................................................................................................................... 24
Objectifs .................................................................................................................................................. 24
Hypothèses .............................................................................................................................................. 24
CHAPTER 1: IMPACTS OF INDOOR MASS STORAGE OF TWO DENSITIES OF
HONEYBEE QUEENS (APIS MELLIFERA L.) DURING WINTER ON QUEEN
SURVIVAL, REPRODUCTIVE QUALITY AND COLONY PERFORMANCE ................... 26
1.1 RÉSUMÉ .................................................................................................................................................. 27
1.2 ABSTRACT .............................................................................................................................................. 28
1.3 INTRODUCTION ....................................................................................................................................... 29
iv1.4 MATERIALS AND METHODS..................................................................................................................... 32
1.4.1 Queen rearing and experimental colonies ...................................................................................... 32
1.4.2 Distribution of queens in colonies and winter preparation ............................................................. 32
1.4.3 Colony overwintering .................................................................................................................... 34
1.4.4 Queen survival, morphometrics and fertility .................................................................................. 34
1.4.5 Queen introduction in nucleus colonies ......................................................................................... 35
1.4.6 Statistical analyses ......................................................................................................................... 36
1.5 RESULTS ................................................................................................................................................. 37
1.5.1 Queen survival ............................................................................................................................... 37
1.5.2 Temperature and relative humidity inside bank colonies ............................................................... 38
1.5.3 Queen morphometrics and fertility after winter ............................................................................. 39
1.5.4 Queen acceptance and performance in nucleus colonies ............................................................... 42
1.5.5 Queen morphometrics and fertility after introduction in nucleus colonies .................................... 43
1.6 DISCUSSION ............................................................................................................................................ 45
1.6.1 Effect of queen density in bank colonies ....................................................................................... 46
1.6.2 Influence of banking on queen reproductive quality ...................................................................... 47
1.6.3 Influence of banking on queen acceptance and performance ......................................................... 49
1.6.4 Queen reproductive quality after introduction in nucleus colonies ................................................ 50
1.6.5 Study limitations ............................................................................................................................ 51
CONCLUSION ................................................................................................................................ 54
BIBLIOGRAPHIE .......................................................................................................................... 60
vListe des tableaux
Table 1. Queen survival in bank colonies at two different queen densities, 40 and 80 (N = 5
banks per group) from their creation until the end of the overwintering period (August
27, 2019 to April 16, 2020). Different letters next to the survival rate values indicate a
significant difference between the two groups of bank colonies (p ≤ 0.05).
viListe des figures
Figure 1. Les trois stades de développement du couvain chez Apis mellifera : œufs (à
gauche), larves (au milieu) et pupes (à droite, recouvertes d’un opercule de cire
foncée). Crédit photo : Naomie Bleau.
Figure 2. Représentation schématique de l’anatomie interne de l’abdomen d’une reine
d’Apis mellifera (adaptée de Spivak et al. 1994).
Figure 3. Pourcentage de mortalité hivernale des colonies d’abeilles au Québec et au
Canada entre les années 2004 et 2020 (MAPAQ 2021).
Figure 4. Boîte de transport de reines, contenant 20 cagettes de type Jz-Bz. Chaque cagette
contient une reine, plusieurs ouvrières accompagnatrices et une réserve de nourriture.
Crédit photo : Andrée Rousseau.
Figure 5. Cadre de type Langstroth modifié supportant plusieurs reines d’Apis mellifera
isolées individuellement dans des cagettes. Sur cette photo, 20 reines sont visibles au
centre du cadre, et 20 autres sont présentes de l’autre côté. Ce cadre est ensuite inséré
au centre d’une banque. Crédit photo : Mireille Levesque.
Figure 6. Schematic representation of the distribution of the 630 queens among the control
nucleus colonies (Control), the 40-queen bank colonies (40 QB) and the 80-queen bank
colonies (80 QB) on August 27, 2019. In bank colonies, queens are isolated in cages
(California Mini Queen Cages, Mann Lake Ltd., Hackensack Minnesota USA #HD-
398) and distributed evenly on both sides of the modified holding frames. Each frame
is inserted in the center of the upper brood chamber of the bank colony.
Figure 7. Mean daily temperature (°C) (A) and relative humidity (%) (B) measured in bank
colonies at two different queen densities (N = 5 per density), from September 4, 2019
to April 16, 2020. The solid vertical line indicates the moment when the colonies
entered the environmentally controlled room (15 °C and 40-50 % RH) on October 10,
2019. The dotted horizontal lines represent the average temperature (A) and relative
humidity (B) in the environmentally controlled room.
Figure 8. Abdomen width (mm) (A), abdomen length (mm) (B), body weight (mg) (C) and
ovary weight (mg) (D) (mean ± SE) of control queens, queens from 40 QB and 80 QB
(N = 10 per group). Values were measured in late April, after the 6-month
overwintering period. The scale of the y-axis of the A, B and C graphs does not start at
zero in order to adequately show the values and their standard errors. A statistical
difference from the control group is indicated by an asterisk above the estimate
(p ≤ 0.05).
Figure 9. Sperm viability (%) (A) and sperm count (x10⁶) (B) (mean ± SE) of control
queens, queens from banks with 40 QB and 80 QB (N = 10 per group). Values were
viimeasured in late April, after the 6-month overwintering period. No statistical difference
was observed between groups (p > 0.05).
Figure 10. Brood population (N eggs, larvae and pupae) (mean ± SE) in nucleus colonies
with control queens (N = 8), queens from 40 QB (N = 5) and queens from 80 QB
(N = 7) measured on May 25, 2020, 12 days after the introduction of the queens. A
statistical difference from the control group is indicated by an asterisk above the
estimate (p ≤ 0.05).
Figure 11. Abdomen width (mm) (A), abdomen length (mm) (B), body weight (mg) (C)
and ovary weight (mg) (D) (mean ± SE) of control queens (N = 8), queens from 40 QB
(N = 5) and 80 QB (N = 7). Values were measured on May 25, 2020, 12 days after the
introduction of the queens. The scale of the y-axis of the A, B and C graphs does not
start at zero in order to adequately show the values and their standard errors. A
statistical difference from the control group is indicated by an asterisk above the
estimate (p ≤ 0.05).
Figure 12. Sperm viability (%) (A) and sperm count (x10⁶) (B) (mean ± SE) of control
queens (N = 8), queens from 40 QB (N = 5) and 80 QB (N = 7). Values were
measured on May 25, 2020, 12 days after the introduction of the queens. A statistical
difference from the control group is indicated by an asterisk above the estimate
(p ≤ 0.05).
viiiListe des abréviations
°C : degré Celcius
40 QB, 80 QB : Banques de 40 reines, Banques de 80 reines
AAC : Agriculture et Agroalimentaire Canada
ACPA, CAPA : Association canadienne des professionnels de l’apiculture, Canadian
Association of Professional Apiculturists
brglm : generalized linear model with binomial responses
CFIA : Canadian Food Inspection Agency
CO2 : dioxyde de carbone
CRAAQ : Centre de Référence en Agriculture et Agroalimentaire du Québec
CRSAD : Centre de Recherche en Sciences Animales de Deschambault
g, kg : gramme, kilogramme
lme : linear mixed-effects
MAPAQ : Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec
MBA : Manitoba Beekeepers’ Association
mm, km : millimètre, kilomètre
PCR : Petit coléoptère de la ruche
SE : Standard Error
μL, mL, L : microliter, milliliter, liter
ixGlossaire
Couvain : Ensemble des œufs, des larves et des pupes chez les insectes sociaux, comme
l’abeille mellifère.
Essaimage : Phénomène observé chez les abeilles lorsque la reine et environ la moitié de la
population d’abeilles (essaim) quittent la ruche afin de former une nouvelle colonie et ainsi
diviser la colonie en deux.
Fertilité: Potentiel reproductif d’un individu, c’est-à-dire le nombre de cellules
reproductrices qu’il peut produire.
Miellat : Substance sucrée excrétée par des insectes parasites vivant sur certaines plantes,
recueillie par les abeilles.
Nucléus : Petite colonie d’abeilles de quelques cadres comportant du couvain, des
provisions de nourriture, des ouvrières ainsi qu’une reine et qui se développera en colonie
mature.
Propolis : Résine provenant des bourgeons de certains arbres et récoltée par les abeilles
pour obturer les fentes des ruches.
Spermathèque : Organe de stockage des spermatozoïdes chez certains animaux femelles,
comme la reine-abeille.
Supersédure : Phénomène de remplacement naturel d’une reine-abeille sans essaimage, se
produisant lorsqu’une reine est malade, vieille ou inefficace.
xÀ ma mère Guylaine et à mon père Renaud pour
votre support inconditionnel.
xiRemerciements
Je tiens avant tout à remercier mon directeur Pierre Giovenazzo, qui a été un directeur en
or durant ce projet de maîtrise. Tu es toujours très présent et compréhensif envers tes
étudiants, surtout avec les deux dernières années de pandémie qui ont été difficiles pour le
moral et la motivation. Merci de m’avoir guidée durant les dernières années, tout en
m’accordant la confiance et l’indépendance dont j’avais besoin. C’est grâce à ta passion des
abeilles durant le cours d’Apidologie que j’ai poursuivi mes études dans ce domaine et je
t’en suis très reconnaissante. J’ai découvert un monde fascinant et qui me passionne moi
aussi maintenant.
Un merci particulier à Andrée Rousseau, qui m’a énormément appuyée tout au long du
projet, en particulier pour la partie expérimentale. Ton expertise et tes connaissances sur le
sujet m’ont permis de réaliser mes manipulations avec assurance. Merci aussi d’avoir fait
partie de mon comité d’évaluation et d’avoir apporté tes commentaires aux différentes
étapes de ce projet.
Merci aussi à tous les autres membres du personnel du CRSAD : Marilène Paillard,
Georges Martin, Mickaël Benoît, Marc-André Corriveau et Martine Bernier, qui
m’ont formée en apiculture et ont contribué aux travaux sur le terrain. Un merci spécial à
Émile Houle et à ses habiletés manuelles, qui ont été indispensables à la réalisation de ce
projet.
Merci à Stéphane Boudreau d’avoir accepté de faire partie de mon comité d’évaluation
tout au long de ma maîtrise et de m’avoir fait part de tes commentaires. Merci à Benjamin
Dainat d’avoir accepté d’évaluer mon mémoire. Merci également à Gaétan Daigle pour
les conseils sur mes analyses statistiques et à Karen Grislis pour les corrections de l’article
en anglais.
xiiMerci beaucoup à mes collègues et amies étudiantes Marie-Lou Morin, Ségolène
Maucourt et Laurence Plamondon. J’ai eu un grand plaisir à vous côtoyer et à me lier
d’amitié avec vous au cours de ces dernières années. Vous avez rendu mon parcours au
deuxième cycle très plaisant et vous m’avez grandement motivée. Merci aussi aux autres
étudiants que j’ai connus grâce au travail en apiculture, Philippe L’Écuyer, Jessica
Tremblay et Justine Sénéchal.
Merci aux partenaires financiers grâce auxquels le projet a pu se concrétiser : le Ministère
de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ) (Programme
Innov’Action Agroalimentaire Volet 1), l’Association canadienne des professionnels de
l’apiculture (ACPA) et le Conseil de recherches en sciences naturelles et en génie du
Canada (CRSNG). Merci à l’ACPA pour l’octroi du prix CAPA Student Merit Award 2021.
Finalement, un merci très spécial à tous mes proches, sans qui je n’aurais pas pu réaliser ce
projet. Merci à ma mère Guylaine pour son écoute formidable, à mon père Renaud de
m’avoir permis de me changer les idées en dépensant mon énergie au tennis et à mon frère
Pascal pour les découvertes musicales qui ont accompagné la rédaction de ce mémoire.
Merci à mes ami(e)s les empanadas (Béatrice, Camille, Blanche, Audrey et
accessoirement Hugh Grant), les lapines (Gabrielle et Audrey), mon moine préféré
Vincent, mes biologistes préférés (Louis-Philippe et Marie-Pier) et ma plus vieille amie
(dans tous les sens) Maialu. Vous m’avez tous et toutes permis de garder le moral en ces
temps pandémiques et je vous adore.
xiiiAvant-propos
Ce manuscrit a été rédigé sous la forme de mémoire avec insertion d’article. Le chapitre 1
de ce mémoire est donc un article scientifique rédigé en anglais. Ce chapitre suit une
introduction en français, correspondant à une revue de littérature, et est suivi par une
conclusion générale en français, revenant sur les résultats et les observations discutés au
chapitre 1. Le chapitre 1 s’intitule « Impacts of indoor mass storage of two densities of
honeybee queens (Apis mellifera L.) during winter on queen survival, reproductive quality
and colony performance ». Il n’est pas encore publié dans la littérature scientifique, mais
sera soumis à la revue « PLoS One » à la suite du dépôt final de ce mémoire.
L’auteure principale (M.L.) a rédigé ce manuscrit et a récolté et analysé les données. La
conceptualisation, la méthodologie et l’administration du projet ont été faites par M.L. en
collaboration avec Pierre Giovenazzo (P.G), Ph. D. et Andrée Rousseau (A.R). P.G. est
chercheur et professeur au département de biologie à l’Université Laval et titulaire de la
Chaire de leadership en enseignement des sciences apicoles, et A.R est chercheure au
Centre de recherche en sciences animales de Deschambault. A.R. a aussi grandement aidé à
la récolte des données. P.G. a également contribué à la révision et l’édition de ce manuscrit
avec M.L., a supervisé l’entièreté du projet en tant que directeur de recherche et a assuré
l’acquisition du financement.
Il est aussi important de souligner que la pandémie de COVID-19 survenue au printemps
2020 a entraîné plusieurs modifications au protocole initial de mon projet. Effectivement,
les manipulations concernant la performance des reines-abeilles en colonie devaient se
dérouler pendant tout l’été 2020. Nous aurions évalué la performance des colonies chaque
mois à l’aide de plusieurs critères de performance, mais les mesures gouvernementales et
universitaires établies au printemps 2020 nous ont obligés à réduire notre protocole afin que
la réalisation de ce projet soit autorisée.
xivIntroduction
L’abeille mellifère
Historique
À ce jour, plus de 20 000 espèces d’abeilles ont été répertoriées à travers le monde. Parmi
elles, celles du genre Apis (Hymenoptera : Apoidea : Apidae: Apinae : Apini) sont les plus
notables puisqu’elles représentent les abeilles produisant du miel en grande quantité. En
apiculture, l’abeille mellifère (Apis mellifera L.) (Ruttner 1988) est l’espèce la plus utilisée,
tant pour sa production de miel que pour ses services de pollinisation. Originaire d’Afrique,
d’Europe et du Moyen-Orient, elle a ensuite été introduite à plusieurs endroits dans le
monde à des fins d’exploitation et se retrouve désormais sur tous les continents sauf
l’Antarctique (Whitfield et al. 2006). De nos jours, l’abeille mellifère est indispensable à
notre économie et notre environnement, puisque le tiers des cultures mondiales dépendent
de sa pollinisation (Aizen et Harder 2009). Au Canada, la valeur annuelle de la pollinisation
des terres agricoles attribuable à l’abeille domestique est estimée à 2,5 milliards de dollars
et celle de la production de miel à environ 200 millions de dollars (AAC 2021).
La colonie d’abeilles
Une des caractéristiques principales de cet insecte est son eusocialité. Cet attribut définit
son organisation sociale et implique une cohabitation des générations, une coopération dans
le soin aux jeunes, une forte cohésion entre les membres et un système de castes
reproductives. Chez l’abeille mellifère, ces castes sont représentées par les ouvrières, les
faux-bourdons et la reine (Haydak 1943, Winston 1991, Hartfelder et Engels 1998).
La caste des ouvrières représente la grande majorité de la colonie et comporte des dizaines
de milliers d’individus. Elles proviennent d’un œuf pondu et fécondé par la reine, et sont
par conséquent diploïdes (Dadant 1976). Ce sont des femelles stériles puisque leurs organes
1reproductifs sont atrophiés. Elles accomplissent la plupart des tâches essentielles à
l’organisation de la colonie et la répartition de ces tâches est déterminée par leur âge
(Huang et Robinson 1996). Les plus jeunes abeilles sont responsables des activités se
déroulant à l’intérieur de la ruche, comme le nettoyage des cellules, les soins envers la
reine, la ventilation de la ruche, la construction des rayons de cire, l’alimentation des
adultes ainsi que l’alimentation du couvain (Figure 1), soit l’ensemble des œufs et des
larves d’abeilles. Les plus vieilles abeilles effectuent quant à elles les activités se déroulant
à l’entrée ou à l’extérieur de la ruche, telles que la défense de la colonie ainsi que la récolte
de nectar, de pollen, de propolis, de miellat et d’eau (Michener 1974, Winston 1991).
Figure 1. Les trois stades de développement du couvain chez Apis mellifera : œufs (à
gauche), larves (au milieu) et pupes (à droite, recouvertes d’un opercule de cire foncée).
Crédit photo : Naomie Bleau.
La caste des faux-bourdons correspond aux mâles de la colonie et comporte des centaines à
des milliers d’individus. Contrairement aux femelles, ils sont haploïdes puisqu’ils sont issus
d’œufs non fécondés. Leur fonction principale dans la colonie est la fécondation des reines,
à laquelle ils ne survivent pas car leur appareil reproducteur ainsi que le dernier segment de
l’abdomen restent accrochés à la reine durant la reproduction (Winston 1991, Parker et al.
2010). Ils ne participent pas à la récolte de nourriture ni aux autres tâches comme le font les
ouvrières, c’est pourquoi ils sont rejetés hors de la ruche à l’automne, afin de limiter la
2consommation des réserves de nourriture nécessaires à la survie hivernale des colonies
(Wilson 1971).
La dernière caste est représentée par la reine. C’est la seule femelle fertile de la colonie, ce
qui la rend cruciale au fonctionnement d’une colonie d’abeilles mellifères (Winston 1991,
Parker et al. 2010). C’est donc à elle que revient le rôle de pondre des œufs et ainsi
d’assurer la croissance de la colonie et le renouvellement rapide de la progéniture
d’ouvrières et de faux-bourdons (Dadant 1976, Winston 1991). La reine régule également
la plupart des activités se déroulant au sein de la colonie grâce aux phéromones qu’elle
dégage. Chez l’abeille mellifère, les phéromones sont des substances chimiques produites
par des glandes se situant au niveau de la tête, des pattes et de l’abdomen. Elles se
transmettent par diffusion dans l’air et sont perçues par les antennes de l’insecte, par
contact physique entre les individus ou encore par trophallaxie, soit le transfert de
nourriture entre les abeilles (Plettner et al. 1997, Maisonnasse 2010, Bortolotti et Costa
2014). Les phéromones conditionnent presque toute la dynamique observée dans une
colonie d’abeilles et les plus importantes sont produites par la reine. Elles ont une grande
influence sur divers comportements observés au sein d’une colonie : inhibition du
développement ovarien des ouvrières, de l’élevage de mâles et de l’élevage de nouvelles
reines, et stimulation de l’élevage de couvain, de la construction de rayons de cire et du
butinage (Winston et al. 1989, Traynor et al. 2014). La reine remplie donc des fonctions
essentielles à la cohésion ainsi qu’à l’organisation de la colonie.
La reine
Vie et anatomie de la reine
Comme les ouvrières, la reine provient d’un œuf fécondé. Chaque œuf fécondé a donc le
potentiel de se développer en ouvrière ou en reine. La différenciation entre ces deux castes
se fait par un facteur alimentaire. Les larves d’ouvrières sont nourries de gelée royale et de
3gelée nourricière, un mélange de miel et de pollen, durant les trois premiers jours de leur
développement, pour ensuite être nourries uniquement de gelée nourricière. Les larves de
reines sont pour leur part nourries exclusivement de gelée royale durant leur
développement, une alimentation riche en divers nutriments comme l’hormone juvénile et
le royalactine et qui permet le bon développement de son système reproducteur (Haydak
1943, Hartfelder et Engels 1998, Wright et al. 2018). La reine est aussi élevée dans une
cellule royale, plus vaste qu’une cellule d’ouvrière. Après 16 jours de développement, la
reine émerge de sa cellule royale. Puisque les ouvrières élèvent plusieurs reines en même
temps, la première reine à naître détruit les cellules concurrentes en les piquant ou en les
abîmant avec ses mandibules. Si plusieurs reines naissent en même temps, elles se battent
jusqu’à ce qu’il n’en reste qu’une (Winston 1991, Gilley 2001).
Cette nouvelle reine, communément appelée une reine vierge, est petite et n’émet pas
beaucoup de phéromones. Avant de pouvoir jouer pleinement son rôle dans la colonie, elle
doit se faire féconder. Dans les premières semaines de sa vie, elle effectue alors un vol
nuptial afin de se reproduire avec une quinzaine de faux-bourdons (Adams et al. 1977).
Lorsque la reine retourne à sa colonie après son vol nuptial, entre 80 et 100 millions de
spermatozoïdes sont présents dans les canaux de son système reproducteur. Cependant,
seulement environ 5 % de ceux-ci migrent jusqu’à la spermathèque, un organe chez la reine
permettant de stocker les spermatozoïdes jusqu’à leur utilisation (Woyke 1962, Klowden
2013). Cet organe est doté d’une fonction nourricière, ce qui permet d’assurer la survie des
spermatozoïdes durant toute la vie de la reine (Verma 1973, Klenk et al. 2004, Wegener et
al. 2013, Tarpy et Olivarez Jr. 2014).
Suite à sa fécondation, la reine subit plusieurs changements physiologiques qui lui
permettront de remplir ses fonctions au sein de sa colonie. Notamment, ses ovaires
atteignent leur maturité et son abdomen s’élargit et s’allonge par le fait même (Figure 2).
Les phéromones produites par la reine deviennent aussi plus abondantes, ce qui lui permet
d’être reconnue par les ouvrières et de réguler les activités de la colonie (Winston et al.
41989, Richard et al. 2007). Quelques jours après sa fécondation, elle commence à pondre
des œufs. Pour pondre un œuf, un ovule est d’abord libéré et circule par un canal appelé
oviducte. Pour que l’œuf se développe en femelle, l’ovule doit être fécondé par un
spermatozoïde, que la reine libère de sa spermathèque. Si l’ovule n’est pas fécondé, l’œuf
se développera en mâle (Dadant 1976). La reine pondra des œufs et vivra jusqu’à ce que le
contenu de sa spermathèque soit épuisé, ce qui peut prendre de 3 à 5 ans (Winston 1991).
Figure 2. Représentation schématique de l’anatomie interne de l’abdomen d’une reine
d’Apis mellifera (adaptée de Spivak et al. 1994).
Qualité de la reine
Puisque la reine joue un rôle essentiel au sein d’une colonie, il est important qu’elle soit de
bonne qualité afin d’assurer la survie et la performance d’une ruche. La qualité d’une reine
se définit par son potentiel reproductif quantitatif et qualitatif, et celui-ci s’exprime de
plusieurs façons au sein d’une colonie (Amiri et al. 2017). Notamment, il se manifeste par
l’intensité et le patron de sa ponte. Une reine performante pond en moyenne 1500 œufs par
jour, et lorsque les conditions sont favorables, ce nombre peut dépasser 2000 œufs par jour
(Dadant 1976, Winston 1991). De plus, une reine de qualité pond ses œufs en zones
concentriques compactes. Un cadre de couvain idéal contient le plus vieux couvain au
centre et le plus jeune en périphérie, et le nombre d’alvéoles vides représente moins de
520 % de la surface de couvain (VanEngelsdorp et al. 2013). Si la quantité ou la qualité de la
ponte d’une reine est réduite, cela limitera l’expansion de la colonie, et par le fait même sa
production de miel et son potentiel de pollinisation (Woyke 1971, Pettis et al. 2016). Elle
deviendra aussi plus susceptible aux maladies et à la période hivernale suivante (Free et
Racey 1968, Döke et al. 2019).
La qualité de la reine peut également se mesurer par la proportion de couvain d’ouvrières et
de couvain de faux-bourdons. De façon générale, le couvain de faux-bourdons représente
environ 10 % du couvain d’une colonie. Une surabondance de couvain de faux-bourdons
peut témoigner de l’épuisement des spermatozoïdes de la spermathèque de la reine. Dans ce
cas-ci, la reine ne peut plus pondre d’œufs fécondés et est qualifiée de « bourdonneuse »
(Dadant 1976, Winston 1991). Ceci retardera considérablement le développement d’une
colonie, puisqu’une nouvelle reine devra se faire élever et se faire féconder avant qu’une
nouvelle génération d’ouvrières puisse être produite.
Outre la ponte de la reine, plusieurs autres éléments sont aussi considérés comme des
indicateurs de la qualité d’une reine puisqu’ils sont souvent reliés à sa performance en
colonie. Notamment, diverses études considèrent le poids d’une reine comme un prédicteur
de son potentiel reproductif et donc de sa performance en colonie (Nelson 1989, Akyol et
al. 2008). Le poids d’une reine est souvent corrélé au poids de ses ovaires, au nombre et à
la taille de ses ovarioles, au stade de développement de ses ovules, au diamètre de sa
spermathèque et au nombre de spermatozoïdes stockés dans sa spermathèque (Akyol et al.
2008, Delaney et al. 2011, Tarpy et al. 2012). Le poids de la reine est donc généralement un
bon indicateur de son attraction auprès des ouvrières de sa colonie et de son potentiel
reproductif (Masry et al. 2015). Cette relation n’est toutefois pas toujours constante,
puisque le poids de la reine peut varier lors de périodes de stress ou avec la présence de
maladies (Kahya et al. 2008). De plus, le poids de la reine varie temporellement selon
l’intensité de sa ponte. Lorsque sa ponte cesse ou diminue, par exemple pendant la période
6hivernale en région nordique, ses ovaires rétrécissent et son poids diminue (Shehata et al.
1981).
La taille et le contenu de la spermathèque d’une reine peuvent aussi permettre d’estimer le
potentiel reproductif d’une reine. Une spermathèque plus large contient généralement plus
de spermatozoïdes, ce qui augmente le potentiel reproductif et l’espérance de vie de la reine
(Tarpy et al. 2012, Collins et Pettis 2013, Hatjina et al. 2014). La viabilité des
spermatozoïdes stockés dans sa spermathèque est également un paramètre important dans
l’évaluation de la qualité d’une reine. En effet, il faut qu’une grande proportion des
spermatozoïdes, soit plus de 80 %, soit viable pour qu’une reine ait un potentiel reproductif
élevé (Den Boer et al. 2009, Tarpy et al. 2012, Amiri et al. 2017).
La qualité d’une reine se manifeste donc de différentes façons, et celle-ci dépend de
plusieurs facteurs biotiques et abiotiques. Elle peut entre autres être influencée par la
nutrition qu’elle reçoit durant son développement. Si une reine n’est pas nourrie assez
longtemps de gelée royale, son système reproducteur se développera moins bien, elle sera
plus petite et dégagera moins de phéromones (Laidlaw et Eckert 1962). Ceci peut
notamment se produire lors du phénomène de supersédure, c’est-à-dire l’élevage de
nouvelles reines par une colonie pour remplacer une reine trop vieille, malade ou
inefficace. En effet, si une reine doit être remplacée, elle n’est parfois plus capable de
pondre des œufs. Les ouvrières doivent alors transférer de jeunes larves, idéalement écloses
depuis moins de 36 heures, dans des cellules royales. L’utilisation de très jeunes larves
garantit qu’elles seront nourries de gelée royale pendant une période suffisante (Fukuda et
Sakagami 1968, Woyke 1971, Visscher 1986). Par contre, si les larves disponibles sont trop
âgées, la reine sera nourrie moins longtemps de gelée royale, ce qui compromettra son
développement. La nutrition reçue par la reine peut aussi être influencée par la force et les
ressources de la colonie. Il faut une grande quantité de jeunes abeilles et de réserves de
pollen dans une colonie afin de produire suffisamment de gelée royale (Prešern and Škerl
2019). Les reines sont alors de meilleure qualité lorsqu’elles sont élevées à des périodes où
les ressources environnementales sont abondantes.
7La période de fécondation de la reine a également un effet sur sa qualité (Adams et al.
1977, Tarpy et al. 2014). En effet, plusieurs conditions sont nécessaires pour assurer
l’efficacité du vol nuptial de la reine. La température doit être tempérée, idéalement entre
20 et 25 °C, le ciel doit être dégagé et la vitesse du vent doit être faible. La quantité de
mâles disponibles dans un rayon d’environ trois kilomètres (km) est aussi une condition
déterminante d’une bonne fécondation (Ruttner 1956, Woyke et Ruttner 1958, Neumann et
al. 1999, Boes 2010). Si ces conditions ne sont pas présentes, la fécondation de la reine ne
sera pas optimale. Elle doit alors effectuer un ou plusieurs autres vols de fécondation,
jusqu’à ce que sa spermathèque soit remplie. Toutefois, si des conditions défavorables se
maintiennent pendant une période prolongée, la reine ne sera pas en mesure de remplir sa
spermathèque, ce qui affectera sa qualité. En effet, plusieurs études montrent que la
quantité de spermatozoïdes stockés dans la spermathèque d’une reine est liée à son
potentiel reproductif, son attraction auprès des ouvrières, la division des tâches dans une
colonie et la communication entre les abeilles (Schlüns et al. 2005, Richard et al. 2007,
Tarpy et al. 2012). Le nombre de mâles avec lesquels une reine s’accouple est aussi un
facteur important puisqu’il détermine la diversité génétique d’une colonie. Une grande
diversité génétique permet à une colonie de s’adapter plus facilement à des changements
biotiques et abiotiques, tels que les variations de température ou la présence de parasites ou
d’agents pathogènes (Mattila et Seeley 2007, Oldroyd et Fewell 2007, Amiri et al. 2017).
La capacité reproductive d’une reine est également influencée par la qualité du sperme des
faux-bourdons avec lesquels elle s’accouple. Entre autres, la viabilité, la motilité ainsi que
la taille des spermatozoïdes des mâles affectent toutes le pourcentage de spermatozoïdes
migrant jusqu’à la spermathèque de la reine (Verma 1973, Gençer et Kahya 2011).
Le potentiel reproductif d’une reine est aussi grandement régi par son génome. En effet,
diverses études montrent que la génétique d’une reine a une influence significative sur
plusieurs paramètres de sa qualité : son poids, le nombre d’ovarioles de ses ovaires, la taille
de sa spermathèque, le nombre de spermatozoïdes dans sa spermathèque et le temps qu’elle
met avant de commencer à pondre (Hoopingarner et Farrar 1959, Dodologlu et Gene 2003,
Facchini et al. 2021). La génétique d’une reine a aussi un grand impact sur la performance
8d’une colonie. En effet, divers critères de performance d’une colonie sont influencés par la
génétique et sont héréditaires : la production de miel, le développement de la colonie au
printemps, le comportement hygiénique ainsi que la consommation de nourriture pendant
l’hiver (Maucourt et al. 2020). La qualité d’une reine est alors fortement dépendante de son
bagage génétique.
Apiculture au Canada
Au Canada, l’industrie apicole représente une partie importante du secteur agricole, tant
pour sa production de miel et d’autres produits de la ruche que pour les services de
pollinisation qu’elle procure aux cultures de fruits, de légumes et de plantes comme le
canola et le soya (Page et Darrach 2016). La production de miel et la pollinisation des
cultures génèrent respectivement 200 millions et cinq milliards de dollars chaque année.
L’apiculture est aussi en croissance constante depuis de nombreuses années. Depuis 2007,
le nombre de colonies au pays a augmenté de 35 %, se situant présentement à plus de
750 000 colonies, réparties parmi environ 10 000 apiculteurs (Page et Darrach 2016, AAC
2021). L’expansion de cette industrie est tout de même restreinte par plusieurs facteurs,
notamment par la période hivernale.
L’hiver et ses défis
Les activités apicoles varient grandement selon les conditions environnementales d’une
région. La récolte de nourriture ainsi que la ponte de la reine d’une colonie dépendent de
plusieurs facteurs externes, tels que les saisons et la disponibilité de plantes mellifères à
proximité. Dans les régions tropicales, subtropicales et là où les hivers sont doux, la ponte
de la reine et l’élevage du couvain n’arrêtent pas ou peu. L’apiculture peut donc se
pratiquer toute l’année. Cependant, dans les régions où les changements saisonniers sont
très prononcés, comme au Canada, les activités apicoles sont suspendues pendant une
longue période chaque année (Dunham 1930, Abou-Shaara et al. 2017).
9En effet, lorsque la température extérieure baisse sous environ 15 °C, les abeilles se
rassemblent à l’intérieur de la ruche afin de se préparer à l’hiver (Watmough et Camazine
1995, Sumpter et Broomhead 1999) et la reine ralentit graduellement sa ponte. Pendant la
période hivernale, la reine ne pond plus ou presque plus (Seeley et Visscher 1985, Mattila
et al. 2001). Durant la saison estivale, les ouvrières ont une durée de vie d’une quarantaine
de jours au maximum. Toutefois, les abeilles nées en automne peuvent survivre jusqu’à six
mois, ce qui permet de compenser l’absence de nouveau couvain pendant l’hiver (Merz et
al. 1979). Ces ouvrières possèdent des réserves lipidiques et protéiques plus élevées
(Münch et Amdam 2010), mais les autres raisons expliquant cette durée de vie prolongée
sont encore mal comprises. La ponte de la reine reprend généralement peu après le solstice
d’hiver (Seeley et Visscher 1985).
Contrairement à plusieurs insectes, les colonies d’abeilles mellifères n’hibernent pas
pendant l’hiver, mais modifient leurs comportements, leurs activités et leur métabolisme.
Elles restent confinées à l’intérieur de la ruche et consomment les réserves de nourriture
qu’elles ont accumulées pendant l’été. Si tout le miel a été récolté par les apiculteurs, les
colonies doivent être nourries avec du sirop avant l’hiver. Afin de conserver une
température supérieure à 15 °C et d’environ 30 °C lorsque du couvain est présent, les
abeilles de la colonie se regroupent et forment progressivement une grappe dont la densité
varie selon la température extérieure (Melathopoulos 2007). Au centre de cette grappe, les
abeilles maintiennent une température constante d’environ 32 °C en agitant les muscles de
leurs ailes. La reine se situe au milieu de cette grappe et se fait nourrir par les ouvrières tout
l’hiver (Southwick 1985, Winston 1991, Parker et al. 2010). Cette adaptation n’est
cependant pas suffisante pour survivre à l’hiver au Canada. Les apiculteurs doivent alors
hiverner leurs colonies, soit à l’extérieur en les recouvrant de matériau isolant à l’extérieur
ou en les entreposant dans un caveau d’hivernage à une température d’environ 5 °C. Cette
température assure un rythme optimal de la consommation des réserves de nourriture par
les abeilles et favorise ainsi leur survie (Desai et Currie 2016). Au printemps, lorsque les
températures augmentent et que les ressources alimentaires deviennent disponibles, les
10isolants sont retirés ou les colonies sont replacées à l’extérieur. À ce moment, les colonies
profitent de ces conditions afin d’accroître leur population.
La période hivernale rend les colonies d’abeilles mellifères plus vulnérables et plusieurs
d’entre elles n’y survivent pas. Au Canada, entre 18 et 45 % des colonies meurent chaque
hiver, la moyenne se situant à 33 % (Bixby et al. 2018) (Figure 3). Les conditions
météorologiques défavorables sont le principal facteur associé à ces mortalités, étant donné
les variations extrêmes de température qui se prolongent parfois jusqu’à tard au printemps.
La qualité des reines est aussi une cause importante des pertes de colonies hivernales. Une
reine de moindre qualité se manifeste souvent par une colonie affaiblie avant d’entrer en
hivernage. Le manque de nourriture est aussi responsable d’une grande partie des mortalités
hivernales. Cette famine peut être provoquée par un nourrissage insuffisant par les
apiculteurs, par un stockage inefficace des réserves de nourriture par les abeilles ou encore
par une production précoce de couvain entraînant un rythme de consommation accrue de
leur réserve de nourriture. Des températures froides prolongées peuvent aussi accélérer la
consommation de nourriture par les colonies (ACPA 2018). En outre, les pertes hivernales
peuvent être provoquées par la faiblesse des colonies à l’automne. Celle-ci peut être causée
par divers facteurs tels que la présence d’agents pathogènes, une exposition prolongée aux
pesticides ou une construction tardive des colonies. Finalement, le mauvais contrôle de
l’acarien parasite Varroa destructor (varroa) est responsable d’une grande partie des
mortalités hivernales (Beyer et al. 2018). Les pertes hivernales représentent donc un aspect
crucial et inévitable de la pratique de l’apiculture au Canada.
11Figure 3. Pourcentage de mortalité hivernale des colonies d’abeilles au Québec et au
Canada entre les années 2004 et 2020 (MAPAQ 2021).
Afin de remplacer ces nombreuses pertes, ou simplement pour augmenter leur cheptel, les
apiculteurs doivent construire de nouvelles colonies d’abeilles au printemps. Ils peuvent
alors diviser leurs colonies survivantes pour obtenir des « nucléi ». Ce terme réfère à des
petites colonies, construites en prélevant quelques cadres de couvain et de nourriture (miel
et pollen) ainsi que des abeilles d’une ruche mère et en les transférant dans une nouvelle
ruche. Ces nucléi se développeront en colonies matures au cours de la saison. Pour les
compléter, une reine doit y être introduite (Dadant 1976). Pour ce faire, l’apiculteur insère
une cagette contenant une reine et une source de nourriture entre les deux cadres centraux
du nucléi. Les ouvrières libèrent la reine tranquillement en grugeant cette nourriture qui
empêche l’accès direct à la reine, ce qui laisse le temps aux abeilles de s’habituer aux
phéromones de la reine.
Cependant, les conditions météorologiques du Canada limitent l’élevage de reines tôt au
printemps. En effet, plusieurs conditions sont nécessaires afin d’élever des reines : des
12Vous pouvez aussi lire

















































