Génomique des populations et association génotype-phénotype des écotypes de touladi du lac Supérieur - Mémoire Alysse Perreault-Payette Maîtrise ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Génomique des populations et association génotype-
phénotype des écotypes de touladi du lac Supérieur
Mémoire
Alysse Perreault-Payette
Maîtrise en biologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Alysse Perreault-Payette, 2016
Génomique des populations et association génotype-
phénotype des écotypes de touladi du lac Supérieur
Mémoire
Alysse Perreault-Payette
Sous la direction de :
Louis Bernatchez, directeur de recherche
Pascal Sirois, codirecteur de recherche
Résumé
L’apparition et le maintien d’écotypes adaptés à différentes niches écologiques, en situation
de sympatrie, est régit par une multitude de facteurs. Ceux-ci sont essentiels pour la
compréhension des processus évolutifs impliqués mais aussi pour la gestion et la
conservation des populations en question. Le touladi (Salvelinus namaycush) est un
salmonidé reconnu pour la présence d’écotypes liée à l’utilisation des ressources et de
l’habitat à travers l’Amérique du Nord. Un total de quatre écotypes a été décrit vivant dans
le lac Supérieur, se différenciant par l’habitat utilisé, l’alimentation, la morphologie ainsi
que l’ostéologie. L’objectif principal de la présente étude était de quantifier l’étendue de la
différentiation génétique entre les différents sites d’échantillonnage ainsi qu’entre les
différents écotypes. Un second objectif était d’identifier des marqueurs potentiellement sous
sélection entre les différents écotypes reflétant de possibles adaptations locales. Pour ce
faire, un total de 486 individus, représentant les quatre écotypes pour chacun des quatre sites
d’échantillonnages, a été génotypé à 6822 SNPs (polymorphisme de nucléotide simple). De
plus, des analyses morphométriques ont été effectuées afin de caractériser l’ampleur de la
divergence morphologique entre les écotypes à chacun des sites. Les résultats ont montré
une différentiation génétique, bien que faible, plus prononcée entre les sites
d’échantillonnage qu’entre les écotypes à chacun de ces sites. Des indices indiquant la
présence de sélection divergente ont aussi été décelés entre les écotypes ou en association
avec des variations morphologiques, dont certains marqueurs représentant des traits
importants dans la divergence des différents écotypes. Les résultats de cette étude
permettront une meilleure gestion et conservation des populations de touladi du lac
Supérieur en plus d’éclairer le choix possible de populations sources pour l’ensemencement
des autres Grands Lacs.
iii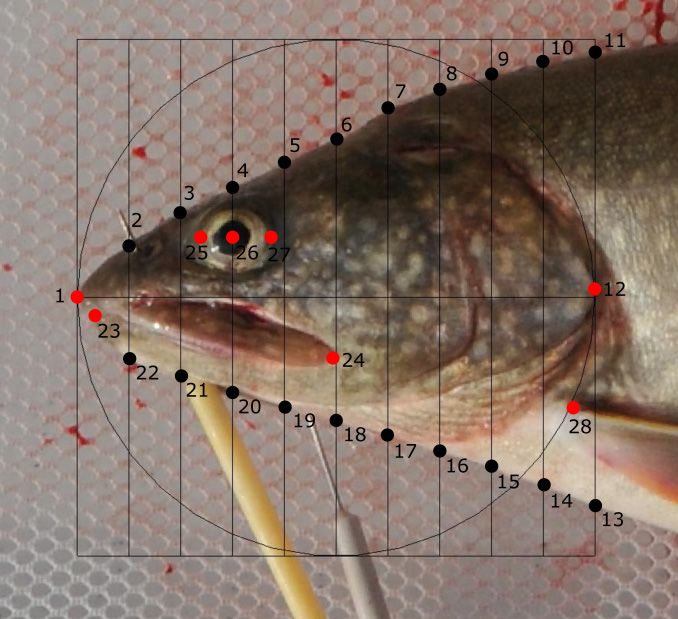
Abstract
Understanding the emergence and maintenance of sympatric ecotypes adapted to various
trophic niches is a central topic in evolutionary biology, and also has implications for
conservation and management. Lake Trout (Salvelinus namaycush) is renowned for the
occurrence of phenotypically distinct ecotypes linked to resource and habitat use throughout
North America. A total of four ecotypes have been described in Lake Superior that differ in
terms of habitat, diet, morphology and osteology. The principal objective of this study was
to quantify the extent of genetic differentiation among sampling sites and among ecotypes.
The secondary objective was to identify markers potentially under divergent selection
among the four ecotypes that may underlie local adaptation. To this end, a total of 486
individuals were genotyped at 6822 SNPs (single nucleotide polymorphism). In addition,
these analyses were conducted alongside morphometric analyses to characterise the extent of
morphological divergence among ecotypes within each sampling site. Results reveal that
overall genetic differentiation is weak and is higher among sites than among ecotypes within
each site. Moreover, we found evidence for divergent selection among ecotypes, and in
some instances in association with morphological variation. These markers represent
ecologically important traits linked to ecotype divergence. Results from this study will
benefit management and conservation practices, and will guide the choice of source
populations for stocking in the Great Lakes.
iv
Table des matières
Résumé .................................................................................................................................................iii
Abstract ................................................................................................................................................ iv
Table des matières ................................................................................................................................. v
Liste des tableaux ................................................................................................................................ vii
Liste des figures ................................................................................................................................. viii
Remerciements ...................................................................................................................................... x
Avant-propos ........................................................................................................................................ xi
Introduction ........................................................................................................................................... 1
L’apparition de nouvelles espèces .................................................................................................... 1
Parallélisme dans la divergence phénotypique................................................................................. 3
Le touladi ........................................................................................................................................... 4
En Amérique du Nord .................................................................................................................... 4
Dans le lac Supérieur ..................................................................................................................... 5
L’origine et le maintien des écotypes ........................................................................................... 6
Problématique ................................................................................................................................... 7
Bouleversement du système aquatique ....................................................................................... 7
Plan de réintroduction des poissons de fonds .............................................................................. 8
Objectifs ............................................................................................................................................ 9
Chapter I: Investigating the Extent of Parallelism in Morphological and Genomic Divergence among
Lake Trout Ecotypes in Lake Superior ............................................................................................... 10
Résumé ............................................................................................................................................ 11
Abstract ........................................................................................................................................... 13
Introduction .................................................................................................................................... 14
Methods .......................................................................................................................................... 17
Results ............................................................................................................................................. 26
Discussion ........................................................................................................................................ 42
Acknowledgements ......................................................................................................................... 51
Conclusion........................................................................................................................................... 52
Analyses morphologiques ............................................................................................................... 52
v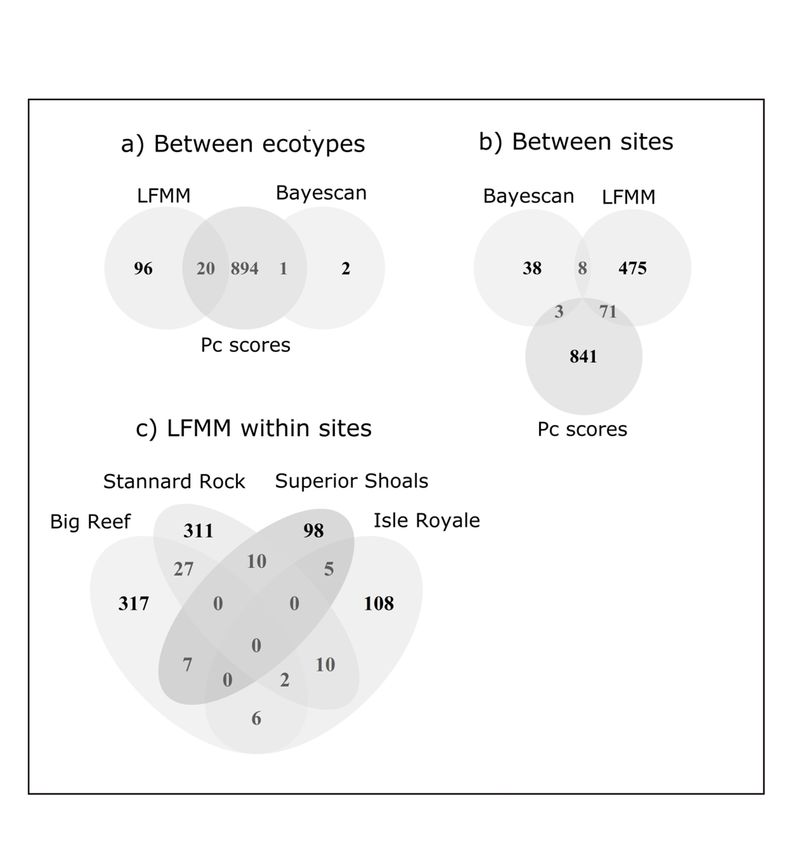
Analyses génomiques et divergence adaptative ............................................................................. 53
Différences historiques et impacts anthropogéniques ................................................................... 54
Implications pour la gestion et la conservation .............................................................................. 56
Limitations et perspectives futures ................................................................................................. 57
Bibliographie ....................................................................................................................................... 59
Annexe 1 ............................................................................................................................................. 67
vi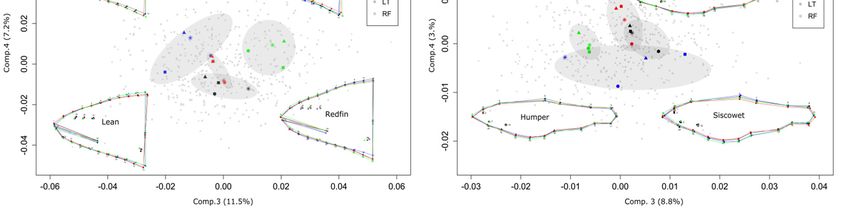
Liste des tableaux
Table 1. Sampling site information and consensus analysis of body shape, head shape and visual
identification. Number of fish sampled per sampling site (N), year of collection and
coordinates is provided. Ecotypes were identified by consensus analysis of body shape (B)
and/or head shape (H) and/or visual identification (V). Fish for subsequent genetic analysis
were chosen based on ecotype consensus. Fish less than 430 mm long were removed prior
to the analysis……………………………………………………………………….......….19
Table 2. Models and parameters selected to assign an ecotype to each fish from the four sites
separately. The number of principal components used and the corresponding cumulative
percentage of explained variance are in brackets. Models chosen and parameterisation of
the covariance matrix are either identified as EII (spherical distribution, equal volume, and
equal shape), VEI (diagonal distribution, variable volume, and equal shape), VII (spherical
distribution, variable volume, equal shape) or EEI (diagonal distribution, equal volume,
and equal shape). The number of groups found (G), the corresponding bayesian
information criterion (BIC) and mean uncertainty are also listed for each model….…....29
Table 3. MANOVA on body and head shape to investigate the effect of the ecotype, the site of
origin, the sex and all interactions. Significant variables are in bold……………..…..…...30
Table 4. Between-group PCA analysis of group distance among ecotypes within and among sites
with 10000 permutation for body (below diagonal) and head (above diagonal) shape.
Ecotypes are: Siscowet (FT), Humper (HT), Lean (LT) and Redfin (RF). Group distances
with significant p-value (< 0.05) are followed by an asterix (*). Within sites analyses are
highlighted in gray and significant between site differences for the same ecotype are in
bold. ………………………………………………………………………………..….….31
Table 5. Number of SNPs remaining after each filtration step. Allelic imbalance corresponds to the
ratio of the number of sequences for the major allele on the number of sequences for the
minor allele……………………………………………………………………….….....33
Table 6. Population statistics estimated with 6822 SNPs: the observed heterozygosity (Ho), the
expected heterozygosity (He), the inbreeding coefficient (Gis), the effective population size
(Ne) and confidence interval in brackets, and the number of polymorphic loci (N). Genome-
wide diversity () and the increase in individual homozygosity relative to mean Hardy-
Weinberg expected homozygosity (Fh) was calculated on the dataset prior to filtration.
Effective population size for ecotypes with sample size < 15 individuals were not
calculated (NA)…………………………………………………………………..………...35
Table 7. Analysis of molecular variance (AMOVA) on 486 individuals and 6822 SNPs. Missing
data has been replaced by random picking in the overall pool of allele frequency…...….36
viiListe des figures
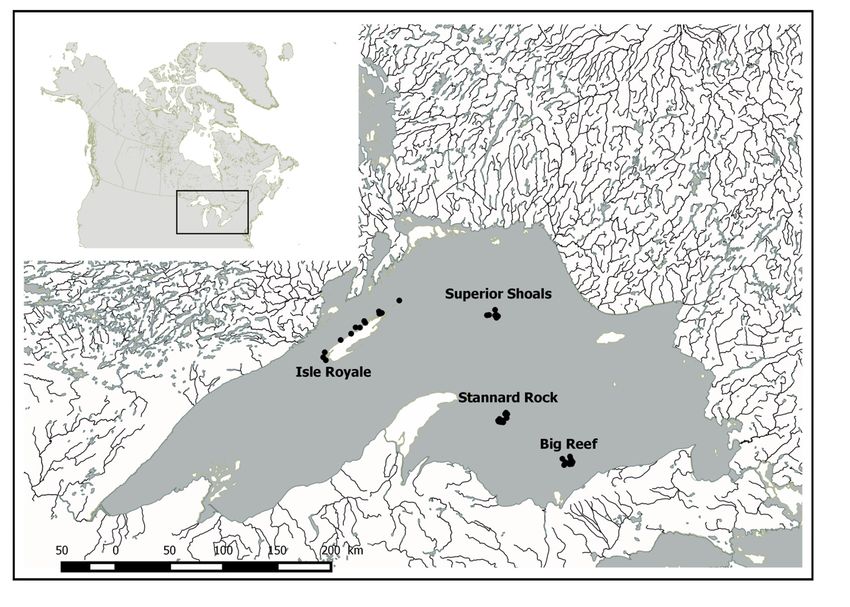
Figure 1. Map of Lake Superior sampling sites; Isle Royale, Superior Shoals, Stannard Rock and
Big Reef. Circles correspond to sampling locations for each site………………….…….18
Figure 2. Landmark and semi-landmark positions digitized on fish body and head. Homologous
landmarks are represented by red dots and semi-landmarks by black dots. (a) Sixteen
homologous landmarks and four semi-landmarks were placed on each fish body; (1) tip of
the snout, (2) posterior tip of the maxillary, (3) center of the eye, (4) top of the head above
the center of the eye, (5) posterior of neurocranium above the edge of opercula, (6) anterior
insertion of dorsal fin, (7) posterior insertion of dorsal fin, (8) anterior insertion of adipose
fin, (9) dorsal insertion of caudal fin, (10) midpoint of hypural plate, (11) ventral insertion
of caudal fin, (12) posterior insertion of anal fin, (13) anterior insertion of anal fin, (14)
anterior insertion of pelvic fin, (15) 50% of body length, (16) 40% of body length, (17)
30% of body length, (18) 20% of body length, (19) anterior insertion of pectoral fin, (20)
connection (isthmus) between branchiostegals. (b) Eight homologous landmarks and
twenty semi-landmarks were placed on each fish head; (1) tip of the snout, (2-11) ten
equally spaced semi-landmarks, (12) posterior edge of the opercula, (13-22) ten equally
spaced semi-landmarks, (23) tip of the lower jaw, (24) posterior tip of the maxillary, (25)
anterior edge of the eye, (26) center of the eye, (27) posterior edge of the eye, (28) insertion
of pectoral fin……………………………….…………………………………………21-22
Figure 3. Between-group PCA on partial warps of 501 Lake Trout. (a) First and second principal
components for head shape representing 56.2% and 15.9% of the variance respectively
distinguishing the four ecotypes. (b) First and second principal components for body shape
representing 65.5% and 14.5% of the variance respectively that distinguish leans from
siscowets based mainly on belly curvature. (c) Third and fourth principal component for
head shape representing 11.5% and 7.2% of the variance respectively distinguishing the
four sites. (d) Third and fourth principal components for body shape representing 8.8% and
3.0% of the variance respectively distinguishing the four sites. The colored points refer to
the mean scores for each ecotype in each site. The sites are: Big Reef (black), Isle Royale
(blue), Stannard Rock (red) and Superior Shoals (green). Ecotypes are: Siscowet (FT),
Humper (HT), Lean (LT) and Redfin (RF). Under each ecotype are drawn the consensus
shapes of all four ecotypes (gray) with the outline of the ecotype in question (black). The
shaded ellipses have been drawn for clarity. ……………………………………………..32
Figure 4. Population structure analysis of Lake Trout; a) Admixture plot based on 486 individuals
and 6822 SNPs (including outliers) for different values of K. Individuals are shown by sites
and ecotypes. b) Neighbour joining tree based on 486 individuals and 6822 SNPs including
outliers. Yellow circles represent Big Reef, orange circles Stannard Rock, blue circles Isle
Royale and green circles Superior Shoals. Bootstrapping support is indicated on each
branch. The four ecotypes are represented for each site; Lean (LT), Humper (HT), Redfin
(RF) and Siscowet (FT).…………………………………………………..………………38
Figure 5. Assignment success of individuals to their sampling sites (a) or ecotypes (b). Percentage
assignment is written below circles with the exact number of individuals assigned within
brackets. Percentage of correct assignment to either sampling sites or ecotypes is in bold.
viiiSites are: Big Reef (BR), Isle Royale (IR), Stannard Rock (SR), Superior Shoals (SS).
Ecotypes are: Humper (HT), Siscowet (FT), Lean (LT) and Redfin (RF).…………..….39
Figure 6. Venn diagrams of outliers detected by LFMM and BAYESCAN among sites, ecotypes or
among ecotypes within sites a) Outliers detected among the four sites by BAYESCAN and
LFMM including outliers detected by LFMM using morphological PC scores. b) Outliers
detected among the four pooled ecotypes by BAYESCAN and LFMM including outliers
detected by LFMM using morphological PC scores. c) Outliers among ecotypes within
each site detected by LFMM…………………………………...………………….…….41
ixRemerciements
Un goût certain pour tout ce qui a trait à la nature a toujours été présent en moi, ainsi qu’un
intérêt particulier pour la vie aquatique sous toutes ses formes. Par conséquent, les métiers
en relation avec la vie sauvage m’ont toujours attirée et c'est pourquoi j’ai choisi d’étudier
dans le domaine de la biologie. C’est également la raison principale de ma venue au
Laboratoire du Dr. Louis Bernatchez, titulaire de la chaire de recherche du Canada en
génomique et conservation de ressources aquatiques.
J’aimerais le remercier pour m’avoir donné la chance de poursuivre ma passion dans son
laboratoire, de même que tous les étudiants, stagiaires et professionnels de recherches
présents, sans qui ce projet n’aurait pu être mené à terme. Plus précisément, j’aimerais
mentionner, pour leur précieuse collaboration, Cécilia Hernandez pour la partie « laboratoire
» du projet, mes collègues de bureau, dont Simon Bernatchez, ainsi que Charles Perrier,
Martin Laporte, Jean-Sébastien Moore, Anne-Laure Ferchand, Clément Rougeux, Laura
Benestan, Thierry Gosselin et Éric Normandeau pour leurs aides et conseils quant à
l’analyse et l’interprétation des résultats.
Ce projet n’aurait pu prendre vie sans le concours et le soutien de la Commission des
Pêcheries des Grands Lacs, et tout particulièrement, sans celui de monsieur Andrew Muir.
Toute ma reconnaissance à C. Krueger, C. Bronte, M. Hansen et tous les membres de
l’équipage du Kiyi pour leur aide lors de l’échantillonnage sur le lac Supérieur. Et c’est sans
oublier la collaboration de monsieur Frederick Goetz, de l'Agence américaine d'observation
océanique et atmosphérique sans qui l’étude du site « Isle Royale » n’aurait pas été possible.
Finalement, j’aimerais exprimer ma gratitude à ma mère et mon conjoint qui, malgré bien
des embûches, ont toujours cru en moi et m’ont encouragée dans cette aventure.
Sur ce, je vous souhaite une bonne lecture.
xAvant-propos
Vous trouverez, au cœur de ce mémoire, un article intitulé « Investigating the Extent of
Parallelism in Morphological and Genomic Divergence among Lake Trout Ecotypes in Lake
Superior ». Cet article a été accepté avec corrections mineures par la revue « Molecular
Ecology ».
Les co-auteurs sont messieurs Andrew Muir, Frederick Goetz, Charles Perrier et Éric
Normandeau, mon co-directeur Pascal Sirois et mon directeur monsieur Louis Bernatchez.
Ont effectué, en grande majorité, le travail de terrain, messieurs Andrew Muir et Frederick
Goetz. Ont participé aux analyses, messieurs Éric Normandeau et Charles Perrier, ce dernier
ayant aussi travaillé à l’interprétation des résultats. Pour ma part, j’ai réalisé le travail en
laboratoire, rédigé l’article, effectué les analyses et interpréter les résultats. De plus, tous les
co-auteurs ont collaboré à la révision et l’amélioration du manuscrit.
Ce projet a été rendu possible grâce à la Commission des Pêcheries des Grands Lacs
(GLFC) ainsi qu’au regroupement Ressources Aquatiques Québec (RAQ).
xiIntroduction
L’apparition de nouvelles espèces
La biodiversité, telle qu’on la connaît aujourd’hui, est le fruit de plusieurs processus
évolutifs agissant de concert ou de façon individuelle sur des individus ou des populations.
La nature de ces forces soit la mutation, la dérive génétique, la migration, la sélection ainsi
que leurs modes d’action, a toujours été au centre de l’étude de l’évolution. On reconnaît
maintenant leur importance dans plusieurs autres domaines tels que la gestion et la
conservation des ressources. Le processus de spéciation est connu comme étant un moteur
de diversité par lequel une espèce se divise en deux ou plusieurs espèces distinctes (Hendry
2009; Weissing et coll. 2011). L’apparition de ces nouvelles espèces est causée par plusieurs
forces agissant comme barrières aux flux de gènes entre ces taxons naissants (Turelli et coll.
2001; Hendry 2009). Ces forces prennent la forme d’isolements reproducteurs agissant à
plusieurs étapes du cycle de vie, soit avant la reproduction (pré-zygotique) ou directement
sur les hybrides produits (post-zygotique). Les barrières pré-zygotiques peuvent être de
types écologiques, comportementales, mécaniques ou gamétiques (Schluter 2001; Turelli et
coll. 2001) tandis que celles post-zygotiques découlent de la stérilité ou la mortalité des
hybrides, ou de leur valeur sélective plus faible en comparaison des parents (Butlin et coll.
2012; Turelli et coll. 2001).
La spéciation est le plus souvent considérée comme un processus graduel où l’accumulation
de différences génétiques amène un isolement reproductif (Hendry et coll. 2009). C’est
pourquoi, la spéciation est représentée comme un continuum, où à l’une des extrémités, on
retrouve des populations présentant des variations adaptatives continues sans aucun
isolement reproductif et, à l’autre extrémité, des populations présentant des différences
adaptatives accompagnées par un isolement reproductif irréversible (Hendry et coll. 2009).
On retrouve donc des populations à différents stades, dits intermédiaires, de ce continuum.
1La compréhension des facteurs menant à un isolement reproductif et, plus spécifiquement à
son niveau d’achèvement, est primordial tant au niveau évolutif qu’au niveau de la
conservation (Seehausen et coll. 2008). Par exemple, le niveau d’achèvement de l’isolement
reproductif peut déterminer si, lorsque remise en contact, deux populations ou espèces vont
fusionner en une seule et même espèce et ainsi perdre les adaptations acquises (Seehausen et
coll. 2008).
On retrouve traditionnellement trois modes de spéciation selon le contexte spatial :
l’allopatrie, la parapatrie et la sympatrie. L’utilisation et la véracité de ce classement a
souvent fait l’objet de débats et de remises en question, mais est encore en vigueur
aujourd’hui (Butlin et coll. 2008; Fitzpatrick et coll. 2009). La forme la plus acceptée est
celle dite allopatrique, où les populations ou espèces sont séparées par des barrières
géographiques empêchant ou réduisant de manière significative la dispersion et, par
conséquent, l’échange de gènes (Mallet et coll. 2009). Ce faisant, les populations
accumulent des différences génétiques par dérive, ou par des pressions de sélection
différentes selon les habitats occupés (Turelli et coll. 2001). Lors d’un contact secondaire,
l’accumulation de ces différences pourra mener à un isolement reproductif empêchant
complètement ou diminuant le flux génique. Pour sa part, la spéciation dite en parapatrie
inclut un certain niveau d’échange de gènes entre les populations seulement à des zones de
contacts ou d’hybridation (Butlin et coll. 2012). À l’autre extrémité, on retrouve la
spéciation dite en sympatrie où les populations occupent une même région géographique et
où tous les individus ont la capacité de se rencontrer physiquement plus ou moins
fréquemment (Mallet et coll. 2009). Ce dernier mode de spéciation est causé par la présence
d’une sélection divergente liée aux facteurs biotiques et abiotiques présents (Turelli et coll.
2001). Ces facteurs pourraient prendre la forme d’une divergence écologique associée à
l’utilisation des ressources et de l’habitat (Blackie et coll. 2003; Hendry 2009).
La présence d’une sélection divergente sur des traits entre populations utilisant différentes
ressources et menant à l’isolement reproductif est souvent appelée spéciation écologique
(Hendry 2009; Schluter 2016; Gavrilets et coll. 2007). Cette forme de spéciation cause
l’apparition d’un certain isolement pré-zygotique sous la forme de sélection sexuelle ou de
2ségrégation temporelle, et post-zygotique lié à la sélection contre les formes intermédiaires
quant à l’exploitation des ressources et à leur compétition (Hendry 2009).
La présence d’un ou de plusieurs phénotypes distincts dans une population utilisant
différentes ressources est appelée le polymorphisme des ressources (Wimberger 1994;
Skúlason et Smith 1995). Ce polymorphisme peut prendre la forme de différences
morphologiques marquées, de différences comportementales ou de traits d’histoire de vie
(Skúlason et Smith 1995). On retrouve souvent le polymorphisme des ressources dans un
contexte de niche inoccupée et de diminution dans la compétition interspécifique (Skúlason
et Smith 1995). Ces conditions, présentes dans les lacs lors du retrait des glaciers, ont été
proposées comme moteur dans la radiation adaptative observée chez les poissons d’eau
douce en Amérique du Nord. Cette radiation se traduit par la différentiation d’une espèce
ancestrale en une série d’espèces adaptées à différentes niches écologiques (Schluter 2001).
Un des exemples les plus connus est l’évolution répétée des formes limnétiques et
benthiques retrouvées en sympatrie chez plusieurs espèces de poissons telles que le corégone
(Coregonus), l’épinoche à trois épines (Gasterosteus aculeatus), et le crapet-soleil (Lepomis
gibbosus) et le crapet arlequin (Lepomis macrochirus) (Jonsson 2001; Wimberger 1994;
Skùlason et Smith 1995).
Parallélisme dans la divergence phénotypique
Le parallélisme phénotypique, soit l’apparition répétée de phénotypes similaires adaptés à
certaines conditions environnementales, a été observé chez plusieurs vertébrés (Elmer et
Meyer 2011; Arendt et Reznick 2008). Les outils génomiques modernes permettent
maintenant de vérifier si ce parallélisme au niveau phénotypique est suivi d’une base
génétique parallèle sous-jacente. Ce parallélisme génétique peut découler du polymorphisme
génétique ancestral présent chez l’espèce ou prendre la forme de nouvelles mutations (Elmer
et Meyer 2011). Le processus d’adaptation à partir du polymorphisme génétique ancestral
est considéré comme plus probable et plus rapide que la mutation pour trois raisons.
Premièrement, les allèles sont déjà disponibles, deuxièmement ils sont présents à une plus
3haute fréquence dans la population, et troisièmement, étant déjà présents, les allèles ont été
testés par la sélection et dès lors sont compatibles avec le reste du génome (Barrett et
Schluter 2008; Elmer et Meyer 2011). Plusieurs exemples de parallélisme phénotypiques
associés à une base génétique similaire ont été documentés chez la souris (Peromyscus
polionotus), l’épinoche (Gasterosteus aculeatus), le corégone (Coregonus), les cichlidés et
le tétra aveugle (Astyanax mexicanus). Par contre, pour les mêmes espèces énumérées
précédemment, une ou plusieurs populations affichant le même phénotype ne montraient pas
de parallélisme au niveau génétique et, par conséquent, le phénotype exprimé serait causé
par un changement moléculaire différent au courant du développement (Elmer et Meyer
2011; Arendt et Reznick 2008). On peut donc observer au sein d’une même population
différente routes évolutives menant à un même phénotype.
Le touladi
En Amérique du Nord
Le touladi est un salmonidé répandu en Amérique du Nord dont la distribution suit les
limites de la calotte glacière du Wisconsin, où suivant le retrait des glaciers, il a colonisé les
plans d’eau de toutes tailles (Muir et coll. 2015; Wilson et Hebert 1996; Eshenroder 2008).
Les populations présentes en Amérique du Nord, proviennent de plusieurs refuges glaciaires,
soit ceux de l’Atlantique, du Béringien, du Mississippi, du Montana et du Nahanni (Wilson
et Hebert 1998). Ce poisson d’eau douce est reconnu pour sa diversité, comparable à celle de
l’Omble chevalier, tant au niveau de la morphologie, des traits d’histoire de vie que de la
physiologie et de l’écologie (Muir et coll. 2015). L’apparition répétée de formes adaptées à
l’eau peu profonde ou ayant des adaptations liées à un mode de vie en eau profonde a été
répertoriée dans plusieurs lacs nord-américains (Muir et coll. 2015). Par exemple, on
retrouve deux écotypes dans le lac Mistassini et le lac Rush, dont l’un adapté à la vie en eau
peu profonde et l’autre aux eaux profondes (Zimmerman et coll. 2007; Muir et coll. 2015). Il
est à noter que dans des lacs de plus grande superficie, on peut retrouver jusqu’à quatre
4écotypes différents. Par exemple, jusqu’à quatre écotypes adaptés aux eaux peu profondes
ont été trouvé au Grand lac de l’Ours (Harris et coll. 2014; Muir et coll. 2015), tandis qu’au
Grand lac des Esclaves, on retrouve un écotype adapté aux eaux peu profondes ainsi que
deux adaptés aux eaux profondes (Zimmerman et coll. 2006; Muir et coll. 2015).
Dans le lac Supérieur
Dans le lac Supérieur, on retrouve quatre écotypes de touladi : le « lean » dit maigre, le «
siscowet » dit gras, le « humper » dit bossu et le « redfin » (Muir et coll. 2014). Ces
écotypes diffèrent principalement au niveau de l’habitat, de l’alimentation, de la
morphologie et dans certains traits d’histoire de vie, tels que la croissance et la taille à la
maturité (Muir et coll. 2015). Le maigre est le seul écotype exploité par la pêche commercial
et récréative en raison du faible taux de lipides présent dans ses tissus (Bronte et Sitar 2008).
Cet écotype est décrit comme ayant une forme allongée, un museau droit et pointu ainsi
qu’un long pédoncule caudal lui permettant de nager de manière prolongée (Burnham-Curtis
& Smith 1994; Moore & Bronte 2001; Zimmerman et coll. 2006). On le retrouve en eaux
peu profondes (< 100 m) près des côtes où il se nourrit de petites espèces de poissons
pélagiques ou de fond tels que l’éperlan arc-en-ciel (Osmerus mordax), le cisco de lac
(Coregonus artedi) et le chabot visqueux (Cottus cognatus) (Goetz et coll. 2011; Ray et coll.
2007; Zimmerman et coll. 2006). À l’opposé, l’écotype gras est le plus abondant et on le
retrouve principalement en eaux profondes (> 100 m) au large des côtes (Bronte et coll.
2003; Ray et coll. 2007; Goetz et coll. 2011). Cet écotype est décrit comme ayant un corps
robuste et épais, une tête plutôt petite, un museau court et incliné ainsi que de grands yeux. Il
est aussi reconnu pour avoir un pédoncule caudal épais et long ainsi que de grandes
nageoires lui permettant d’effectuer de soudaines poussées de vitesse pour attraper ses
proies. De plus, l’écotype gras tient son nom du niveau élevé de lipides dans ses viscères lui
donnant une flottabilité plutôt neutre, et par le fait même, facilitant les migrations verticales
dans la colonne d’eau à la poursuite de ses proies. Celles-ci sont composées principalement
de corégoninés (corégones, ciscos, ménominis), de chabots de profondeur (Myoxocephalus
thompsoni) et de lottes (Lota lota) (Goetz et coll. 2011; Ahrenstorff et coll. 2011; Hrabik et
5coll. 2014; Bronte et coll. 2003; Bronte and Sitar 2008; Burnham-Curtis and Smith 1994;
Hansen et coll. 2012). Habitant les hauts-fonds au large des côtes, l’écotype bossu est peu
répandu dans le lac Supérieur (Zimmerman et coll. 2006). On le reconnaît à sa petite taille, à
sa petite tête ayant un museau incliné et de grands yeux, à une paroi abdominale plutôt
mince ainsi qu’à un niveau de lipides intermédiaire (Bronte et coll. 2003; Goetz et coll.
2011; Moore et Bronte 2001). C’est le seul écotype qui ne soit pas piscivores à l’âge adulte,
se nourrissant principalement de Mysis diluviana (Stafford et coll. 2014). Finalement,
l’écotype « redfin » a été décrit récemment près de l’Isle Royale dans la partie ouest du lac
Supérieur (Muir et coll. 2014). Il est décrit comme ayant un corps robuste, une tête, un
museau et des yeux larges, un pédoncule caudal long et épais ainsi que de grandes nageoires
(Muir et coll. 2014). On le retrouve en eaux de profondeur moyenne (~ 80 m) et il semble
avoir un niveau de lipides semblable et même plus élevé que l’écotype gras (Muir et coll.
2014; Hansen et coll. 2016). De plus, il est de plus grande taille, plus lourd, plus longévif et
a une croissance plus lente que les autres écotypes à l’Isle Royale (Hansen et coll. 2016).
L’origine et le maintien des écotypes
Plusieurs hypothèses ont été avancées pour expliquer la présence de ces écotypes dans le lac
Supérieur (Wilson et Mandrak 2004; Eshenroder 2008). La première est la plasticité
phénotypique induite par l’environnement qui se traduit par l’apparition de plusieurs
phénotypes à partir d’un seul génotype selon des variations biotiques ou abiotiques. La
deuxième hypothèse sous-entend la présence d’une base génétique entre les différents
écotypes, et donc d’un certain isolement reproductif (Goetz et coll. 2010). Plusieurs
évidences pointent vers la deuxième hypothèse, soit la présence d’une base génétique, pour
expliquer l’apparition et le maintien des écotypes. En premier lieu, les écotypes (maigre,
bossu, et gras) diffèrent au niveau de certains os crâniens, plus spécifiquement au niveau de
la forme de l’opercule, du supraethmoïde ou du dermethmoïde. Des variations
environnementales ou ontogéniques sont peu plausibles pour expliquer les différences
morphologiques observées avec ces os (Burnham-Curtis et Smith 1994). De plus, la
progéniture des écotypes maigres et gras a été élevée en milieu contrôlé et retenait la plupart
des traits distinctifs les séparant (Goetz et coll. 2010). La même étude a aussi décelé des
6différences au niveau de l’expression de certains gènes, entre ces écotypes, associés au
métabolisme des lipides. Aussi, deux études utilisant des microsatellites ont montré, bien
que faiblement, une plus grande variance liée aux écotypes qu’aux localités (Page et coll.
2004; Guinand et coll. 2012). Par contre, plusieurs études ont démontré un schéma contraire,
soit une plus grande variance liée aux sites d’échantillonnage qu’aux écotypes (Dehring et
coll. 1981; Ihssen et coll. 1988; Baillie et coll. 2016). L’homogénéisation génétique et
phénotypique des écotypes de touladi, vivant dans le lac Supérieur, a été avancée pour
expliquer la présence d’une plus grande différentiation génétique entre les sites
d’échantillonnage qu’entre les écotypes (Baillie et coll. 2016). En effet, en étudiant des
échantillons préeffondrements et contemporains, Baillie et collaborateurs (2016) ont
identifié un regroupement génétique par écotypes plus importants chez les individus
préeffondrements que chez les individus contemporains qui se regroupent majoritairement
par site d’échantillonnage. La surpêche, l’ensemencement intensif et l’introduction
d’espèces invasives ont été énoncés pour expliquer le plus grand échange de gènes, observé
aujourd’hui, entre les différents écotypes (Baillie et coll. 2016).
Problématique
Bouleversement du système aquatique
Jusqu’à dix variétés de touladis ont été décrites de manière anecdotique par les pêcheurs du
lac Supérieur, selon des distinctions au niveau de l’apparence physique, de l’habitat et du
comportement. Celles-ci étaient exploitées commercialement, tout particulièrement la forme
maigre dû à son faible taux en lipides (Goodier 1981; Bronte et Sitar 2008). Dans les années
1960, la pêche s’est effondrée et le touladi a disparu des Grands Lacs à l’exception du lac
Supérieur et d’une partie du lac Huron (Bronte et Sitar 2008). Cet effondrement a été causé
principalement par l’activité humaine, dont la surpêche, la dégradation de l’habitat et
l’introduction d’espèces envahissantes notamment la lamproie marine (Bronte et Sitar 2008;
Page et coll. 2003; Page et coll. 2004). En ce qui a trait aux salmonidés, six espèces ont été
introduites dont la truite brune et arc-en-ciel, ainsi que les saumons chinook, coho, rose et
7atlantique. Elles ont été introduites afin de diversifier et d’augmenter la pêche sportive, mais
ont eu pour effet de changer la dynamique des poissons-fourrages ainsi que de leurs
prédateurs (Bronte et coll. 2003). Le contrôle de la lamproie marine et l’ensemencement
intensif ont permis de restaurer les stocks de touladis dans le lac Supérieur, mais bien en
deçà des niveaux historiques (Bronte & Sitar 2008). Malgré plusieurs efforts de restauration
dans les autres Grands Lacs, aucune population reproductrice autonome n’a pu être créée
dans ces derniers (Page et coll. 2003).
Plan de réintroduction des poissons de fonds
Le rétablissement des poissons de fonds tels que le touladi dans les Grands Lacs est encore
incertain et a fait l’objet d’un article par deux biologistes de la Commission des Pêcheries
des Grands Lacs (Zimmerman et Krueger 2009). Ceux-ci ont examiné plusieurs facteurs
pouvant freiner ou empêcher son rétablissement dans les Grands Lacs. À cet effet, quatre
sujets de recherche ont été désignés comme capitaux, soient :
− une plus grande compréhension des variations spatiales et temporelles liées au
recrutement,
− l’identification de la ou des sources de mortalité des premiers stades de vie,
− la comparaison de l’histoire de vie et de l’écologie de l’écotype maigre avec ceux
des autres écotypes, et
− la détermination du niveau de différentiation génétique entre les écotypes et les
populations.
La diversité génétique est associée au potentiel évolutif d’une espèce face aux changements
(Toro et Caballero 2005). C’est ainsi que le maintien d’une grande diversité génétique, ainsi
qu’une grande diversité phénotypique, augmenterait la résilience et la résistance des espèces
face aux changements dans l’environnement. Donc, l’établissement d’une combinaison de
populations adaptées à divers habitats devrait augmenter la résilience du touladi dans un
même plan d’eau (Zimmerman et Krueger 2009).
L’utilisation d’outils génomiques de pointe ainsi que des approches pangénomiques
permettrait d’augmenter la résolution génétique entre les différents écotypes, et ce faisant de
8préciser les mécanismes soutenant l’origine et le maintien des écotypes de touladi dans le
Lac Supérieur. De plus, un échantillonnage à plusieurs sites connus pour l’abondance de
cette espèce permettrait de déterminer la connectivité génétique à travers le lac, mais aussi
de vérifier la présence de variabilité génétique liée à la localité.
Objectifs
L’objectif principal de cette étude était d’approfondir les connaissances quant à l’origine et
l’ampleur de la différentiation génétique des différents écotypes de touladi dans une optique
de gestion et de conservation. Afin d’atteindre cet objectif, deux cibles spécifiques ont été
identifiés soit : (i) de documenter et comparer l’étendue de la structure et de la connectivité
génétique entre les différents écotypes et entre les différents sites d’échantillonnage et (ii)
d’identifier de possibles divergences adaptatives incluant les associations génotype-
phénotype entre les différents écotypes.
9Chapter I: Investigating the Extent of Parallelism in
Morphological and Genomic Divergence among
Lake Trout Ecotypes in Lake Superior
Chapitre I: Évaluation de l’étendue du parallélisme dans la divergence morphologique
et génomique des écotypes de touladi du lac Supérieur
10Résumé
Comprendre le processus de spéciation écologique par lequel de nouvelles espèces émergent
est au centre de la biologie évolutive, mais il est tout aussi important en ce qui a trait à la
conservation et la gestion. Le touladi (Salvelinus namaycush) est reconnu pour la présence
d’écotypes associés à l’utilisation de l’habitat et des ressources à travers l’Amérique du
Nord. Nous avons utilisé le séquençage de nouvelle génération afin de définir la structure
génétique fine des quatre écotypes de touladis décrits dans le lac Supérieur, le plus grand lac
en Amérique du Nord. Pour ce faire, 486 individus provenant des quatre écotypes présents
en sympatrie à quatre sites d’échantillonnage ont été génotypés à 6822 SNPs en utilisant la
technologie de séquençage « RAD ». De plus, les traits phénotypiques de tous les individus
séquencés ont été documentés sous la forme d’analyses morphométriques conjointes de la
forme de la tête et du corps ainsi qu’une identification visuelle. Les résultats obtenus ont
révélé la présence de différents niveaux de différentiation morphologique et génétique à
l’intérieur des différents sites. De manière générale, la différentiation génétique était faible,
mais significative et était en moyenne près de trois fois plus prononcées entre les sites (FST
Moyen = 0.016 [0.012; 0.021]) qu’entre les écotypes à chacun de ces sites (FST Moyen=
0.005 [0.004; 0.007]). Ceci indique un flux de gènes plus important ou un ancêtre commun
plus récent à l’intérieur de chaque site, qu’entre les populations du même écotype. Malgré,
le peu d’indication quant à la présence d’une origine commune des différentes populations
appartenant à chaque écotype, des individus correspondant à la description des différents
écotypes ont été identifiés à chacun des sites, ce qui suggère une convergence dans les traits
phénotypiques. Des indices de la présence de sélection divergente ont été trouvés entre les
écotypes ou en association avec des variations morphologiques. En effet, plusieurs
marqueurs associés au métabolisme des lipides ainsi qu’à l’acuité visuelle sont
particulièrement intéressants dans un contexte de divergence entre les écotypes.
Globalement, la présence de différents niveaux de différentiation génomique entre les
écotypes à chacun des sites ainsi que la présence de loci différentiés associés à des fonctions
11biologiques d’intérêt supportent un scénario de divergence répété des différents écotypes en
sympatrie, à l’intérieur de chaque localité.
12Abstract
Understanding the emergence of species through the process of ecological speciation is a
central question in evolutionary biology which also has implications for conservation and
management. Lake Trout (Salvelinus namaycush) is renowned for the occurrence of
different ecotypes linked to resource and habitat use throughout North America. We used
next generation sequencing to unravel the fine genetic structure of the four Lake Trout
ecotypes described in Lake Superior. A total of 486 individuals from four sites where the
four ecotypes occur in sympatry were genotyped at 6822 filtered SNPs using RADseq
technology. Phenotypic traits of all sequenced fish were documented using a combination of
morphometric analyses. Our results revealed different extent of morphological and genetic
differentiation within the different sites. Overall, genetic differentiation was weak but
significant and was on average three times higher between sites (Mean FST = 0.016) than
between ecotypes within sites (Mean FST = 0.005) indicating higher level of gene flow
and/or a more recent shared ancestor between ecotypes within each site than between
populations of the same ecotype. Evidence of divergent selection was also found between
ecotypes and/or in association with morphological variation. Outlier loci found in gene
related to lipid metabolism and visual acuity were of particular interest in this context of
ecotypes divergence. Overall, the occurrence of different levels of both genomic and
phenotypic differentiation between ecotypes within each site with several differentiated loci
linked to relevant biological functions support the presence of a continuum of divergence in
Lake Trout.
13Introduction
The study of diversification and ultimately speciation is central to evolution and relevant for
conservation biology (Weissing et al. 2011). The most common and established mechanism
of speciation is divergence in allopatry, where spatial and geographical barrier prevent gene
flow, thus allowing genetic incompatibilities to accumulate, subsequently resulting in
reproductive isolation following secondary contact (Coyne and Orr 2004; Tittes and Kane
2014). Some examples of allopatric isolation mechanisms in fishes include the glacial cycles
in North America responsible for the origin of many freshwater species (April et al. 2013),
the rise and fall of Lake Tanganyika, and the barriers created by high water flow in large
rivers such as the Amazon or Congo River (reviewed in Bernardi 2013). However, a
geographic barrier is not always needed and speciation can emerge in sympatry, or in
parapatry despite high gene flow, by divergent selection on ecologically important traits
(Tittes and Kane 2014; Gavrilets et al. 2007). Divergent selection on ecological traits can be
caused by biotic and abiotic influences where adaptations to different environment or
ecological niches result in the emergence of reproductive incompatibilities (Bernardi 2013;
Nosil et al. 2009). The latter may create a continuum of divergence from continuous
variation within a single gene pool, to ecotype formation and finally to complete
differentiation and reproductive isolation (Lu and Bernatchez 1999; Nosil et al. 2009;
Hendry 2009). Models and case studies have shown that sympatric speciation is possible
under gene flow when few loci underlying the divergent trait undergo strong selection,
whereas gene flow homogenises the rest of the genome (Gavrilets et al. 2007; Franchini et
al. 2013).
Ecological speciation has been extensively documented in several geologically young fish
species living in sympatry. For instance sympatric speciation has occurred in Midas cichlids
(Amphilophus spp.) (Franchini et al. 2013), Lake Victoria cichlids (Wagner et al. 2013) but
more commonly in several temperate freshwater fishes such as stickleback (Gasterosteus
spp.), smelt (Osmerus spp.) and mainly in salmonids such as whitefish (Coregonus spp.),
trout (Salmo spp.), Pacific salmon (Oncorhynchus spp.) and charrs (Salvelinus spp.) (Taylor
1999; Jonsson and Jonsson 2001). Sympatric speciation is usually linked to trophic
14Vous pouvez aussi lire

















































