L'engraissement spontané du foie chez les palmipèdes : état des lieux et perspectives de recherche - INRA ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
L’engraissement spontané INRA Prod. Anim.,
2018, 31 (2), 117-130
du foie chez les palmipèdes :
état des lieux et perspectives de recherche
Christelle KNUDSEN1, Cécile BONNEFONT1, Laurence FORTUN-LAMOTHE1, Karine RICAUD2, Xavier FERNANDEZ1
1
GenPhySE, Université de Toulouse, INRA, ENVT, 31326, Castanet Tolosan, France
2
INRA, Univ Pau & Pays Adour, E2S UPPA, UMR 1419, Nutrition, Métabolisme, Aquaculture, 64310, Saint Pée sur Nivelle,
France
Courriel : christelle.knudsen@inra.fr
Aujourd’hui, la production de foie gras repose sur le gavage des animaux. Dans un contexte d’intensification
du questionnement sociétal relatif aux impacts du gavage sur le bien-être animal, questionnement relayé par
les associations de protection animale ainsi que par le Conseil de l’Europe, l’INRA a entrepris depuis 2009 des
recherches sur des méthodes alternatives à cette pratique pour la production de foie engraissé.1
Introduction L’embucage, acte d’introduction animal est interdite, sauf pour rai-
de l’embuc de gavage dans le jabot son de santé. Outre cette mesure, le
(canards) ou pseudo-jabot (oies), et Comité Permanent de la Convention
Chez les oiseaux, dont les palmi- le caractère forcé de l’ingestion sont Européenne pour la protection des
pèdes, le foie constitue un organe aujourd’hui remis en question par les animaux dans les élevages a recom-
de synthèse et de stockage lipidique associations de protection animale, mandé que des études portant sur
transitoire, utilisé en phase pré-migra- mais aussi par le conseil de l’Europe. des méthodes alternatives à la prise
toire notamment (Odum, 1960 ; Pond, Depuis 1999 le conseil de l’Europe forcée d’aliment chez les palmipèdes
1978). Cette aptitude est exploitée recommande ainsi que la produc- soient mises en place dans les pays
chez les palmipèdes dans le cadre de la tion de foie gras ne soit exercée dans européens producteurs de foie gras, à
production de foie gras lors du gavage. aucun pays européen n’étant pas déjà savoir la France, la Hongrie, la Bulgarie,
Selon la définition de Labie et Tournut producteur à cette date (Article 24 du l’Espagne et la Belgique (Articles 24
(1970), le foie gras des palmipèdes texte relatif aux canards de Barbarie et et 25). Ces recommandations, ainsi que
gavés est l’expression d’une stéatose les hybrides de canards de Barbarie et les controverses autour de la filière foie
hépatique, autrement dit un engrais- de canards domestiques et Article 25 gras, ont entraîné le déploiement de
sement du foie, d’origine nutrition- du texte relatif aux oies domestiques produits alternatifs au foie gras à base
nelle, hypertrophique et réversible. et à leurs croisements du 22 juin 1999). de matières premières végétales ou
Ce mets est considéré depuis 2006 Cette recommandation n’a à ce jour animales. Les produits issus de matières
comme faisant partie du patrimoine été retranscrite spécifiquement pour premières animales peuvent se distin-
culturel et gastronomique protégé les palmipèdes que dans la législa- guer en deux catégories : les prépara-
en France (Article L.654-27-1 du code tion Italienne en 2001 et l’était déjà tions à base de foies non engraissés
rural) et est générateur de 2 milliards dans la législation polonaise depuis supplémentées en graisse animale
d’euros de chiffre d’affaires, 30 000 1997, mais les législations de plusieurs ou végétale et les foies engraissés
emplois directs et 100 000 indirects en pays Européens tels que l’Allemagne, spontanément, sans alimentation for-
France en 2014 (estimations du Comité le Danemark, la Suède, le Royaume cée. Quelques éleveurs en France, en
Interprofessionnel des Palmipèdes à Uni et les Pays Bas stipulent d’ores et Espagne et aux États Unis notamment
Foie Gras – CIFOG). déjà que l’alimentation forcée d’un ont ainsi témoigné de la possibilité
1
Cet article est adapté de la synthèse présentée aux Journées de la Recherche Avicole 2017 (Knudsen et al., 2017b).
https://doi.org/10.20870/productions-animales.2018.31.2.2318 INRA Productions Animales, 2018, numéro 2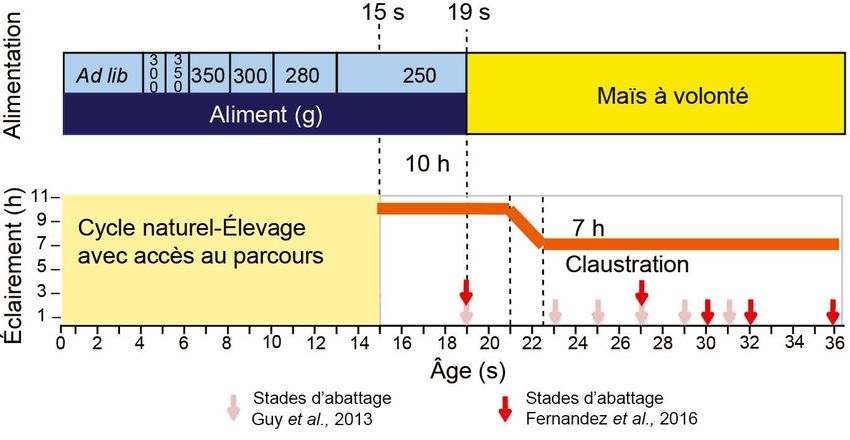
118 / Christelle knudsen et al.
’induire une stéatose hépatique spon-
d ermettant un faible captage des lipides
p une forte mobilisation d’énergie, comme
tanée chez les palmipèdes, mais aucune par cet organe. La lipogenèse de novo se la reproduction ou la migration. Chez les
donnée bibliographique chiffrée n’est fait à partir de glucides alimentaires qui oiseaux le stockage des lipides s’effec-
disponible sur ces élevages. sont dégradés en glucose dans la partie tue ainsi principalement au niveau des
supérieure du tube digestif, puis absor- adipocytes et des myocytes, mais peut
La présente synthèse a pour objectif bés à travers la paroi de l’intestin grêle également se faire de façon transitoire
de définir l’état d’avancement des tra- et transporté jusqu’au foie. Le glucose y au niveau des hépatocytes.
vaux menés sur l’engraissement hépa- est alors métabolisé en acides gras puis
tique des palmipèdes sans alimentation en triglycérides. Le foie constitue le site 1.2. Comportement
forcée et d’en présenter les perspec- principal, et quasi exclusif, de la synthèse alimentaire et métabolisme
tives de recherche. Nous nous focalise- endogène de lipides chez les oiseaux, hépatique chez les oiseaux
rons ainsi sur les différentes recherches alors que chez les mammifères, elle s’ef- migrateurs
académiques menées sur ce sujet fectue principalement au niveau des tis-
et les perspectives de recherches en sus périphériques, adipeux notamment Afin de répondre aux besoins énergé-
ayant découlé. Nous parlerons de foies (O’Hea et Leveille, 1969 ; Evans, 1972 ; tiques importants lors de leur voyage,
« engraissés » pour se référer au produit Leveille et al., 1975). Ainsi, bien que les oiseaux migrateurs développent
de systèmes sans gavage et non de « foie les sites de synthèse varient, chez les un engraissement important durant
gras » puisque, par l’absence de l’acte de oiseaux, comme chez les mammifères, la phase pré-migratoire. Cet engrais
gavage, le produit ne répond pas à sa les précurseurs glucidiques alimentaires sement est généralement dépendant
définition réglementaire actuelle (Article sont assemblés principalement en trig- de la distance à parcourir, des arrêts
L.654-27-1 du code rural). lycérides, mais aussi en phospholipides potentiels sur le trajet et du gabarit de
et en cholestérol. Les lipides alimen- l’oiseau (King et Farner, 1965). Ainsi, chez
taires et endogènes peuvent ensuite certains oiseaux, comme la fauvette des
1. Bases biologiques être exportés vers les tissus adipeux et jardins (Sylvia borin) ou le colibri à gorge
de la stéatose hépatique les muscles ou stockés. L’exportation des rubis (Archilochus colubris), le poids vif
chez les oiseaux lipides via la circulation sanguine se fait peut être plus que doublé en l’espace de
sous forme de lipoprotéines en s’asso- quelques jours ou semaines, avec des
ciant à l’apolipoprotéine B pour les trig- réserves essentiellement sous forme de
La stéatose hépatique d’origine lycérides, formant les VLDL (« Very Low lipides (Odum et Connell, 1956 ; Odum,
nutritionnelle se définit comme l’ac- Density Lipoproteins ») et l’apoprotéine 1960 ; Pond, 1978 ; Bairlein et Gwinner,
cumulation de lipides sous forme de A-I pour les phospholipides et le cho- 1994). Cet engraissement semble prin-
triglycérides dans les cellules du foie, lestérol, formant les HDL (« High Density cipalement induit par une hyperpha-
les hépatocytes. Dans cette partie nous Lipoproteins ») (Hermier et al., 1988 ; gie, i.e. une consommation alimentaire
allons définir les voies métaboliques et Hermier, 1997). Plusieurs facteurs méta- accrue, mais aussi par une améliora-
les comportements alimentaires spéci- boliques, la choline notamment, parti- tion de l’efficacité digestive (Bairlein et
fiques aux oiseaux, migrateurs ou non, cipent à l’exportation des triglycérides. Gwinner, 1994), même si cette améliora-
induisant ce phénomène. Afin de permettre le captage des acides tion est variable selon les espèces (King
gras par les tissus périphériques, les et Farner, 1965). Il a ainsi été observé
1.1. Voies métaboliques VLDL circulantes sont ensuite hydroly- qu’en captivité plusieurs espèces
de synthèse, de transport sées via la lipoprotéine lipase à proximité d’échassiers migrateurs, ayant accès à
et de stockage des lipides des sites utilisateurs (principalement les une alimentation à volonté, pouvaient
chez les oiseaux muscles, les tissus adipeux et le foie). Les avoir une ingestion jusqu’à 8 fois supé-
acides gras libérés traversent les cellules rieure à celle nécessaire pour couvrir
Les lipides peuvent avoir deux ori- endothéliales et sont métabolisés en leur métabolisme basal, et dépassant
gines : alimentaire ou endogène. Les triglycérides, qui constituent la forme les niveaux nécessaires à la reproduc-
lipides alimentaires sont dégradés dans principale de stockage des lipides. tion ou au maintien en conditions de
l’intestin grêle. Chez les mammifères les froids extrêmes par exemple (Kvist et
lipides alimentaires sont transportés Dans des conditions métaboliques Lindstrom, 2003). Une modification du
sous forme de chylomicrons dans le sys- dites « basales » (hors croissance, repro- régime et des préférences alimentaires
tème lymphatique avant de rejoindre le duction…) il existe un équilibre entre la des oiseaux migrateurs, avec une inges-
système veineux, court-circuitant ainsi lipogenèse et l’exportation des lipides. tion d’aliments riches en glucides et en
le foie (McWilliams et al., 2004), alors Mais lorsque l’activité de lipogenèse lipides comme les fruits par exemple, a
que chez les oiseaux, les lipides simples dépasse la capacité d’exportation des également pu être observée en phase
sont absorbés à travers la paroi des lipides, à savoir la capacité de synthèse et de migration (Guglielmo, 2018). Durant
entérocytes et réassemblés avant d’être de sécrétion des lipoprotéines, un phé- cette période les niveaux d’ingestion
transportés par la veine porte jusqu’au nomène de stéatose hépatique, c’est-à- importants permettent aux oiseaux de
foie (Bensadoun et Rothfeld, 1972). Ces dire d’engraissement du foie, se produit. couvrir leurs besoins protéiques mal-
lipides passent donc par le foie avant Ce phénomène peut être observé dans gré des teneurs en protéines dans les
de rejoindre la circulation g énérale, des situations particulières nécessitant aliments faibles.
INRA Productions Animales, 2018, numéro 2L’engraissement spontané du foie chez les palmipèdes : état des lieux et perspectives de recherche / 119
Peu de données sont disponibles quant La modulation de la quantité et de la à certains stades de vie. Les animaux
à la répartition de la masse graisseuse, qui qualité nutritionnelle des aliments dis- domestiques ont fait l’objet d’un pro-
semble variable selon les espèces (Pond, ponibles semble également accélérer cessus de domestication et de sélec-
1978 ; Guglielmo, 2018). La majeure par- l’engraissement chez certaines espèces tion et sont élevés dans des conditions
tie de l’engraissement semble cependant comme la fauvette des jardins (Sylvia maitrisées par l’homme pour répondre
se faire au niveau sous-cutané, une pro- borin, Totzke et al., 2000 ; Cornelius à ses objectifs (production, sport, récréa-
portion minoritaire étant stockée dans le et al., 2013). Les facteurs environne- tion…). En conséquence, leur compor-
foie et les muscles. Le poids de foie aug- mentaux induisant le comportement tement peut être assez éloigné de celui
mente durant la migration, mais cette d’hyperphagie pré-migratoire semblent des animaux sauvages. C’est pourquoi,
prise de poids n’est pas toujours associée cependant être très différents entre le dans cette synthèse nous parlerons de
à un engraissement. Chez une espèce printemps et l’automne. En effet, les comportement « normal » des animaux,
de passereaux (Dumetella carolinensis) conditions environnementales varient au sens de « attendu pour cette espèce
par exemple le poids de foie augmente énormément entre le printemps et en l’absence de contraintes fortes »,
d’environ 25 % avant la migration d’au- l’automne, et les réponses physio- plutôt que de comportement naturel. Il
tomne alors que son taux de lipides reste logiques de migration des animaux s’agit par exemple, pour les palmipèdes,
constant (Corder et al., 2016). À l’inverse également (Agatsuma et Ramenofsky, de comportements tels que celui de se
chez la bernache du Canada (Branta 2006, Cornelius et al., 2013). Les oiseaux déplacer librement, de battre des ailes,
canadensis minima), un engraissement seraient ainsi plus sensibles à la pho- d’avoir des interactions sociales, etc. Afin
périphérique associé à un engraisse- topériode au printemps qu’en automne de répondre aux attentes sociétales rela-
ment hépatique a été observé durant (Bairlein et Gwinner, 1994). tives au gavage, les systèmes alternatifs
la migration de printemps (Raveling, développés pour la production d’un foie
1979). De même, chez le bruant à gorge Des facteurs hormonaux pourraient engraissé ne doivent pas avoir recours
blanche (Zonotrichia albicollis), une aug- également être des inducteurs du com- à l’alimentation forcée et doivent per-
mentation du taux de lipides hépatiques portement migratoire, mais peu de mettre aux animaux d’exprimer un com-
de 6 à 11 % a pu être observée durant la données concordantes sur ces leviers portement « normal ». Partant de ces
phase pré-migratoire (Odum, 1960), tout d’action sont disponibles (Ramenofsky deux prérequis et des connaissances
comme chez l’étourneau roselin (Sturnus et Wingfield, 2007). La migration de de la physiologie et du comportement
roseus) où le taux de lipides hépatiques printemps serait régulée par les hor- des palmipèdes, deux axes de travail
est plus que doublé en phase pré-migra- mones androgènes chez les femelles peuvent être exploités pour induire une
toire printanière (Pilo et George, 1983). La comme les mâles, en lien avec la mise stéatose spontanée : le niveau d’inges-
migration semble ainsi être associée à un en place de la reproduction à cette tion et le métabolisme lipidique.
engraissement h épatique variable selon période, alors que la migration autom-
les espèces. nale serait en partie régulée par les 2.1. Choix du modèle
hormones thyroïdiennes. L’implication animal et des inducteurs
Plusieurs facteurs environnemen- de ces régulations hormonales dans le potentiels de l’hyperphagie
taux sont responsables du comporte- comportement hyperphagique et la
ment d’hyperphagie pré-migratoire, mise en place d’un engraissement reste Trois paramètres induisant un com-
la photopériode étant le facteur pré- cependant peu documentée. L’effet de portement hyperphagique et un
pondérant chez les oiseaux migra- la prolactine a par exemple été étudié engraissement chez les palmipèdes
teurs sauvages (King et Farner, 1965 ; chez la caille des blés (Coturnix cotur- sauvages en période pré-migratoire
Bairlein et Gwinner, 1994 ; Cornelius nix, Boswell et al., 1995) et l’oie des ont été retenus pour être appliqués au
et al., 2013). Selon les espèces et la sai- neiges (Anser caerulescens caerulescens, niveau expérimental dans les premières
son, les conditions climatiques locales Campbell et al., 1981), mais son impli- études effectuées par l’INRA : la modu-
telles que la température, l’hygrométrie, cation dans l’engraissement pré-migra- lation de l’éclairement, de la tempéra-
le champ géomagnétique, la vitesse et toire n’a pas pu être mise en évidence, ture et de la disponibilité alimentaire.
la direction du vent, la disponibilité alors qu’elle a pu l’être chez le bruant
alimentaire et les interactions sociales à couronne blanche (Zonotrichia leuco- L’espèce domestique avec les meil-
modulent également ce comportement phyrs gambelii, Meier et Farner, 1964). leures prédispositions à la stéatose
(Cornelius et al., 2013). La température spontanée semble être l’oie grise lan-
influence ainsi indirectement la date de daise (Anser anser) qui est restée, d’un
départ de la migration d’automne via
2. La démarche point de vue génétique, proche de son
ses effets sur la disponibilité hydrique et de conception innovante ancêtre, l’oie cendrée sauvage migra-
alimentaire. De plus, pour une tempéra- utilisée trice (Heikkinen et al., 2015), et qui pré-
ture inférieure à la neutralité thermique, sente une forte capacité d’ingestion
l’engraissement est fortement corrélé et une bonne aptitude à la stéatose
à la température ambiante, affectant Un engraissement périphérique (tableau 1). C’est pourquoi, les premiers
ainsi directement la prise alimentaire, accompagné d’un engraissement travaux de l’INRA ont été conduits sur
comme c’est le cas chez la majorité des hépatique a ainsi été observé naturel- ce modèle animal. L’oie ne représentant
homéothermes (King and Farner 1965). lement chez certains oiseaux sauvages actuellement qu’une faible proportion
INRA Productions Animales, 2018, numéro 2120 / Christelle knudsen et al.
Tableau 1. Aptitudes à la stéatose spontanée des différents palmipèdes domes- la durée du jour en bâtiment obscur
tiques communément utilisés dans la filière foie gras (d’après Guy et al., 1997 ; (de 10 à 7 h/j). Les premiers résultats,
Baéza et al., 2005 ; Baéza et al., 2013).
obtenus en période hivernale ont per-
mis d’apporter une preuve de concept
Interactions en montrant la possibilité d’induire un
Capacité Comportement Aptitudes
possibles
d’ingestion migratoire physiologiques comportement hyperphagique transi-
avec la maturité
en 24h des ancêtres à la stéatose toire chez le jars après une période de
sexuelle
restriction alimentaire (Guy et al., 2013 ;
Oie grise *** Fernandez et al., 2016). La consomma-
**** *** **
des Landes (Oie cendrée) tion alimentaire était ainsi supérieure
à 400 g/j pendant les trois premières
Canard semaines d’alimentation au maïs,
** 0 **** ?
de Barbarie puis diminuait graduellement pour
atteindre des niveaux d’ingestion infé-
Canard rieurs à 300 g/j (figure 2). Ces niveaux
*** * *** ?
mulard
étaient donc supérieurs à ceux obser-
vés dans les rares études disponibles
Canard ***
** * ? chez l’oie adulte nourrie à volonté avec
commun (Colvert)
de l’aliment complet (340 et 380 g/j,
* : aptitude faible ; ** : aptitude modérée ; *** : aptitude forte ; **** : aptitude très forte ; ? : non déterminée. Sauveur et al., 1988 ; Arroyo et al.,
2012a) témoignant d’une hyperphagie
de la production française de foie gras 3. Les leviers d’actions transitoire. Ces travaux ont également
(< 2 %), par rapport à celle issue du utilisés et les résultats permis de tester les hypothèses avan-
canard, mulard principalement, l’INRA a obtenus cées quant aux facteurs induisant cette
également réalisé une expérimentation hyperphagie chez l’oie et d’initier leur
avec ce dernier modèle. Afin d’éviter la optimisation.
multiplication des facteurs expérimen- 3.1. Induction
taux et dans une démarche de réduc- et modulation b. L’éclairement
tion des effectifs animaux, chez l’oie de l’hyperphagie La lumière est un inducteur impor-
comme le canard les premiers essais tant du comportement d’hyperphagie
ont été menés exclusivement avec des Des méthodes d’induction de l’hy- pré-migratoire chez les oiseaux sau-
animaux d’un seul sexe. Les mâles ont perphagie impliquant la destruction du vages. Les travaux de Bonnefont et al
été choisis afin de s’affranchir des effets centre de satiété (noyau ventromédian (2015) ont permis d’évaluer son effet
de la ponte chez les femelles. de hypothalamus) ont été testées dans chez l’oie grise landaise en comparant
les années 1970 (Auffray et Blum, 1970). en période hivernale trois programmes
2.2. Choix du modèle Ce type de méthodes nous semble lumineux : maintien de 10 h d’éclaire-
alimentaire cependant peu applicable pour des ment (lot A) ; une diminution de l’éclai-
raisons éthiques et économiques. Nous rement de 10 à 7h/j par pas de 30 min
Lors de la phase pré-migratoire, avons donc fait le choix dans cette syn- tous les 2 jours à partir de la mise à
au-delà de l’hyperphagie, des modi- thèse de ne présenter que les systèmes disposition du maïs à volonté (lot B),
fications métaboliques, notamment développés répondant aux exigences tel que cela a été utilisé par Guy et al
dans le foie, favorisent également la définies précédemment. (2013) et Fernandez et al (2016) ; et un
lipogenèse et l’engraissement périphé- programme analogue au lot B réalisé
rique et hépatique (Guglielmo, 2018). a. Une stratégie d’élevage de façon plus précoce, qui permettait
Dans ce cadre, l’aliment a un rôle pri- basée sur le comportement d’atteindre 7h d’éclairement le jour de
mordial. Il doit être appétent pour migratoire la mise à disposition du maïs (lot C).
favoriser l’hyperphagie, avoir une forte Les protocoles des premiers tra- La consommation au cours de la pre-
valeur énergétique sous forme de pré- vaux amorcés par l’INRA en 2009 sur mière semaine d’accès au maïs était
curseurs glucidiques de la lipogenèse la stéatose spontanée ont été basés plus élevée dans le lot C que dans les
(amidon), avoir une bonne digestibilité sur le comportement migratoire des deux autres lots (436 vs 343 et 330 g/j),
et être pauvre en composés favorisant oiseaux sauvages et sur les pratiques même si sur l’ensemble de la période la
l’exportation des lipides dans les tissus et observations réalisées chez les ani- consommation de maïs était identique
périphériques (choline notamment). maux reproducteurs. Le protocole dans les 3 lots. L’hypothèse que le cycle
Le maïs répond à l’ensemble de ces cri- expérimental (figure 1) a ainsi consisté lumineux déclenche l’hyperphagie n’est
tères, ce qui en fait la matière première à appliquer une restriction alimentaire donc pas vérifiée, mais les résultats
privilégiée pendant le gavage (Baéza à la fin de la période de croissance sui- obtenus mettent en évidence l’intérêt
et al., 2013), et l’aliment retenu pour les vie d’une distribution à volonté de maïs d’une stimulation lumineuse précoce
premières études sur l’induction d’une pendant plusieurs semaines, de façon pour optimiser ce comportement. Des
stéatose hépatique spontanée. concomitante avec une réduction de études restent cependant à effectuer
INRA Productions Animales, 2018, numéro 2L’engraissement spontané du foie chez les palmipèdes : état des lieux et perspectives de recherche / 121
Figure 1. Schéma expérimental appliqué dans les travaux de Guy et al. (2013) et d’alimentation au maïs était réduite
de Fernandez et al. (2016) visant à induire une stéatose spontanée chez le jars (254 vs 316 g/j/oie, Fernandez et al,
(Anser anser).
résultats non publiés). Comme observé
chez l’oie sauvage, l’induction d’un
comportement hyperphagique chez
l’oie domestique au printemps requer-
rait donc d’utiliser d’autres facteurs
stimulants.
d. Modulation de la disponibilité
alimentaire
La modulation de la disponibilité
alimentaire est aussi un inducteur
potentiel du comportement hyper-
phagique chez les oiseaux migrateurs.
Dans le cadre de la conduite d’élevage
conventionnelle les animaux sont
soumis à une restriction alimentaire
horaire avant gavage afin de favoriser
Figure 2. Consommation de maïs inerté offert à volonté chez le jars (Anser anser) à une hyperphagie lors de l’accès à l’ali-
compter de 19 semaines d’âge après une phase de restriction alimentaire (d’après ment et ainsi développer le volume de
Guy et al., 2013 en rose ; Fernandez et al., 2016 en rouge). leur pseudo-jabot, et donc leurs capa-
cités d’ingestion (Arroyo et al., 2012b).
700
L’application d’une restriction alimen-
600
taire en amont de l’alimentation au
maïs serait dès lors un inducteur de l’hy-
500 perphagie. Les premières études chez le
Ingestion (g/j/oie)
canard ont également montré qu’une
400 modulation de la disponibilité alimen-
taire (restriction horaire de l’accès à la
300 mangeoire) durant l’alimentation au
maïs post restriction alimentaire induit
200 une augmentation transitoire de l’in-
gestion (Knudsen et al., 2017a) comme
100
cela a également pu être démontré
0 avec une alimentation restreinte aux
1 8 15 22 29 36 43 50 57 64 71 78 85 92 99 106 113 granulés (Guy et al., 1997).
Jours d’alimentation au maïs
3.2. Induction
de la stéatose hépatique
afin de déterminer plus finement l’effet c limatiques non contrôlées (bâtiment
de la stimulation lumineuse sur le com- non climatisé). Il serait donc intéressant Dans le cadre des travaux de Guy et al.
portement d’hyperphagie. d’évaluer l’effet de la température dans (2013) et de Fernandez et al. (2016), un
le cadre d’essais en ambiance contrôlée engraissement périphérique et hépa-
c. La température afin de définir et d’optimiser plus fine- tique a été observé (figure 3) puisque,
Comme chez la quasi-totalité des ment l’impact de la température sur après 12 semaines d’alimentation au
homéothermes, des températures le comportement d’hyperphagie des maïs, le poids de foie était de 500 g en
inférieures à la neutralité thermique palmipèdes. moyenne et le gras abdominal repré-
favorisent la prise alimentaire et l’en- sentait environ 570 g (7,2 % du poids
graissement chez les oiseaux migra- Enfin, un essai avec un protocole vif ). Toutefois, la variabilité du poids
teurs (King et Farner, 1965). Ceci a pu identique à celui de Fernandez et al. de foie était élevée (CV de 45 %, allant
être suggéré chez l’oie grise landaise (2016) a pu être mené au printemps, et d’environ 170 à 1 100 g). Au-delà de
dans le cadre des travaux de Guy et al. les niveaux d’hyperphagie étaient net- 12 semaines d’alimentation au maïs, le
(2013) et de Fernandez et al. (2016). tement inférieurs à ceux obtenus en poids de foie stagne et aucun engrais-
Ainsi, plus les températures sont basses, période hivernale. L’hyperphagie était sement hépatique supplémentaire
plus la consommation alimentaire est ainsi de plus courte durée (consom- n’est observé alors que l’engraisse-
élevée. Les essais menés jusqu’à pré- mation supérieure à 400 g/j pendant ment périphérique semble se stabili-
sent ont cependant été effectués en 2 semaines au lieu de 3) et la consom- ser plus rapidement, après 6 semaines
périodes hivernales, avec des conditions mation globale sur les 12 semaines d’alimentation au maïs. Les travaux de
INRA Productions Animales, 2018, numéro 2122 / Christelle knudsen et al.
Figure 3. Évolution du poids de foie en fonction de l’âge lors d’une alimentation L’aptitude à la stéatose hépatique sans
à volonté au maïs chez le jars (Anser anser) (d’après Guy et al., 2013 en rose ; alimentation forcée dépend donc en
Fernandez et al., 2016 en rouge).
premier lieu de la capacité d’hyperphagie
600 des oies durant les premières semaines
d’alimentation au maïs. Il serait intéres-
sant de vérifier ces corrélations dans des
500
conditions d’élevage collectif.
400 b. Modulation du métabolisme
Poids de foie (g)
lipidique
300 L’hyperphagie favorise fortement la
stéatose hépatique, mais d’autres fac-
200
teurs métaboliques rentrent également
en compte dans les mécanismes d’en-
graissement pré-migratoires (Cornelius
100 et al., 2013). En effet, chez l’oie grise
landaise, après la période d’hyperpha-
0 gie, la consommation à volonté de
19 21 23 25 27 29 31 33 35 37 maïs diminue alors que le poids de foie
Âge (semaines) continue d’augmenter (Guy et al., 2013 ;
Fernandez et al., 2016). L’engraissement
périphérique semble stable à partir de
Fernandez et al. (2015) ont comparé les a. Corrélation entre 6 semaines de maïs (Fernandez et al.,
performances du système de produc- hyperphagie et stéatose 2016), dépassant les niveaux observés
tion alternatif et du système de produc- Chez les oiseaux migrateurs sauvages, en gavage (7,2 % de PV vs 4,7-5,5 %,
tion conventionnel (avec gavage) chez l’engraissement est fortement corrélé à Hermier et al., 1999 ; Davail et al., 2000).
des oies grises landaises mâles. Les jars l’hyperphagie pré-migratoire. L’étude de Cela suggère une saturation de ces lieux
du système alternatif ont été abattus à Fernandez et al. (2016) a permis de vérifier de stockage, pouvant expliquer l’engrais-
224 j, après 12 semaines d’alimentation ce phénomène chez l’oie grise landaise et sement hépatique ultérieur. À l’opposé,
libre au maïs, alors que les autres jars de déterminer la corrélation entre inges- dans le cadre d’une stéatose induite par
ont été abattus à 115 j après 17 jours de tion et stéatose hépatique sur des oies gavage, une durée de jeûne prolongée,
gavage. Ces travaux montrent que pour logées individuellement. Dans ce dispo- ou une diminution de l’ingestion, pro-
des poids vifs de 8,2 kg et de 9,3 kg res- sitif, la consommation moyenne des oies voque une diminution du poids de foie
pectivement, le niveau d’engraissement était significativement inférieure à celle (Babilé et al., 1998 ; Arroyo et al., 2017).
de la carcasse obtenu est globalement d’animaux en groupe (– 29 % durant la Ainsi certaines stéatoses se prolongent
similaire, voir supérieur, à celui obtenu première semaine). Ceci s’explique par au-delà de la période d’hyperphagie. Il
après un gavage, alors que les poids la rupture du mode de vie grégaire des semble donc y avoir une modulation des
de foie sont très inférieurs (445 ± 24 animaux qui ne bénéficiaient donc plus mécanismes métaboliques qui se met
vs 1 102 ± 23), suggérant une lipoge- des effets d’entraînement du groupe en en place lors de la stéatose spontanée
nèse active, mais une exportation des matière de comportement alimentaire chez l’oie qu’il est important d’explorer.
lipides néo-synthétisés accrue. De plus, et qui étaient potentiellement en situa- Des travaux sont actuellement en cours
les foies issus d’animaux ayant déve- tion de stress de par leur isolement social, à l’INRA pour répondre à ce question
loppé une stéatose spontanée sont bien que les animaux pouvaient se voir et nement et définir la nature de la stéatose
légèrement moins engraissés (53 vs s’entendre. Les poids de foie de ces ani- mise en place.
56 % de lipides) et ont un rendement maux étaient également inférieurs à ceux
à la cuisson plus faible (82 vs 88 %). d’animaux élevés en groupe mais la varia- c. Effet de l’espèce animale
Leurs concentrations en glycogène bilité individuelle du poids de foie était Comme précisé précédemment, la
(103 vs 70 µmol/g) et en glucose (45 vs en revanche identique (CV de 62 et 63 %, variabilité interindividuelle de l’hy-
35 µmol/g) sont supérieures à celles des respectivement), ce qui a permis d’étudier perphagie et de la stéatose hépatique
animaux gavés, alors que les profils en les corrélations entre ingestion et stéatose est très forte chez l’oie grise landaise.
lipides neutres des foies sont similaires. dans des conditions représentatives de la Plusieurs pistes de recherches permet-
variabilité du caractère à ce stade. Cette tant d’expliquer ce phénomène, et donc
Ces résultats permettent donc d’af- étude a permis de mettre en évidence potentiellement de le réduire, existent
firmer qu’une stéatose spontanée est une corrélation forte entre le niveau d’in- et seront développées dans la partie 5.
possible chez l’oie grise landaise, même gestion de maïs et le poids de foie après 8,
si les performances obtenues sont infé- 11 et 13 semaines d’alimentation au maïs Les premiers travaux menés chez le
rieures à celles obtenues après gavage (r² = 0,6, 0,7 et 0,8 respectivement), avec canard mulard (Knudsen et al., 2017a)
des animaux, en particulier sur le plan un effet important de l’ingestion précoce ont montré que l’application d’une stra-
de l’homogénéité du poids de foie. (au cours des trois premières semaines). tégie d’élevage analogue à celle mise
INRA Productions Animales, 2018, numéro 2L’engraissement spontané du foie chez les palmipèdes : état des lieux et perspectives de recherche / 123
en place chez l’oie (Guy et al., 2013) ne à foie gras. La principale adaptation générés par la production de foies
permettait pas d’induire d’hyperphagie réalisée porte sur l’évaluation du pilier engraissés, obtenu par gavage ou de
durable. Des essais préliminaires chez environnemental pour lequel l’analyse façon spontanée grâce à une ACV. Les
le canard de Barbarie ont indiqué des des cycles de vie (ACV) a été utilisée plu- impacts environnementaux de la pro-
résultats similaires (Guy et al, données tôt que des indicateurs de pratiques au duction d’un kilogramme de foie sont
non publiées). L’hyperphagie engen- niveau de l’atelier, répondant en cela ainsi plus importants pour le système
drée était en effet de très courte durée aux recommandations de Lairez et al. où la stéatose est obtenue de façon
(570 g d’ingestion sur la première jour- (2015). spontanée (tableau 2, figure 4). Ces
née), le canard régulant très rapide- résultats s’expliquent principalement
ment son ingestion lors de la transition 4.1. Performances par une durée d’engraissement plus
alimentaire, confirmant les résultats environnementales longue (12 semaines vs 16 jours) et une
obtenus chez le canard en croissance consommation d’aliment accrue (64,1 vs
préalablement restreint (Guy et al., Les travaux de Brachet et al., (2015b) 42,6 kg par oie et 124,5 vs 52,3 kg par
1997). Cette brève hyperphagie lors ont permis de déterminer les impacts kg de foie) pour des poids de foie infé-
d’une alimentation au maïs à volonté environnementaux potentiellement rieurs (514 vs 815 g, tableau 3). L’aliment
était associée à une augmentation
négligeable du poids de foie (84 g de Tableau 2. Impacts environnementaux de la production d’un kg de foie chez le
jars (Anser anser) dans les systèmes conventionnel (avec gavage) et alternatif (sans
foie en moyenne au point culminant, gavage) (d’après Brachet et al., 2015b).
après 2 semaines d’alimentation au
maïs, vs 58 g avant alimentation au
Impact potentiel Conventionnel Alternatif
maïs). Une très forte variabilité de l’ap-
titude à la stéatose spontanée inter-
Potentiel de changement climatique (kg CO2-eq.) 53,02 140,55
espèces semble donc être observée et
les effecteurs de l’hyperphagie et de la
Potentiel d’eutrophisation (kg PO43--eq.) 0,37 0,84
stéatose semblent être très dépendants
de l’espèce, comme c’est aussi le cas Potentiel d’acidification (kg SO2-eq.) 0,75 1,74
chez les oiseaux migrateurs sauvages
comme présenté précédemment. Potentiel de toxicité terrestre (kg 1,4-DB-éq) 0,15 0,32
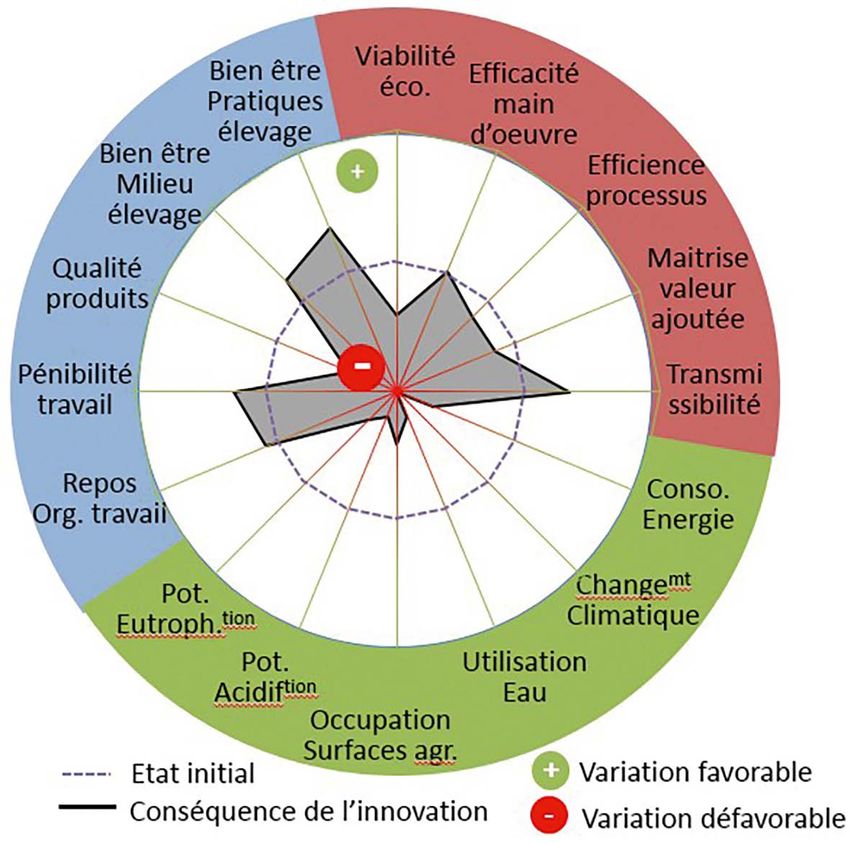
4. Évaluation Consommation d’énergie primaire (MJ-eq) 406,66 905,62
de la durabilité
Consommation de la ressource en eau (m3) 3,44 8,16
du système innovant
Occupation des surfaces (m2.an) 66,74 142,68
Comparativement au système
conventionnel, les performances pon- Figure 4. Évaluation de la durabilité d’un système de production de foie engraissé
dérales en stimulation de la stéatose résultant d’une stéatose spontanée chez le jars (Anser anser) (Cadre conceptuel
spontanée sont en moyenne moi- adapté de Litt et al., 2014 ; et résultats d’après Brachet et al., 2015b ; Fernandez
et al., 2015).
tié moindres que dans le cadre d’un
gavage (500 vs 1 000 g de poids de
foie) et la variabilité individuelle de ces
performances est deux à trois fois plus
élevée qu’à l’issue d’un gavage (45-60 vs
20 %) pour une durée d’engraissement
cinq fois plus élevée (12 semaines vs 16
jours) (Guy et al., 2013 ; Fernandez et al.,
2016). Ce système d’élevage innovant,
et le produit qui en est issu, est donc
très différent d’un système convention-
nel et il convient de définir ses impacts
en termes de durabilité. Ici, nous avons
fait le choix d’évaluer la durabilité selon
ses trois piliers fondateurs : le pilier
environnemental, le pilier économique
et le pilier social. Pour cela, nous avons
adapté le cadre conceptuel proposé par
Litt et al. (2014) pour évaluer la durabi-
lité des ateliers d’élevage de p almipèdes
INRA Productions Animales, 2018, numéro 2124 / Christelle knudsen et al.
Tableau 3. Principales caractéristiques et performances zootechniques dans les La qualité du produit a été com-
systèmes de production conventionnel (foie gras issu du gavage) et alternatif (foie parée pour des foies issus d’animaux
engraissé spontanément) chez le jars (Anser anser) (d’après Brachet et al., 2015b).
engraissés spontanément ou gavés
via des analyses sensorielles et des
Conventionnel Alternatif tests d’acceptabilité par les consom-
mateurs. Ceux-ci indiquent que, dans
IC(1) Démarrage– Élevage 4,28 5,73 une même gamme de poids, les foies
obtenus par engraissement spontané
ITM(2) Gavage/engraissement 17,55 54,37
sont moins bien notés et moins bien
appréciés que les foies issus du gavage
Quantité d’aliment consommée (kg/oie)
(Fernandez et al., 2015) notamment sur
des critères d’aspect, d’odeur et de goût
Démarrage 9,3 8,9
(tableau 4). Il semble dès lors pertinent
d’explorer les mécanismes sous-jacents
Élevage 19,0 27,2
à la stéatose spontanée pour un objec-
tif finalisé d’optimisation du système de
Gavage/engraissement 14,3 28,0
production.
Âge d’abattage (jours) 115 224
L’acte d’embucage est supprimé avec
Poids d’abattage (g) 9 280 8 267 ± 963
le système d’élevage basé sur la stéa-
tose spontanée et les animaux sont
Poids de foie (g) 815 514 ± 231 élevés en groupe et au sol durant toute
la durée d’élevage. Ce sont des points
Muscle magret/foie 0,33 0,80 ± 0,54 très positifs en termes de réponse aux
demandes des citoyens consomma-
Mortalité (%) teurs (figure 4), et qui justifieraient le
développement du système de pro-
Démarrage– Élevage 5 7 duction alternatif au gavage. La maitrise
de l’éclairement impose cependant une
Gavage/engraissement 1 4 claustration des animaux sur une grande
partie de la durée d’élevage pouvant
(1)
: IC : Indice de Consommation (aliment consommé/gain de poids vif ). avoir un impact potentiellement néga-
(2)
: ITM : Indice de Transformation du Maïs (maïs consommé pendant l’engraissement/poids de foie).
tif sur le bien-être animal. Des études
comparatives spécifiques seraient donc
et les déjections animales expliquent ce qui pourrait dégrader certaines nécessaires pour définir précisément
90 % des impacts environnementaux. performances environnementales. l’impact de ces deux systèmes sur dif-
Une optimisation du système est donc Dans cette évaluation le système sera férentes composantes du bien-être
nécessaire afin de diminuer les impacts donc considéré comme saisonnalisé. animal au cours des différentes phases
environnementaux. Les autres paramètres de ce pilier (la d’élevage. Enfin, les animaux n’étant
viabilité économique, l’efficacité de la pas gavés, le système proposé s’affran-
4.2. Performances main d’œuvre, l’efficience du proces- chit des contraintes posturales liées au
économiques sus, la maitrise de la valeur ajoutée) gavage et est moins exigeant en temps
sont quant à eux difficiles à évaluer en de travail et donc en main d’œuvre. Une
Pour être durable, un atelier d’éle- l’absence de marché réel. Toutefois, la réduction de la pénibilité au travail et
vage de palmipèdes à foie gras doit faible quantité de produit obtenu pour une augmentation du temps de repos
être rentable, flexible et adaptable l’instant, et sa saisonnalité, oblige- peuvent donc être escomptées avec ce
(Litt et al., 2014). On peut supposer raient à proposer un prix très au-delà système.
que la flexibilité du système alterna- du prix actuel du foie gras pour couvrir
tif est améliorée dans la mesure où il les coûts de production. Le système alternatif au gavage,
s’affranchit des investissements, très tel qu’il est expérimenté aujourd’hui
importants, liés à la salle de gavage, 4.3. Performances pourrait donc permettre d’améliorer
mais dégradée par la saisonnalité de la sociales les performances en termes de bien-
production (actuellement une bande être animal et de qualité de vie de l’éle-
par an en hiver). La production pour- Litt et al (2014) ont défini que, dans le veur, mais dégraderait l’ensemble des
rait être dessaisonnée mais implique- pilier social, pour être durables les sys- autres performances de la durabilité
rait des investissements importants tèmes doivent produire des produits de (figure 4). À ce stade, une optimisation
pour contrôler la photopériode et la qualité, respecter le bien-être animal, et du système est nécessaire ainsi qu’une
température des bâtiments, augmen- préserver les conditions de travail et la évaluation plus fine et complète de sa
tant ainsi la consommation d’énergie qualité de vie de l’éleveur. durabilité.
INRA Productions Animales, 2018, numéro 2L’engraissement spontané du foie chez les palmipèdes : état des lieux et perspectives de recherche / 125
Tableau 4. Appréciation des foies de jars (Anser anser) selon le système de pro- à améliorer le bien-être animal durant
duction, conventionnel (foie gras issu du gavage) ou alternatif (foie engraissé la phase de restriction alimentaire
spontanément) et la gamme de poids dans le cadre de tests d’acceptabilité par le
(Brachet et al., 2015a).
consommateur (d’après Fernandez et al., 2015).
b. Conduite d’élevage
Stimulés Stimulés
Gavés P-values Les premiers travaux sur la stéatose
600-800 g 800-1 000 g
spontanée chez l’oie domestique ont
Aspect 6,38 ± 1,84b 5,00 ± 2,35a 5,28 ± 2,23a *** montré un fort impact des conditions
environnementales sur le comporte-
Odeur 6,43 ± 1,84b 5,86 ± 2,07ab 5,59 ± 2,07a *** ment d’hyperphagie.
Goût 6,90 ± 1,78b 5,90 ± 2,21a 5,23 ± 2,39a * Jusqu’à présent, seule la durée d’éclai-
rement a pu être étudiée en période
Texture 6,91 ± 1,73b 5,28 ± 2,38a 5,35 ± 2,33a *** hivernale chez l’oie (Bonnefont et al.,
2015). Plusieurs autres facteurs, comme
Appréciation globale 6,83 ± 1,55b 5,77 ± 2,16a 5,59 ± 2,07a *** la nature du spectre lumineux et son
intensité pourraient cependant avoir
* : p < 0,05 ; *** : p < 0,001 ; a, b : des lettres différentes entre moyennes d’une même ligne indiquent un impact significatif sur le compor-
des différences significatives au seuil de 5 %.
tement d’ingestion, les performances
pondérales et le bien-être des palmi-
5. Perspectives sur le niveau d’ingestion des animaux. pèdes, comme c’est le cas chez les pou-
de recherche Plusieurs facteurs favorisant son inges- lets de chair (Mendes et al., 2013 ; Parvin
tion peuvent ainsi être travaillés : son et al., 2014). Concernant la température,
appétence, et sa capacité d’hydratation des études en conditions climatiques
Nous avons fait le choix dans cette syn- et d’induction d’une satiété. contrôlées permettraient d’évaluer
thèse de présenter les perspectives de précisément son effet sur l’hyperpha-
recherche possibles pouvant répondre Les palmipèdes, comme la majorité gie et la stéatose, et ainsi de définir des
au mieux à l’ensemble des critères de des oiseaux, sont sensibles à l’aspect itinéraires techniques optimaux. Enfin,
durabilité énoncés précédemment sur visuel (couleur) et tactile (taille et tex- les premiers travaux chez le canard ont
ses trois piliers fondateurs : le pilier envi- ture des particules) des aliments (Owen, montré qu’une modulation de la durée
ronnemental, le pilier économique et le 1978), ainsi qu’à leur flaveur (Arroyo de mise à disposition de l’aliment per-
pilier social en évitant de léser de façon et al., 2012b). Ainsi, selon l’appétence mettait de relancer brièvement l’hyper-
importante un pilier au profit d’un autre. de ceux-ci la consommation sera plus phagie (Guy et al., 1997 ; Knudsen et al.,
Pour répondre à ces critères, le système ou moins élevée. Bien que le maïs soit 2017a). Cette modulation, si elle est
d’élevage mis en place pour stimuler la déjà un aliment très appétent, l’hyper- vérifiée chez l’oie et le canard, permet-
stéatose spontanée doit être optimisé phagie pourrait être stimulée en opti- trait dès lors de maximiser l’ingestion
afin de : i) répondre aux enjeux sociétaux misant l’appétence et la préhension de sur une période donnée. La maitrise des
et améliorer l’acceptabilité du produit l’aliment par les palmipèdes, en modi- paramètres d’ambiance peut cepen-
auprès du consommateur ; ii) réduire son fiant sa forme et sa présentation par dant avoir un impact sur le bien-être
impact environnemental ; iii) réduire son cuisson par exemple. animal qu’il conviendrait d’évaluer.
coût de production. Afin de répondre à Enfin, bien que les performances pon-
ces enjeux, la conduite d’élevage doit être La capacité d’ingestion est également dérales puissent être dégradées, une
optimisée, avec pour objectif d’entraîner fonction de paramètres physiologiques conduite d’élevage en lumière natu-
des comportements d’hyperphagie et inhérents à l’animal, à savoir la vitesse relle pourrait être envisagée afin de se
d’améliorer les performances hépatiques, de transit et la capacité volumique de rapprocher au mieux des conditions de
sans dégrader les rendements en viande, son tractus digestif, le pseudo-jabot vie normales de l’animal, en permet-
tout en répondant aux prérequis énoncés notamment. La taille de celui-ci peut tant en particulier l’accès à un parcours
d’absence d’alimentation forcée et d’ex- être développée durant la phase de extérieur. L’impact sur les performances
pression du comportement normal de croissance à l’aide d’aliments à haute pondérales, économiques et environne-
l’animal. Les poids de foie et leur niveau capacité d’hydratation, et donc à fort mentales de ce type de système mérite-
de stéatose doivent être homogénéisés pouvoir gonflant (Arroyo et al., 2012b ; rait également d’être évalué.
tout en améliorant leur qualité sensorielle. Brachet et al., 2015a). L’utilisation d’ali-
ments de ce type pourrait dès lors être Enfin, dans un souci de durabilité,
5.1. Induire et maintenir envisagée durant la phase de restriction l’utilisation des deux sexes dans le dis-
une hyperphagie alimentaire permettant ainsi de déve- positif de stéatose spontanée serait
lopper les capacités d’ingestion des préférable, et des travaux sur cette
a. Facteurs alimentaires animaux. De plus, la forte occupation thématique sont en cours. La maturité
Comme précisé en partie 2, le choix du tractus digestif favorise la sensation sexuelle n’ayant pas lieu de façon syn-
de l’aliment joue un rôle primordial de satiété, et pourrait donc contribuer chrone chez les mâles et les femelles,
INRA Productions Animales, 2018, numéro 2Vous pouvez aussi lire

















































