Modulation de l'expression de Med15 au foie dans le vieillissement et l'obésité - Mémoire Kevin Gonthier Maîtrise en sciences pharmaceutiques ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Modulation de l'expression de Med15 au foie dans le
vieillissement et l'obésité
Mémoire
Kevin Gonthier
Maîtrise en sciences pharmaceutiques - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Kevin Gonthier, 2019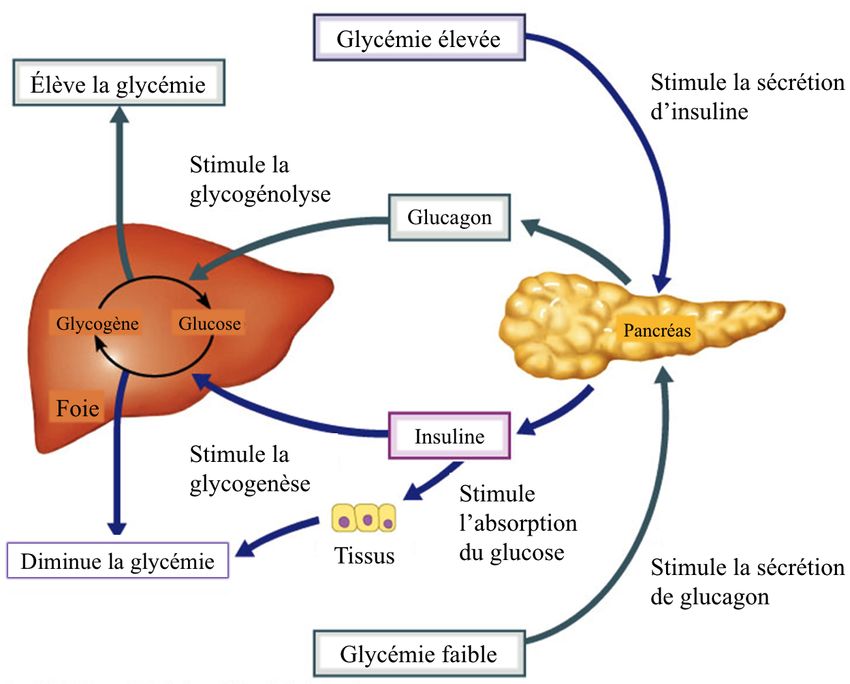
Modulation de l’expression de Med15 au foie dans le
vieillissement et l’obésité
Mémoire
Kevin GONTHIER
Sous la direction de :
Frédéric Picard, directeur de recherche
ii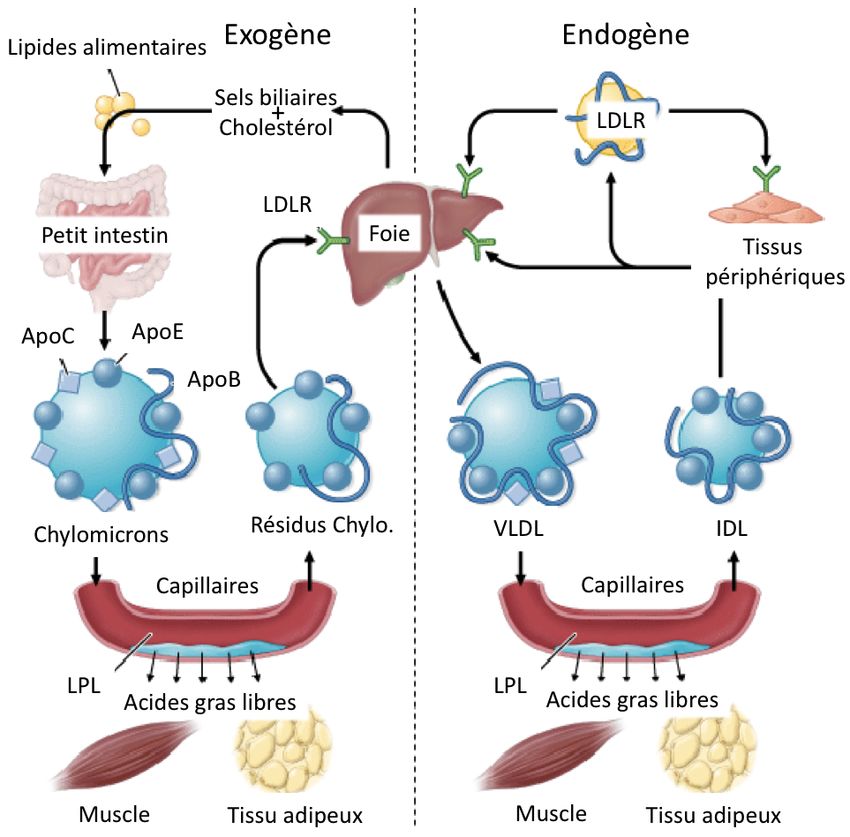
RÉSUMÉ
Chez le nématode Caenorhabditis elegans (C. elegans), le cofacteur mdt-15, orthologue à
Med15 chez le mammifère, est essentiel à l’homéostasie des lipides. Aussi, l’inhibition
pharmacologique de l’interaction entre Med15 et le facteur de transcription SREBP améliore
le profil lipidique chez des souris obèses. Le foie, organe important du métabolisme
énergétique, peut subir des dérèglements lors du vieillissement et dans l’obésité. La
modulation des niveaux hépatiques de Med15 dans ces conditions pathologiques est toutefois
inconnue. Cette étude visait donc à évaluer l’expression de Med15 au foie dans le
vieillissement et l’obésité. Les modèles de vieillissement étaient des souris C57Black/6J
(B6), des rats Sprague-Dawley (SD) et des rats Lou (modèle de vieillissement réussi) de
différents groupes d’âges et sous diète faible en gras (LFD). MED15 a également été mesuré
dans du foie humain provenant de 3 groupes de patients obèses jeunes, d’âge intermédiaire
et vieux. Afin de dissocier les effets du vieillissement de ceux de l’obésité, Med15 a été
mesuré dans des souris ob/ob ou db/db sous LFD et des souris B6 sous diète riche en gras
(HFD) âgées de 4 mois. L’expression de Med15 était diminuée chez les souris B6 et les rats
SD âgés mais demeurait stable chez les rats Lou vieux et les patients âgés. Les niveaux de
Med15 étaient diminués chez les souris ob/ob et db/db. En revanche, ses niveaux protéiques
hépatiques étaient augmentés chez les souris sous HFD. La conclusion générale, tirée des
liens établis entre les résultats présentés ici et la littérature, est que l’expression de Med15
serait bénéfique chez un organisme sain mais que sa diminution permettrait de freiner les
troubles métaboliques associés au vieillissement et à l’obésité. Des études in vitro et in vivo
sur les impacts des variations observées dans cette étude permettraient de caractériser Med15
comme modulateur métabolique chez le mammifère.
iii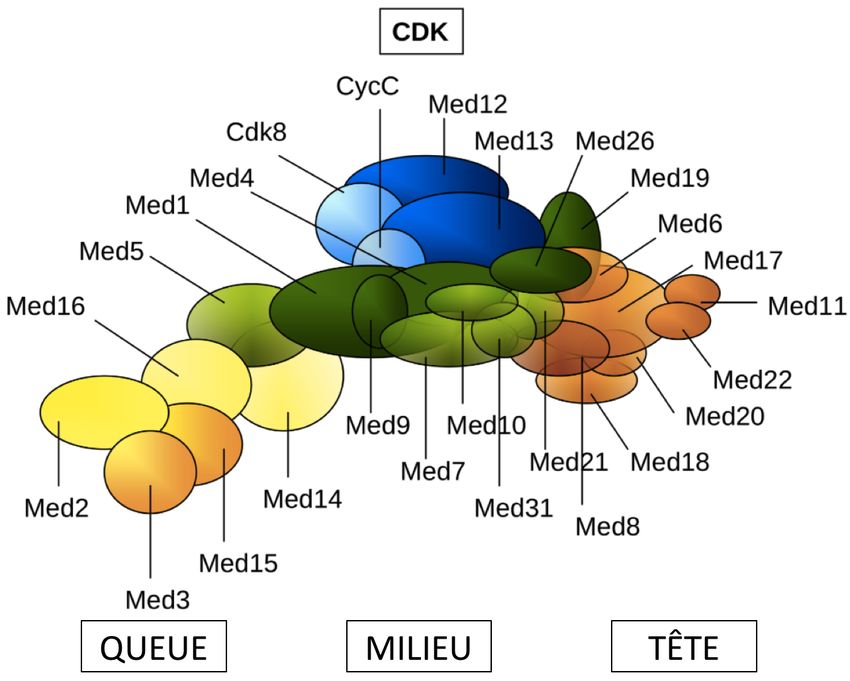
ABSTRACT
In the nematode Caenorhabditis elegans (C. elegans), the mdt-15 cofactor, orthologous to
the mammalian Med15, is essential for lipid homeostasis. Furthermore, pharmacological
inhibition of the interaction between Med15 and Sterol Regulatory Element Binding Protein
(SREBP) transcription factor improves the lipid profile in obese mice. The liver, important
organ of energy metabolism, may undergo disorders during aging and in obesity. Modulation
of Med15 hepatic levels under these two conditions is however unknown. This study aimed
therefore to evaluate hepatic Med15 expression in several aging and obesity models. The
aging models were C57Black/6 Jackson (B6) mice, Sprague-Dawley (SD) rats and Lou rats
(a successful aging model) in different age groups and under low-fat diet (LFD). MED15
was also measured in human liver from 3 groups of obese young, middle-aged and old
patients. In order to dissociate the effects of aging from those of obesity, Med15 expression
was measured in 4 months old ob/ob or db/db mice under LFD and B6 mice under high-fat
diet (HFD). Med15 expression was decreased in old B6 mice and SD rats but remained stable
in old Lou rats and elderly patients. Med15 levels were diminished in ob/ob and db/db mice.
However, Med15 protein levels were increased in mice under HFD. The general conclusion,
drawn from links established between the results presented here and the literature, is that
Med15 expression would be beneficial in a healthy organism but its decrease would curb the
metabolic disorders associated with aging and obesity. In vitro and in vivo studies on the
impacts of the variations observed in this study would allow for the Med15 characterization
as a key metabolic modulator in mammals.
iv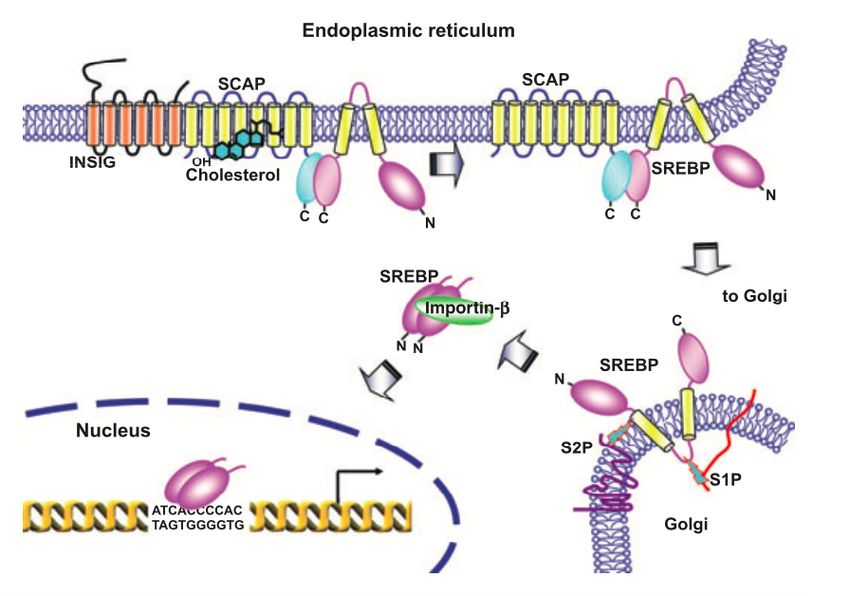
TABLE DES MATIÈRES
Résumé ............................................................................................................................ iii
Abstract ........................................................................................................................... iv
Table des matières ........................................................................................................... v
Liste des tableaux............................................................................................................ vi
Liste des figures ............................................................................................................. vii
Liste des abréviations ................................................................................................... viii
Remerciements ................................................................................................................. x
1 Introduction .............................................................................................................. 1
1.1 Métabolisme énergétique ................................................................................... 1
1.1.1 Sentiers métaboliques et hormones........................................................ 1
1.1.2 Le foie, centre intégrateur du métabolisme énergétique ......................... 4
1.1.3 Désordres fonctionnels au foie et pathologies associées......................... 6
1.2 Acteurs et mécanismes moléculaires .................................................................. 9
1.2.1 Facteurs de transcription ....................................................................... 9
1.2.2 Récepteurs nucléaires ...........................................................................11
1.2.3 Corégulateurs .......................................................................................14
1.3 Complexe Mediator ..........................................................................................15
1.3.1 Structure et versatilité fonctionnelle .....................................................15
1.3.2 Fonctions primaires et régulatrices .......................................................16
1.3.3 Importance métabolique .......................................................................16
1.4 Cofacteur nucléaire Med15 ...............................................................................18
1.4.1 Études chez la levure............................................................................19
1.4.2 Études chez le nématode ......................................................................20
1.4.3 Études chez le mammifère ...................................................................22
2 Chapitre Justification ..............................................................................................25
3 Chapitre Description des modèles à l’étude ............................................................27
4 Chapitre Méthodologie ............................................................................................31
5 Chapitre Résultats....................................................................................................35
6 Chapitre Discussion .................................................................................................41
7 Conclusion.....................................................................................................................52
8 Bibliographie ............................................................................................................53
v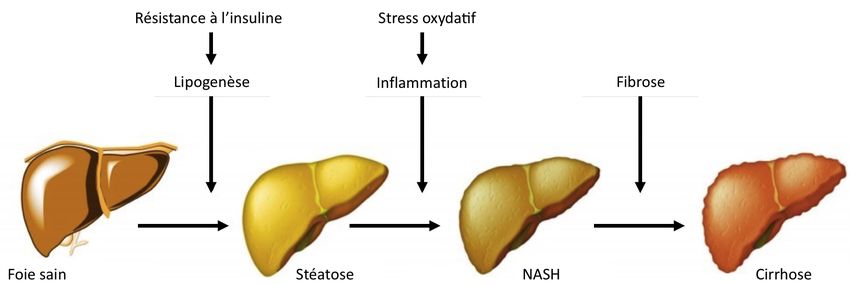
LISTE DES TABLEAUX
Tableau 1 Comparaisons de l’identité de la séquence du gène Med15 chez l’humain....22
Tableau 2 Paramètres physiologiques des patients composant la cohorte ......................30
Tableau 3 Séquences des amorces des gènes mesurés chez la souris .............................32
Tableau 4 Séquences des amorces des gènes mesurés chez l’humain ............................32
vi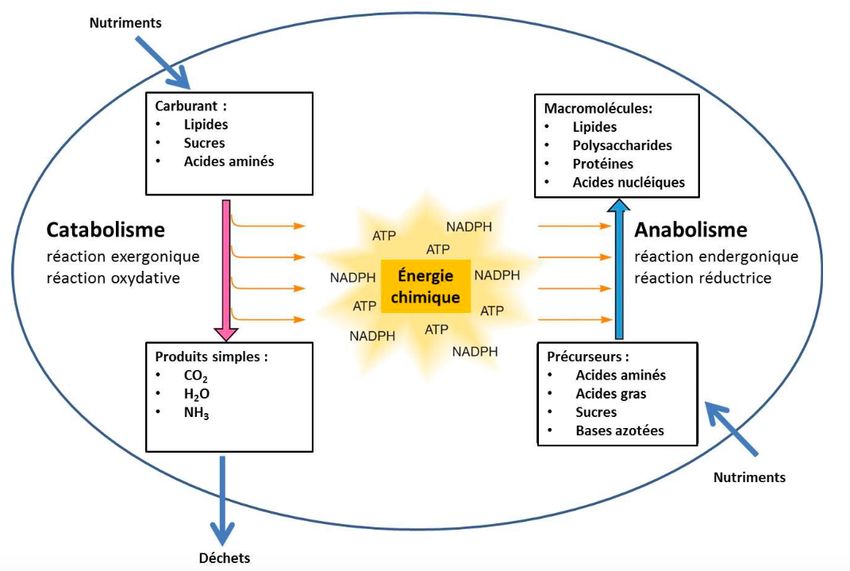
LISTE DES FIGURES
Figure 1 Composantes du métabolisme et substrats énergétiques ................................... 1
Figure 2 Voies du métabolisme des lipides .................................................................... 5
Figure 3 Régulation de la glycémie ............................................................................... 6
Figure 4 Progression de la maladie hépatique ................................................................ 8
Figure 5 Processus de maturation du facteur de transcription SREBP ...........................10
Figure 6 Fonctions des PPARs .....................................................................................13
Figure 7 Architecture du corégulateur transcriptionnel Mediator ..................................15
Figure 8 Contrôle transcriptionnel par le cofacteur nucléaire Med15 ............................24
Figure 9 Expression relative d’ARNm Mediator et des protéines Med15 dans du foie de
Figure 9 souris mâles au cours du vieillissement ..........................................................35
Figure 10 Évaluation des niveaux protéiques du gène Med15 dans du foie de rats SD
Figure 10 vieillissants sous RC ou non et dans du foie de rats Lou à différents âges .......37
Figure 11 Expression relative d’ARNm Mediator et des protéines MED15 dans du foie
Figure 11 de patients obèses jeunes, d’âge intermédiaire et vieux ...................................38
Figure 12 Expression relative d’ARNm Mediator et des protéines Med15 dans du foie de
Figure 12 souris +/+, ob/ob et db/db ...............................................................................39
Figure 13 Expression relative d’ARNm Mediator et des protéines Med15 dans du foie de
Figure 13 souris soumises au HFD ou au LFD ...............................................................40
vii
LISTE DES ABRÉVIATIONS
15dPGJ-2 15-désoxy-delta12,14-prostaglandine J2
ABC Adenosine Triphosphate Binding Cassette
ABCA1 ATP Binding Cassette A1
AC Acétyl-CoA
ACC Acétyl-CoA carboxylase
ACLY ATP citrate lyase
ADNc ADN complémentaire
AL ad libitum
AMPK Adenosine Monophosphate-activated Protein Kinase
ANOVA Analyse de la variance (ANOVA)
Apo-CIII Apolipoprotéine C-III
ApoB Apolipoprotéine B
ARN Pol II ARN Polymérase II
ARNi ARN interférence
ARNm ARN messager
B6 C57BL/6J
bHLH-Zip Hélice-Boucle-hélice & Glissière à Leucine
BMI Indice de masse corporelle
C. elegans Caenorhabditis elegans
CERUL Comité d’éthique de la protection des humains
ChIP Immunoprécipitation de chromatine
Ck2 Caséine kinase 2
Co-IP Co-immunoprécipitation
CPAUL Comité d’éthique de la protection des animaux de
l’Université Laval
CREB cAMP Response Element Binding Protein
CREBBP CREB-Binding Protein
CYP7A1 Cholesterol 7 alpha-hydroxylase
Diète riche en gras HFD
ERK Extracellular signal-regulated kinase
FASN Acide gras Synthase
FOXO Forkhead box O
FXRs Farsenoid X receptors
GA Appareil de Golgi
GLUT Transporteur de glucose
GTFs Facteurs généraux de transcription
HDL Lipoprotéine de haute densité
HNF4α Facteur nucléaire hépatocytaire 4 alpha
IDL Lipoprotéine de densité intermédiaire
INSIG1 Insulin Induced Gene 1
INSIG2 Insulin Induced Gene 2
Insulin-like Growth Factor-1 IGF-1
KIX Kinase-inducible domain interacting
viii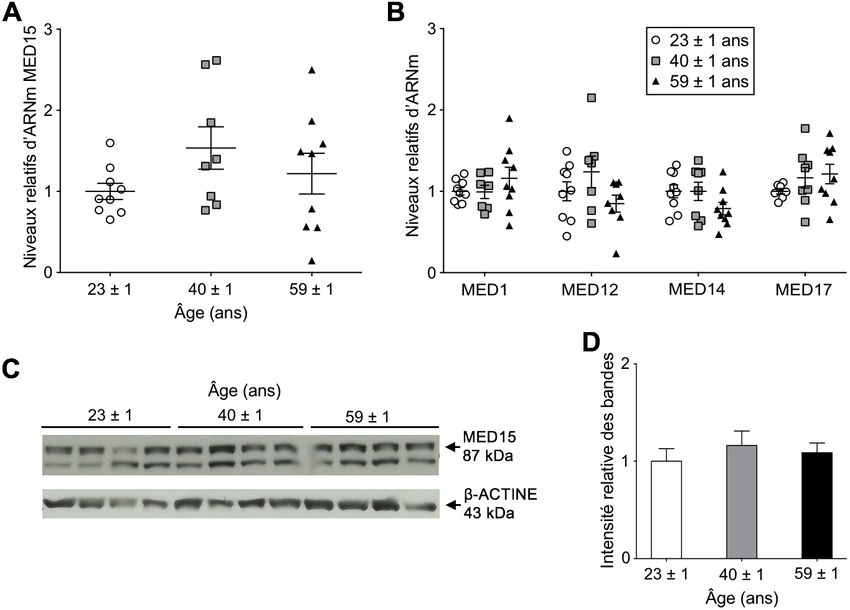
LDL Lipoprotéine de faible densité
LDL-R Récepteur aux LDL
LFD Diète Faible en Gras
LPL Lipoprotéine Lipase
LXR Liver X Receptor
LXRE Élément de réponse LXR
MAPK Mitogen-activated protein kinase
MetS Syndrome métabolique
mTOR mechanistic target of Rapamycin
MTTP Microsomal Triglyceride Transfer Protein
NAFLD Stéatose hépatique non alcoolique
NASH Stéatohépatite non alcoolique
NHR-49 Nuclear Hormone Receptor-49
OAF1 Oleate-Activated Transcription Factor
ob Obese gene
ob-r Récepteur à la leptine
p300 Histone acétyltransférase p300
PDR Pleiotropic Drug Resistance
PIC Complexe de pré-initiation
PPARs Récepteur active par les proliférateurs de peroxysomes
PVDF Polyfluorure de Vinylidène
qPCR Chaîne par Polymérase en temps réel
RC Restriction calorique
RQRV Réseau Québécois de Recherche sur le Vieillissement
RT Transcription Inverse
RXR Retinoid X receptor
S. cerevisiae Saccharomyces cerevisiae
S1P Site-1P
S2P Site-2P
sbp-1 Sterol Regulatory Element Binding Protein
SCAP SREBP Cleavage Activating Protein
SD Sprague-Dawley
SDS-PAGE électrophorèse sur gel de polyacrylamide en présence
de dodécylsulfate de sodium
SRE Sterol Response Element
SREBP Sterol regulatory element binding protein
T° Température
T2D Diabète de type 2
T3 Hormone Thyroïdienne Triiodothyronine
TAD Domaine de Transactivation
TFs Facteurs de transcription
TGF-β Facteurs de Croissance Transformant de la classe β
USF1 Upstream stimulatory factor 1
VLDL Lipoprotéine de très basse densité
ix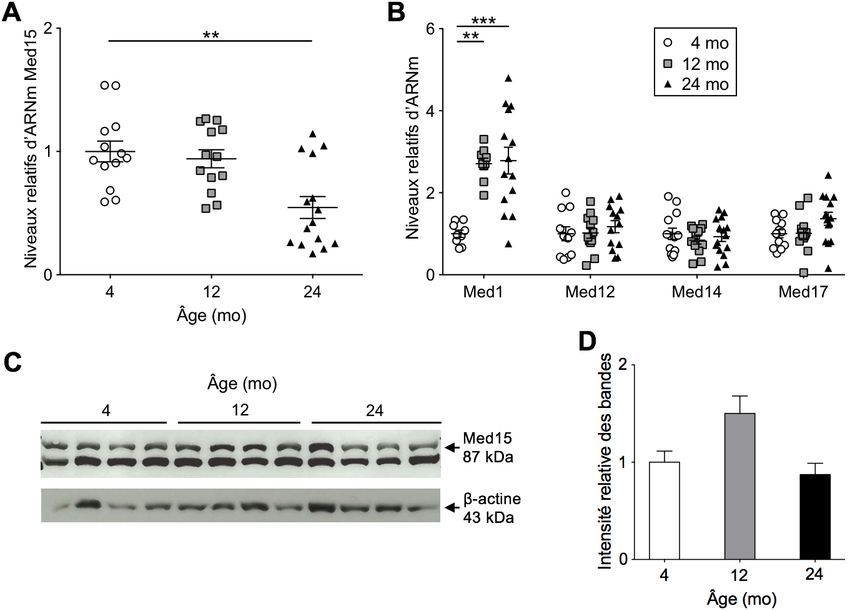
REMERCIEMENTS
Je voudrais tout d’abord exprimer toute ma gratitude au Dr Frédéric Picard, qui m’a permis
d’intégrer le fascinant monde de la recherche en tant que stagiaire dans son laboratoire et qui
m’a accompagné dans mon cheminement au 2ème cycle en tant que mon superviseur. Il a
toujours été présent pour répondre à mes nombreuses interrogations sur une diversité de
sujets liés à la recherche. Son enseignement sur la communication écrite et orale de la science
a aussi été très formateur. J’ai énormément appris à son contact. De plus, il a su m’inspirer à
poursuivre mon parcours dans la voie de la recherche biomédicale. Je mettrai tout en œuvre
pour faire honneur au training scientifique rigoureux qu’il m’a fourni.
Merci à Stéphanie, professionnelle de recherche du laboratoire. Sa supervision lors de mon
apprentissage de nouvelles techniques fut appréciée. Ses recommandations et solutions en
regard aux difficultés rencontrées dans les manipulations ont été d’une grande aide. Je
remercie également les autres membres de l’équipe.
Chloé, ma chère amie et collègue de laboratoire, toujours d’humeur radieuse et accueillante.
Les nombreux souvenirs que j’ai eu la chance de forger avec elle au fil de ces mois au
laboratoire (et à la cafétéria) me sont très précieux.
J’aimerais remercier le Dr Mathieu Laplante, avec qui j’ai eu la chance de partager plusieurs
conversations intéressantes et enrichissantes en regard au milieu scientifique. Merci pour ses
conseils en matière de cheminement de carrière.
Un merci également aux gens qui composent l’équipe du Dr Laplante. Yves, qui doit à tout
prix préserver son sens de l’humour original. Ce fut un vrai plaisir de travailler à ses côtés.
Romain, l’homme qui faisait des Western Blots, avec qui j’ai pu échanger des avis aussi bien
sur les manips que sur l’entraînement en salle. Mathilde, l’étudiante matinale, avec laquelle
j’ai pu partager diverses conversations toutes plus intéressantes les unes que les autres.
xMerci tout particulièrement à Marie-Pier, ma compagne de vie, qui a été présente à chaque
moment. Ses encouragements, son support et son écoute constante dans mon processus
d’apprentissage de l’équilibre entre les énergies consacrées à la recherche et celles qui
doivent être investies dans des aspects plus personnels et qui sont nécessaires pour « garder
le cap », sont un vrai cadeau. Merci!
Je veux aussi exprimer toute ma reconnaissance envers ma famille, qui m’a toujours supporté
de bien des manières, et ce de façon inconditionnelle. Elle m’a accompagné et épaulé à
chaque étape de mon parcours universitaire. Merci infiniment.
Merci au Dre Pierrette Gaudreau ainsi qu’au Réseau Québécois de Recherche sur le
Vieillissement (RQRV) pour le support financier
xi1. INTRODUCTION
1.1 MÉTABOLISME ÉNERGÉTIQUE
1.1.1 Sentiers métaboliques et hormones
Le métabolisme représente l’ensemble des réactions biochimiques impliquées dans la
distribution, la conversion et l’utilisation de divers substrats comme les lipides, les glucides
et les protéines (Fig. 1 – adaptée de (1)). Les réactions de transformations de ces biomolécules
sont les sentiers métaboliques (1).
Fig. 1: Composantes du métabolisme et substrats énergétiques (adaptée de (1)).
Les lipides remplissent plusieurs fonctions biologiques telles que le stockage de l'énergie, la
composition de membranes cellulaires et la coordination de processus de signalisation (2).
La lipogenèse caractérise les réactions de synthèse (anabolisme) d’acides gras à partir
d’acétyl-CoA (AC) et de la conversion subséquente des acides gras en triglycérides
entreposés dans les adipocytes et autres tissus périphériques (3-4-5). Relevant du
1catabolisme, la lipolyse se décrit comme l’hydrolyse de ces triglycérides et conduit au
relargage systémique de ses produits, des acides gras non-estérifiés, éventuellement captés
par les tissus périphériques. C’est là qu’entre en jeu la β-oxydation mitochondriale, se
traduisant comme la dégradation intracellulaire d’acides gras en AC (5). En période de jeûne,
les réserves d’AC sont utilisées pour alimenter la synthèse de corps cétoniques (cétogenèse)
au foie. Les corps cétoniques servent de source d’énergie importante pour le cœur et les
muscles mais surtout pour le cerveau qui demeure non-accessible aux acides gras libres (6).
Les glucides sont une autre source de carburant métabolique. Les voies de biosynthèse et de
dégradation à travers lesquelles circulent le glucose et ses produits comprennent
premièrement la néoglucogenèse, c’est-à-dire la production de glucose à partir de substrats
non-glucidiques tels le pyruvate et le lactate, qui est majoritairement effectuée dans le foie
(7-8). Ce glucose peut ensuite circuler jusqu’aux tissus cibles où il sera capté par des
transporteurs membranaires spécifiques. Par exemple, GLUT1 permet l’absorption cellulaire
dans presque tous les tissus alors que GLUT2 est surtout exprimé au foie, au pancréas et dans
l’intestin. L’expression de ces deux transporteurs est régulée par les concentrations
plasmatiques de glucose. Régulé par l’insuline, GLUT4 permet au glucose de pénétrer les
cellules cardiaques, musculaires et les adipocytes (9-10-11). Une fois absorbé, le glucose est
dégradé en pyruvate ou en lactate pour servir de carburant ; c’est la glycolyse (7-8).
Deux sentiers sont intégrés au métabolisme du glycogène. D’abord, la glycogenèse est le
processus de conversion et du stockage du glucose sous forme de glycogène dans le foie et
dans les muscles. Le glycogène hépatique sert à réapprovisionner les tissus périphériques en
énergie lors d’un jeûne alors que dans les muscles squelettiques, le glycogène est utilisé lors
d’une activité physique intense. Le deuxième sentier est la glycogénolyse ou dégradation du
glycogène en glucose (12-13).
Au cœur des processus intégrés dans le contrôle du métabolisme énergétique opèrent
plusieurs hormones clés agissant de façon coordonnée (14-15). En voici quelques-unes :
2D’abord, le pancréas est le site de production d’insuline (16). Cette hormone anabolique
favorise la lipogenèse et l’absorption cellulaire du glucose tout en inhibant la production
hépatique de glucose (17). L’insuline agit principalement au foie (18), dans le muscle
squelettique et dans le tissu adipeux (19). Sécrétée par les cellules pancréatiques α, le
glucagon exerce une action antagoniste à l’insuline en stimulant la glycogénolyse, la
néoglucogenèse et la lipolyse (20).
La leptine est une hormone sécrétée majoritairement par les adipocytes blancs et codée par
le gène ob (21). Cette cytokine cible des récepteurs spécifiques codés par le gène db et
exprimés notamment dans le noyau arqué de l’hypothalamus et dans le foie. Aussi connue
sous le nom d’hormone de satiété, la leptine contrôle la prise alimentaire (22). En tant
qu’hormone catabolique, elle est considérée comme un inhibiteur de la lipogenèse et stimule
aussi la lipolyse (23).
L’adiponectine est une autre hormone sécrétée par le tissu adipeux et capable d’améliorer la
sensibilité à l’insuline (21). Ses principaux tissus cibles sont le foie et le muscle squelettique.
Dans ces deux tissus, l’action de l’adiponectine induit des effets sur le métabolisme du
glucose et des lipides ; diminution de la production hépatique de glucose contre une
augmentation de la captation du glucose et de l’oxydation des acides gras dans le muscle
(24). Des effets bénéfiques tels qu’une protection contre l’athérosclérose et l’inflammation
chronique lui sont également associés (25). De façon générale, un ratio adiponectine/leptine
élevé est indicateur d’une bonne santé métabolique (26).
L’insulin-like growth factor 1 (IGF-1) est une hormone catabolique sécrétée par le foie et
impliquée dans la croissance et la prolifération cellulaire. Elle favorise la différenciation
adipocytaire et le captage des acides gras par les muscles, aidant ainsi à réduire le stockage
hépatique de lipides (27). Elle accroît également la sensibilité à l’insuline (28).
L’hormone thyroïdienne triiodothyronine (T3) est aussi impliquée dans la coordination du
métabolisme des lipides. Ses effets connus incluent une synthèse accrue de récepteurs aux
LDL (LDL-R), une clairance améliorée du cholestérol et des triglycérides circulants. T3
3ralentit l’accumulation hépatique de lipides (stéatose hépatique) en favorisant l’oxydation
des acides gras mais diminue la sensibilité à l’insuline (29).
Les glucocorticoïdes occupent une position centrale dans la régulation de l’homéostasie. Ces
molécules stimulent la néoglucogenèse par le foie et la synthèse de glycogène. De plus, elles
modulent à la baisse l’absorption cellulaire du glucose. Les glucocorticoïdes diminuent la
sensibilité à l’insuline (30-31). Leurs actions dans la balance énergétique des lipides résultent
en une augmentation de la lipogenèse au foie et une mobilisation des acides gras dans le tissu
adipeux (32).
1.1.2 Le foie, centre intégrateur du métabolisme énergétique
L’équilibre entre le métabolisme des lipides et des glucides est sous le contrôle du foie de
par son rôle clé au cœur de la régulation fine de ces processus. Deux voies contrôlent les flux
de substrats lipidiques et glucidiques ; exogène et endogène (Fig. 2 – adaptée de (33)) (34).
En état postprandial, le corps reçoit un apport important de lipides alimentaires (voie
exogène). Ces lipides sont hydrolysés dans la lumière intestinale par l’action de sels biliaires
puis réestérifiés à l’intérieur de chylomicrons, qui font partie d’un groupe de complexes
moléculaires appelés lipoprotéines servant au transport des lipides. Suivant leur synthèse, les
chylomicrons migrent vers les tissus périphériques où leur contenu en triglycérides sont de
nouveau hydrolysés par la lipoprotéine lipase. Ces lipides sont éventuellement stockés sous
forme de triglycérides dans les cellules adipeuses où ils servent de réserves énergétiques. Les
chylomicrons résiduels migrent ensuite jusqu’au foie pour être captés et soit participer à la
synthèse de lipides complexes ou être stockés (35-36). La voie endogène peut se décrire par
une sécrétion hépatique de Very-low-density lipoproteins (VLDL), transporteurs des
triglycérides entreposés dans les hépatocytes, lors d’un jeûne. L’hydrolyse graduelle de ces
VLDL dans les tissus cibles altèrent leur composition ; ils se convertissent en Intermediate-
density lipoproteins (IDL) puis en Low-density lipoproteins (LDL), appauvris en
triglycérides. Le foie et l’intestin participent à la formation des High-density lipoproteins
(HDL), une autre classe de lipoprotéines servant au captage du cholestérol périphérique et à
promouvoir son recyclage au foie (34-37-38).
4Fig. 2: Voies du métabolisme des lipides (adaptée de (33)).
Les apports alimentaires déclenchent une cascade de signalisation hormonale par les cellules
pancréatiques β qui répondent par une sécrétion d’insuline. L’insuline régule d’abord à la
baisse la glycémie postprandiale en inhibant la sécrétion de glucagon (Fig. 3 – adaptée de
(38)). Les apports exogènes de glucose exacerbent d’autant plus cette inhibition et, de concert
avec l’insuline, inhibe la néoglucogénèse (endogène) par le foie (39). En phase de jeûne, la
sécrétion d’insuline est inhibée en faveur du glucagon et conséquemment de la dégradation
du glycogène et de la néoglucogenèse afin de réapprovisionner les cellules appauvries en
glucose (Fig. 3 – adaptée de (38)) (40-41). Cette production hépatique de glucose compense
5les faibles apports exogènes et engendre donc indirectement la lipolyse et l’oxydation des
acides gras pour le rétablissement d’un niveau basal de la glycémie.
Fig. 3: Régulation de la glycémie (adaptée de (38)).
1.1.3 Désordres fonctionnels au foie et pathologies associées
Le vieillissement est associé à des changements physiologiques importants tels qu’une
formation de graisses corporelles élevée, une sensibilité diminuée à l’insuline et une
redistribution des lipides à des sites ectopiques et viscéraux. La baisse de sensibilité à
l’insuline favorise la lipolyse dans le tissu adipeux ce qui élève les taux d’acides gras
circulants et stimule la stéatose hépatique. De plus, la sécrétion de cytokines pro-
inflammatoires augmente et est accompagnée d’une élimination diminuée de radicaux libres
(42-43). Ces dérèglements favorisent une inflammation chronique entre autres du tissu
6adipeux et du foie, altérant d’autant plus les fonctions physiologiques normales de ces
organes métaboliques (42-43).
Les déséquilibres d’ordre nutritionnel sont fortement liés à l’épidémie d’obésité qui frappe
depuis plusieurs années les populations des pays développés. Définie comme une
accumulation excessive de lipides, cette condition favorise l’apparition d’un éventail de
signes physiologiques reflétant un métabolisme altéré (44). Les gens atteints d’obésité
peuvent présenter une tolérance au glucose diminuée, des niveaux élevés de lipides sanguins
et des réponses inflammatoires anormales. D’autres troubles métaboliques typiques de
l’obésité incluent un stress oxydatif exacerbé, un débalancement dans l’équilibre des
adipokines sécrétées (une conséquence directe d’un tissu adipeux dysfonctionnel) et une
stéatose hépatique (45).
Plusieurs dérèglements du métabolisme associés au vieillissement (46) et à l’obésité (47)
possèdent comme cause sous-jacente la résistance à l’insuline. Celle-ci est notamment
associée à l’obésité abdominale, à l’hypertension artérielle, à une tolérance au glucose
diminuée et à la dyslipidémie. L’ensemble de ces symptômes, nommé syndrome métabolique
(MetS) (48), favorise l’apparition du diabète de type 2 et de maladies cardiovasculaires.
Le foie, organe central dans le maintien de l’homéostasie énergétique, est donc susceptible à
des déclins fonctionnels (insuffisance hépatocellulaire) dans le vieillissement et l’obésité.
L’engorgement de lipides intra-hépatocytaires associé à ces deux conditions pathologiques
constitue le premier stade de la stéatose hépatique non alcoolique (NAFLD) (43). Dans ce
contexte physiologique, une série de réseaux régulateurs interconnectés composés de cellules
immunitaires est déclenchée et mène à l’infiltration de macrophages et de cytokines pro-
inflammatoires dans le tissu hépatique. La combinaison de la stéatose et de l’activation
chronique de ces voies inflammatoires conduit au stade suivant de la maladie; la
stéatohépatite non alcoolique (NASH) (Fig. 4 – adaptée de (49)). Cet état sévère de la
pathologie est caractérisé par de l’inflammation lobulaire et par un stress oxydatif important
induisant des lésions au foie. Les mécanismes de cicatrisation sont alors activés en réponse
à ces lésions tissulaires, expliquant pourquoi la fibrose est généralement présente lors du
7NASH (50). Ces tissus cicatriciels perturbent la circulation du sang dans l’organe et
contribuent à l’apparition de la cirrhose, stade ultime de la maladie, qui altère de façon
importante les fonctions hépatiques normales (51). Il y a risque d’infection dans l’abdomen,
de carcinomes hépatocellulaires et la transplantation hépatique doit être considérée.
Fig. 4: Progression de la maladie hépatique (adaptée de (49)).
En résumé, le vieillissement et l’obésité mènent à un état d’insulinorésistance favorisant une
élévation du taux de glucose sanguin et une hyperlipidémie par une lipolyse mal contrôlée
dans le tissu adipeux. Une des conséquences physiologiques directes de ce processus est
l’accumulation excessive de lipides au foie. La stéatose diminue la sensibilité des hépatocytes
à l’action de l’insuline, consolidant d’autant plus des troubles métaboliques, et peut
provoquer la maladie hépatique.
Le vieillissement et l’obésité comportent donc un chevauchement dans la portée de leurs
effets biologiques et des mécanismes sous-jacents. Des distinctions phénotypiques entre ces
deux situations physiologiques sont toutefois rapportées dans la littérature, notamment dans
une étude visant à cerner la contribution différentielle entre le vieillissement et une
alimentation riche en gras (HFD) dans le développement de l’insulinorésistance (52). Les
données physiologiques tirées du paradigme établi par les auteurs, comprenant des souris
jeunes ou vieilles soumises à une diète faible en gras (LFD) ou au HFD, suggèrent que le
vieillissement affecterait davantage la tolérance au glucose que la sensibilité à l’insuline. De
plus, l’expression au foie des gènes liés au métabolisme du glucose serait plus susceptible à
des variations lors du vieillissement que dans l’obésité, dont les effets dans la régulation
8transcriptionnelle seraient plus importants pour les gènes propres à l’homéostasie des lipides
(52). Une étude plus récente a évalué les impacts du vieillissement et de l’obésité dans une
dynamique englobant le métabolisme et les réponses immunitaires (53). Des groupes de
souris jeunes et minces ou spontanément obèses ont été comparées à des groupes de souris
âgées et minces ou nourries au HFD. Les animaux caractérisés par l’insulinorésistance la plus
importante et le plus haut degré de stéatose hépatique étaient ceux du modèle d’obésité
nutritionnelle. De plus, les foies de ces souris étaient les plus ciblés en terme d’infiltration de
macrophages (53). Finalement, une autre étude chez la souris intégrant l’évaluation des effets
physiologiques du HFD et du vieillissement montre que l’avancement en âge génère aussi
des dérèglements du métabolisme similaires à ceux induits par l’obésité (54). Dans tous les
cas, le vieillissement et l’obésité altèrent l’équilibre énergétique régissant les voies de
biosynthèse et de dégradation de biomolécules et provoquent des changements qui reflètent
une diversité de troubles métaboliques. Bien qu’il soit possible que les effets de ces deux
situations pathologiques soit au moins partiellement médié par des processus différents et
que l’obésité apporte les conséquences les plus sévères, il est fort probable que le
vieillissement soit à la base d’effets synergiques sur celles-ci. Comme le stress métabolique
engendré par l’avancement en âge se déroule plus passivement que dans l’obésité, il se peut
que l’organisme puisse s’y adapter plus facilement, tamponnant ainsi ses contrecoups.
1.2 ACTEURS ET MÉCANISMES MOLÉCULAIRES
1.2.1 Facteurs de transcription
Les mécanismes moléculaires qui régulent l’expression des enzymes, des transporteurs et des
récepteurs impliqués dans le métabolisme énergétique sont donc des éléments clés de la
recherche centrée sur les processus et pathologies discutés ci-haut. Au cœur de ces
mécanismes interagissent une diversité d’acteurs comme les facteurs de transcription (TFs)
qui se fixent aux séquences activatrices de gènes cibles pour stimuler leur expression (55).
Les Sterol regulatory element binding proteins (SREBPs) sont des facteurs de transcription
qui modulent la lipogenèse et qui régulent l'homéostasie du cholestérol. Leur famille est
constituée de SREBP1a et SREBP1c (des isoformes) ainsi que de SREBP2 qui est transcrit
9d’un gène à part (SREBF2) (56). SREBP s’exprime initialement sous forme d’un précurseur
maintenu au réticulum endosplasmique où il est physiquement lié avec SREBP Cleavage-
Activating Protein (SCAP). Le déclenchement de la maturation de SREBP est dépendante
des niveaux intracellulaires de cholestérol. Lorsque ceux-ci sont appauvris, le complexe
hétérodimérique SREBP-SCAP est relargué du réticulum endoplasmique vers l’appareil de
Golgi (GA) par dissociation de SCAP, doté d’un domaine de captage des stérols, d’avec les
protéines Insulin Induced Gene 1 (INSIG1) et Insulin Induced Gene 2 (INSIG2). Au GA, les
protéines Site-1P (S1P) et Site-2P (S2P) exercent une action protéolytique à deux sites
spécifiques du précurseur de SREBP pour en libérer un facteur de transcription possédant
des domaines hélice-boucle-hélice et glissière à leucine (bHLH-Zip). SREBP mature migre
ensuite au noyau et se fixe à un Sterol Response Elements (SRE) situé dans les régions
promotrices de gènes lipogéniques (Figure 5 - (57)) (58-59). Ce facteur de transcription est
régulé au niveau transcriptionnel et dans son activation post-traductionnelle (accumulation
nucléaire) par des mécanismes intégrant la signalisation de l'insuline (59). En somme,
SREBP contrôle efficacement le métabolisme des lipides, stimulant la lipogenèse lorsque
nécessaire pour préserver l'équilibre dans les niveaux intracellulaires de cholestérol.
Fig. 5: Processus de maturation du facteur de transcription SREBP (57).
10Les Forkhead box O (FOXO) sont une autre famille de facteurs de transcription importante
pour le métabolisme et comprenant 4 membres ; FOXO1, FOXO3, FOXO4 ainsi que
FOXO6. Parmi ceux-ci, le facteur de transcription FOXO1 est bien caractérisé dans la
littérature comme étant un régulateur central des gènes néoglucogéniques, une fonction
régulée par plusieurs voies de phosphorylation (60). En période postprandiale, l’insuline
inhibe FOXO1 par phosphorylation. Une fois inactivé, FOXO1 est expulsé du noyau et ne
peut plus exercer son action transcriptionnelle, ce qui ralenti significativement la
néoglucogénèse. Lors d’un jeûne, la glycémie diminue et entraîne une baisse des niveaux
circulants d’insuline. Cet évènement permet l’activation de FOXO1 qui migre alors au noyau
pour potentialiser la transcription de ses gènes cibles (60-61). FOXO1 participe donc au
maintien de l’homéostasie en stimulant la production de glucose par le foie.
En résumé, l’importance de l’insuline dans le métabolisme énergétique se reflète notamment
à travers sa modulation de l’activité de régulateurs transcriptionnels tels que SREBP et
FOXO1 et illustre bien les liens établis entre différentes adaptations physiologiques et les
mécanismes moléculaires sous-jacents.
1.2.2 Récepteurs nucléaires
Les récepteurs nucléaires sont une superfamille de protéines régulatrices de grande
importance dans le contrôle des mécanismes transcriptionnels chez les eucaryotes. Chez
l’humain, 48 gènes codant pour des récepteurs nucléaires sont répertoriés (62). Ils sont
impliqués entre autres dans la différenciation et la croissance cellulaire, la morphogenèse, la
reproduction et le métabolisme (63-64). Les récepteurs nucléaires se distinguent des autres
familles de facteurs transcriptionnels par leur mode d’activation ; ils lient directement des
ligands spécifiques dans le cytoplasme puis migrent au noyau. Ces ligands comprennent
notamment des hormones stéroïdiennes telles que les glucocorticoïdes et les
minéralocorticoïdes mais aussi des stéroïdes sexuels comme les œstrogènes et les androgènes
(65-66). Suivant la translocation nucléaire, ils peuvent réguler l’expression de divers gènes
en réponse au signal initial pour stimuler les adaptations physiologiques appropriées (64).
11Les Peroxisome proliferator-activated receptors (PPARs) sont des récepteurs nucléaires qui
comprennent les sous-types PPARα, PPARγ et PPARδ (67). Ceux-ci sont impliqués dans
divers processus biologiques comme le métabolisme des acides gras et des glucides mais
aussi dans le contrôle de la voie de l’insuline et la modulation de réponses inflammatoires .
Ils se distinguent par leurs rôles physiologiques (toutefois caractérisés par un certain
chevauchement) et partiellement par leur distribution tissulaire (Figure 6 – adaptée de (68))
(69).
En période de jeûne, PPARα (dominant au foie et principalement activé par liaison avec des
acides gras) favorise la β-oxydation par les mitochondries et les peroxysomes (70). Ce
récepteur nucléaire soutient la diminution des taux circulants des triglycérides en agissant
comme un répresseur transcriptionnel de l’apolipoprotéine C-III (Apo-CIII), qui elle est
capable d’inhiber l’enzyme lipolytique lipoprotéine lipase (LPL) (70-71). PPARγ est plus
spécifiquement exprimé dans le tissu adipeux et est un maître régulateur de la prolifération
et de la différenciation adipocytaire et des évènements lipogéniques. Un de ses ligands
naturels les plus connus est le 15dPGJ2, un dérivé des prostaglandines (72). En plus de
réguler l’expression d’adipokines telles que l’adiponectine et la leptine, l’activation de
PPARγ est fortement associée à une sensibilité accrue à l’insuline (72-73-74). PPARδ, le
troisième membre de la famille, est exprimé entre autres dans le tissu adipeux, dans des
cellules immunitaires comme les macrophages et dans les muscles squelettiques. Ce
récepteur nucléaire lie notamment des acides gras provenant de particules VLDL captées par
les macrophages (75). Son rôle dans la suppression des processus inflammatoires est bien
connue et en fait une cible thérapeutique potentielle d’importance contre l’athérosclérose
(75-76).
12Fig. 6: Fonctions des PPARs (adaptée de (68)).
Les Liver X receptors (LXRs) sont des récepteurs nucléaires activés entre autres lorsqu’ils
lient des oxystérols (77). Ils contrôlent la dynamique du transport inverse du cholestérol
(recyclage hépatique) entre autres par l’activation du transporteur ATP Binding Cassette A
1 (ABCA1) (77). De plus, LXR est un facteur transcriptionnel important pour la transcription
du gène SREBP-1. Il est intégré à un mécanisme clé de la régulation insulino-dépendante de
SREBP-1c de par sa liaison à une séquence d'élément de réponse LXR (LXRE) dans le
promoteur SREBP-1 (78-79). Ainsi, LXR favorise simultanément les processus lipogéniques
et l'élimination des excès de cholestérol dans la circulation. Les acides biliaires servent de
ligands pour les Farsenoid X receptors (FXRs). Une fois activés, ces récepteurs nucléaires
régulent finement les niveaux de cholestérol en inhibant l’activité de la Cholesterol 7 alpha-
hydroxylase (CYP7A1), une enzyme essentielle à la conversion du cholestérol en acides
biliaires. Les FXRs exercent aussi un contrôle sur la sécrétion des lipoprotéines (80-81).
Les PPARs, LXR et FXR s’hétérodimérisent avec le Retinoid X receptor (RXR) en formant
un complexe régulateur nécessaire à l’activation de la transcription de leurs gènes cibles (82).
13Enfin, le récepteur Hepatocyte Nuclear Factor 4 alpha (HNF4α) permet l’activation de gènes
clés de l’homéostasie des lipides tels que Microsomal Triglyceride Transfer Protein (MTTP)
et Apolipoprotein B (ApoB) et potentialise la différenciation des hépatocytes (83).
Les récepteurs nucléaires peuvent aussi être activés par l’intermédiaire de médicaments. Par
exemple, les fibrates sont des acides carboxyliques qui servent d’agonistes pour PPARα et
leur usage est prescrit chez les patients atteints d’hypertriglycéridémie (84). D’autre part, la
pioglitazone est un ligand synthétique de PPARγ et est utilisée chez les patients diabétiques
comme agent hypoglycémiant (85).
1.2.3 Corégulateurs
Les corégulateurs sont des protéines nucléaires capables de stimuler (coactivateurs) et de
réprimer (corépresseurs) la transcription génique chez les eucaryotes. Les stratégies
moléculaires utilisées par les corégulateurs comprennent premièrement le remodelage
structural de la chromatine (86-87). À l’état basal, celle-ci est repliée autour de complexes
protéiques appelés histones. Elle est maintenue sous forme empaquetée via des interactions
entre les charges acide désoxyribonucléique (ADN) négatives et les résidus lysines des
histones chargés positivement (88). Les corégulateurs peuvent acétyler ces résidus et donc
inhiber ces forces électrostatiques ; il en résulte un déroulement de la chromatine qui devient
accessible aux facteurs régulateurs qui se positionnent aux activateurs (ou répresseurs) de
leur gènes cibles (86-87). La deuxième fonction des corégulateurs consiste à induire le
recrutement de ces facteurs de transcription jusqu’aux promoteurs proximaux des sites
d’initiation de la transcription. Il s’ensuit la formation du complexe de pré-initiation de la
transcription qui s’effectue par un recrutement de l’acide ribonucléique (ARN) Polymérase
II (ARN Pol II) et d’autres facteurs composant cet assemblage (89-90). Finalement, les
cofacteurs transcriptionnels peuvent initier la transcription en servant de pont physique de la
signalisation entre les TFs fixés aux régions promotrices et la machinerie générale de
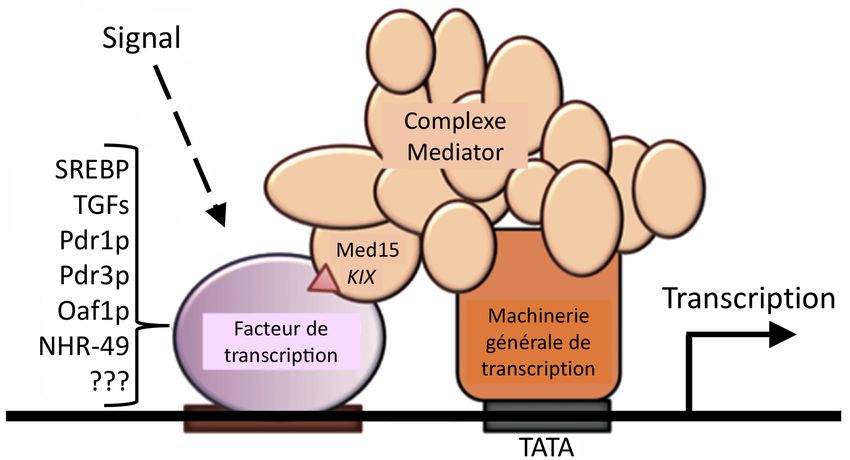
transcription (89-90). Un cofacteur nucléaire fondamental à la régulation transcriptionnel du
génome du domaine des eucaryotes et qui intègre toutes les fonctions décrites ci-haut est le
complexe Mediator.
141.3 COMPLEXE MEDIATOR
1.3.1 Structure et versatilité fonctionnelle
Mediator est le grand architecte moléculaire gouvernant la transcription des eucaryotes (91-
92-93-94). Chez les mammifères, ce corégulateur très conservé dans l’évolution est composé
de 26 sous-unités réparties en 4 modules ; Tête, Centre, Queue et CDK (Fig. 7 – adaptée de
(95)) (96). Mediator est doté d’une versatilité fonctionnelle et structurale; ses différentes
sous-unités modulent différents gènes et voies de signalisation (95-97). Ce trait lui permet
d’interagir dans un contexte spatio-temporel distinct avec plusieurs TFs de différentes
familles, un concept dont la compréhension demeure toutefois incomplète à ce jour.
Fig. 7: Architecture du corégulateur transcriptionnel Mediator (adaptée de (95)).
Les modules eux-mêmes ont des fonctions spécifiques. Par exemple, le module de Tête est
essentiel à la formation du complexe de pré-initiation et sert de plate-forme de liaison à
15l’ARN Pol II et aux facteurs généraux de transcription (98-99). Le module du Milieu participe
aussi à cet assemblage et en renforce la stabilisation (100). Le module de Queue coordonne
le recrutement aux facteurs transcriptionnels liés aux séquences activatrices en servant de
points d’attache directe. Les effets de répression transcriptionnelle par Mediator nécessite le
module malléable CDK, capable de s’en détacher (94-101).
1.3.2 Fonctions primaires et régulatrices
Mediator stimule le remodelage de la chromatine en jouant un rôle dans le recrutement des
histones acétyles transférases et donc augmente l’accessibilité des gènes aux TFs et à la
machinerie basale transcriptionnelle (102). Ce corégulateur permet aussi la formation du
complexe de pré-initiation (PIC) de la transcription et permet le recrutement de l’ARN Pol
II (102). De plus, il sert de plateforme de transmission de signaux entre le TF lié avec son
élément de réponse et les facteurs généraux de transcription (GTFs) (93). En somme,
Mediator est un centre régulateur de la transcription génique.
1.3.3 Importance métabolique
Les sous-unités discutées ici-bas ont été choisies de par la nature des fonctions moléculaires
et du positionnement au sein du complexe Mediator qui les caractérisent : elles font toutes
parties d’un module différent. De façon intéressante, les différentes sous-unités qui
composent ce complexe universel contrôlent de façon sélective le métabolisme (103).
• Med1
Le cofacteur Med1, sous-unité du module du Milieu (Fig. 7 – adaptée de (95)), est une
protéine régulatrice centrale dans le recrutement de divers facteurs de transcription et de
récepteurs nucléaires clés du métabolisme (104-105). Dans des hépatocytes, l’expression de
gènes cibles de PPARα est au moins partiellement dépendante de l’activité de Med1 (106-
107-108). De plus, il a été montré que la délétion génétique de ce cofacteur engendre des
déficiences transcriptionnelles dans la prolifération hépatocytaire (108-109), dans la β-
oxydation (106) et qu’il interagi physiquement avec PPARα (110). Ces multiples évidences
16indiquent que Med1 semble être un partenaire fonctionnel clé de PPARα dont il régulerait
les rôles. Med1 peut interagir avec PPARγ dans la régulation de l’adipogenèse (111-112), et
joue aussi un rôle d’importance dans la coordination du métabolisme du glucose et des voies
de l’insuline (113). D’autres études ont rapporté le caractère essentiel de Med1 dans le
développement physiologique sain du cœur et la coordination transcriptionnelle de gènes
essentiels aux fonctions cardiaques chez le mammifère (114-115).
• Med12
Les rôles de Mediator dans le contrôle de la transcription génique se reflètent aussi dans
l’acheminement de signaux intégrés dans diverses cascades de signalisation telles que la voie
canonique Wnt (92). Cette voie est initiée avec la liaison des protéines Wnt à leurs récepteurs
membranaires spécifiques qui entraîne leur activation d’où découle une accumulation
cytoplasmique de protéines β-caténines (116). Les β-caténines migrent ensuite au noyau pour
induire le recrutement de cofacteurs nucléaires et déclencher la transcription de gènes
spécifiques à des processus tels que la prolifération cellulaire et le développement
embryonnaire (117). Une étude a caractérisé Med12, intégré dans le module CDK (Fig. 7 –
adaptée de (95)), en tant que partenaire des β-caténines et révélé que leur association est
nécessaire pour stimuler son recrutement aux promoteurs de gènes impliqués dans la voie
Wnt. L’invalidation génétique de Med12 induit une suppression de la carcinogenèse au moins
partiellement médiée par une inhibition de la voie Wnt (118), qualifiant Med12 comme cible
thérapeutique potentielle contre le cancer.
• Med14
Située dans le module de Queue (Fig. 7 – adaptée de (95)), la protéine Med14 sert de
régulateur transcriptionnel important dans les processus physiologiques sous le contrôle de
PPARγ mais indépendant de l’expression de Med1 et de la liaison PPARγ-ligand (119). En
effet, une étude poussée des rôles moléculaires de Med14 a révélé que cette sous-unité
favorise la transcription de gènes impliqués dans l’accumulation de lipides tout en stimulant
l’adipogenèse dans des cellules 3T3-L1, qu’elle stimule le recrutement de Mediator aux
17Vous pouvez aussi lire

















































