Réseau trophique de la tordeuse des bourgeons de l'épinette dans différents habitats et contextes - Mémoire Valentine Glaus Maîtrise en biologie ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Réseau trophique de la tordeuse des bourgeons de
l'épinette dans différents habitats et contextes
Mémoire
Valentine Glaus
Maîtrise en biologie végétale - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Valentine Glaus, 2022
Réseau trophique de la tordeuse des bourgeons de
l'épinette dans différents habitats et contextes
Mémoire
Valentine Glaus
Sous la direction de :
Valérie Fournier, directrice de recherche
Véronique Martel, codirectrice de recherche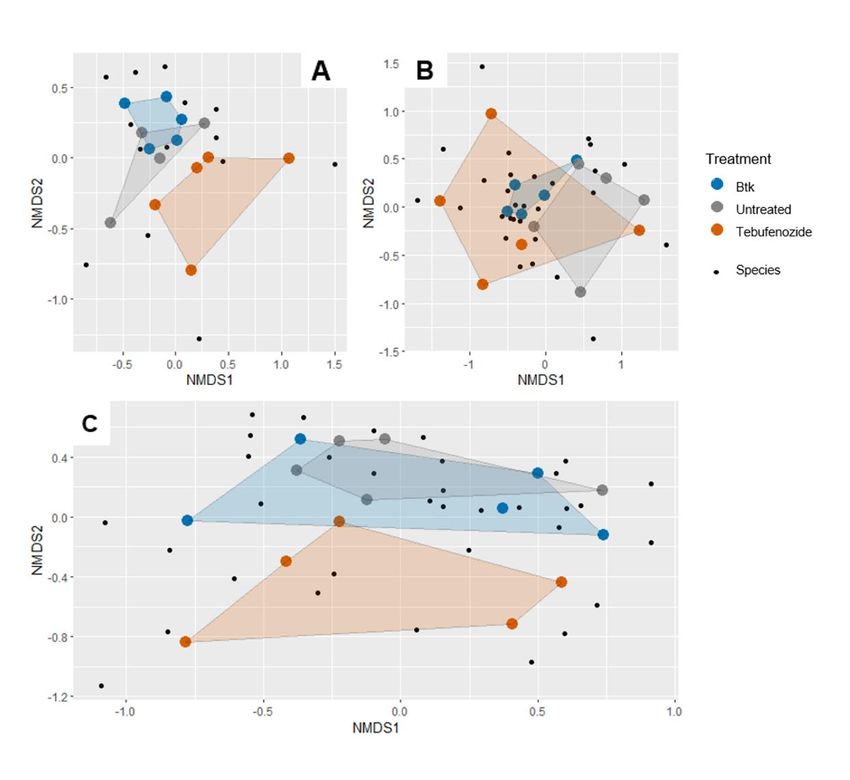
Résumé
La tordeuse des bourgeons de l’épinette (Choristoneura fumiferana Clemens;
Lepidoptera : Tortricidae; TBE), qui s’attaque au sapin baumier (Abies balsamea (L.)
Mill.) et aux épinettes (Picea spp. Mill.), est le défoliateur le plus important des forêts
boréales d’Amérique du Nord. Son réseau trophique comporte plus d’une centaine
de parasitoïdes qui offrent un contrôle naturel des populations et la compréhension
de celui-ci dans différents contextes est essentiel. Le premier objectif du projet était
d’améliorer la compréhension du réseau trophique de la TBE entre deux épidémies,
peu étudié alors qu’il permettrait de mieux comprendre les processus du
déclenchement d’une épidémie. Une meilleure connaissance du réseau permettra
d’identifier les effets potentiels des interventions de lutte. Un contrôle des
populations de TBE visant à prévenir les épidémies est à l’étude au Nouveau-
Brunswick depuis 2014. Malgré la spécificité élevée des insecticides utilisés, les
traitements pourraient avoir des impacts sur d’autres espèces de chenilles qui sont
des hôtes alternatifs aux parasitoïdes, mettant en péril le contrôle naturel lors des
années suivantes. Le deuxième objectif était de définir si ces traitements ont des
impacts non ciblés en étudiant la communauté de chenilles ainsi que son taux de
parasitisme. L’analyse des 1 978 chenilles récoltées aux sites non-épidémiques a
permis d’identifier 19 nouvelles associations entre des parasitoïdes de la TBE et des
hôtes alternatifs, améliorant ainsi la compréhension du réseau trophique de la TBE
à ce stade. L’analyse moléculaire des 659 chenilles récoltées au Nouveau-
Brunswick suggère que les traitements de la stratégie d’intervention hâtive n’ont
aucun effet négatif, que ce soit sur l’abondance, la richesse de chenille ou le taux
de parasitisme. Cette étude révèle donc que les traitements ciblant la TBE, effectués
dans ces conditions, n’affectent pas négativement la communauté de chenilles,
suggérant que les parasitoïdes ne manqueront pas d’hôtes alternatifs.
iiAbstract
The spruce budworm (Choristoneura fumiferana Clemens; Lepidoptera: Tortricidae;
SBW), which attacks balsam fir (Abies balsamea (L.) Mill.) and spruce (Picea spp.
Mill.), is the most important defoliator of North American boreal forests. Its food web
includes more than 100 parasitoids that provide natural population control and
understanding this in different contexts is essential. The first objective of the project
was to improve the understanding of the food web of SBW between two epidemics,
which has been little studied and which would allow a better understanding of the
processes of epidemic initiation. A better understanding of the food web will help
identify the potential effects of control interventions. Control of SBW populations to
prevent outbreaks has been under investigation in New Brunswick since 2014.
Despite the high specificity of the insecticides used, the treatments could have
impacts on other caterpillar species that are alternative hosts to the parasitoids,
jeopardizing natural control in subsequent years. The second objective was to define
if these treatments have non-target impacts by studying the caterpillar community as
well as its parasitism rate. Analysis of the 1,978 caterpillars collected at the non-
epidemic sites identified 19 new associations between SBW parasitoids and
alternate hosts, improving the understanding of the SBW food web at this stage.
Molecular analysis of 659 caterpillars collected in New Brunswick suggested that the
early intervention strategy treatments had no negative effect on either abundance,
caterpillar richness or parasitism rate. Therefore, this study shows that treatments
targeting SBW under these conditions do not negatively affect the caterpillar
community, suggesting that parasitoids will not lack alternative hosts.
iiiTable des matières
Résumé .................................................................................................................... ii
Abstract ................................................................................................................... iii
Table des matières .................................................................................................. iv
Liste des figures ...................................................................................................... vi
Liste des tableaux .................................................................................................. vii
Liste des abréviations ............................................................................................ viii
Remerciements ....................................................................................................... ix
Avant-propos ........................................................................................................... x
INTRODUCTION GÉNÉRALE ................................................................................ 1
ÉTAT DES CONNAISSANCES ............................................................................... 3
Écosystèmes forestiers et perturbations .............................................................. 3
La tordeuse des bourgeons de l’épinette ............................................................. 6
Le réseau trophique de la TBE ............................................................................ 8
Parasitoïdes ...................................................................................................... 8
Hôtes alternatifs .............................................................................................. 11
Autres ennemis naturels ................................................................................. 13
Dynamique de population .................................................................................. 14
Lutte contre la TBE ............................................................................................ 16
Protection du feuillage .................................................................................... 18
Contrôle de population .................................................................................... 18
Impacts non ciblés des traitements ................................................................. 21
Problématique .................................................................................................... 25
Objectifs et hypothèses ...................................................................................... 26
Méthodologie ..................................................................................................... 26
Échantillonnage .............................................................................................. 26
Méthode d’analyse .......................................................................................... 27
CHAPITRE 1: DESCRIPTION OF THE SPRUCE BUDWORM NETWORK AT A
NON EPIDEMIC STAGE: THE IMPORTANCE OF PLANT DIVERSITY ............... 29
1.1 Résumé ........................................................................................................ 30
1.2 Abstract ........................................................................................................ 31
1.3 Introduction .................................................................................................. 32
1.4 Material and methods ................................................................................... 33
1.5 Results ......................................................................................................... 36
iv1.6 Discussion ................................................................................................ 38
CHAPITRE 2: EVALUATION OF THE NON-TARGET IMPACTS OF AN
INSECTICIDE-BASED POPULATION CONTROL FOR EASTERN SPRUCE
BUDWORM (LEPIDOPTERA: TORTRICIDAE) .................................................... 41
2.1 Résumé ........................................................................................................ 42
2.2 Abstract ........................................................................................................ 43
2.3 Introduction .................................................................................................. 44
2.4 Material and methods ................................................................................... 46
2.5 Results ......................................................................................................... 52
2.6 Discussion .................................................................................................... 59
2.7 Conclusion ................................................................................................... 62
CONCLUSION GÉNÉRALE .................................................................................. 64
Bibliographie.......................................................................................................... 68
Annexe 1 ............................................................................................................... 82
Annexe 2 ............................................................................................................... 83
Annexe 3 ............................................................................................................... 85
vListe des figures
Figure 1 : Superficie perturbée annuellement d’intensité modérée ou grave au
Canada entre 1990 et 2019 (National Forestry Database 2022a, 2022b, 2022c) ... 5
Figure 2 : Superficie perturbée d’intensité modérée ou grave par les principaux
insectes forestiers au Canada entre 1990 et 2019 (National Forestry Database
2022c). .................................................................................................................... 5
Figure 3 : Tordeuse des bourgeons de l’épinette (Choristoneura fumiferana
Clemens) au sixième stade larvaire et adulte. Source : Ressources naturelles
Canada. ................................................................................................................... 7
Figure 4: Non epidemic sites sampled in the province of Quebec in 2020 ........... 34
Figure 5: Trophic network of the spruce budworm parasitoids, their alternative hosts
and the plant hosts in two non epidemic sites in the province of Québec, Canada.
The width of the boxes and links represent the frequency of the species or
interaction. The dark gray links between the parasitoids and caterpillars represent
new interactions, not mentioned in the literature. See Table 1 for the names of
species. ................................................................................................................. 37
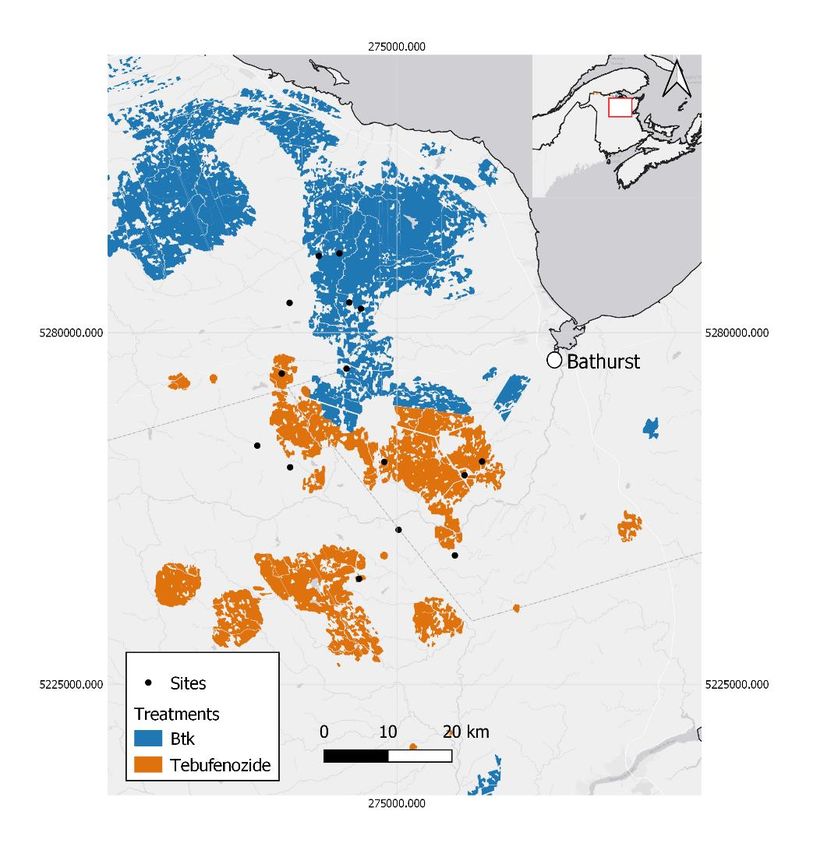
Figure 6: Sites sampled in New Brunswick the year of the insecticide treatments
(2018) (Btk, tebufenozide or control) and one year post-treatment (2019). ........... 48
Figure 7: Number of non-target caterpillars, richness (number of species) and
parasitism rates per treatments in New Brunswick. Insecticides were applied in 2018
only. Different letters indicate significant differences from post-hoc test applied when
an effect o treatment was detected, and parasitism rates were analyzed per year.
.............................................................................................................................. 53
Figure 8: Caterpillar’s richness, abundance and parasitism rate for each sampling
month, from May to September, in 2018, the year of the treatment, and in 2019, the
year following the treatment. ................................................................................. 55
Figure 9: Ordination plot (NMDS, k=2) with Bray-Curtis dissimilarity matrix on the
caterpillar community A: Early-season the year of the treatment (stress value: 0. 117)
B: Late-season the year of treatment (stress value: 0.100) C: One year post
treatment (stress value: 0.176). ............................................................................. 58
viListe des tableaux
Table 1: Species of parasitoids, caterpillars and plant found in the trophic network.
.............................................................................................................................. 38
Table 2: Sampling dates for the year of insecticide treatments of Btk and
tebufenozide (2018) and one year following the treatment (2019). ....................... 49
Table 3: Statistical values from the GLMMs explaining the caterpillar species
richness (number of species), abundance and the associated parasitism rate, for
both years combined and separated. .................................................................... 54
Table 4: Results of the pairwise PERMANOVA (999 permutations of the data)
conducted on the composition of the caterpillar community between each type of
treatment. .............................................................................................................. 56
viiListe des abréviations
Bt : Bacillus thuringiensis
Btk : Bacillus thuringiensis variété kurstaki
EIS : Early Intervention Strategy
TBE : Tordeuse des bourgeons de l’épinette
SBW : Spruce budworm
SIH : Stratégie d’intervention hâtive
viiiRemerciements
Tout d’abord un gros merci à Véronique Martel de m’avoir donné cette opportunité
de maitrise et de m’avoir embarqué dans de multiples projets avec d’innombrables
présentations. Tu m’as permis de me dépasser et j’en suis reconnaissante. Merci
pour ta confiance, tes encouragements, tes conseils et tes multiples relectures de
documents. Merci à Valérie Fournier de m’avoir accueilli dans ton laboratoire, de
m’avoir accompagné tout au long de ce projet et d’avoir veillé, avec Frédéric, au bon
déroulement du projet.
Sébastien, Simon, Jessica, Paule, Laurence, Louis-Philippe et Julien, merci pour
votre aide sur le terrain et au laboratoire, merci aussi pour votre bonne humeur
quotidienne. Je suis reconnaissante d’avoir pu réaliser ma maitrise dans votre
laboratoire. Sébastien, toujours en mission, ce projet n’aurait pas pu être possible
sans toi, merci tellement! Simon et Sébastien, toujours le mot pour me faire rire.
Merci pour votre aide, vous m’avez tout appris. Jess, merci pour tous les beaux
moments passés au bureau qui ont apporté beaucoup de légèreté. Merci Paule pour
ta présence rassurante et ta bienveillance. Audrey, qui m’a formé en biomoléculaire
et qui a répondu à mes multiples questions, merci pour ta patience et tes bons
conseils. Merci à Michel Cusson, et tous les membres de son laboratoire, Audrey,
Madjid, Catherine, Sandrine, de m’avoir accueilli pour réaliser mes analyses.
Merci à toute la famille pour votre support inconditionnel et de croire autant en moi,
vous êtes trop bien. Merci à tout.es mes ami.es, qui ont été là pour me soutenir, me
changer les idées et m’écouter quand j’en avais besoin, vous êtes les meilleur.es
merci tellement. Merci, Félix de m’avoir fait rire, merci pour ton écoute et tes
encouragements infinis. Ta présence a embelli cette aventure.
ixAvant-propos
Ce mémoire de maitrise est constitué d’une introduction générale au sujet, de deux
chapitres sous forme d’articles scientifiques et d’une conclusion générale.
L’introduction et la conclusion ont été rédigées en français alors que les deux articles
ont été rédigés en anglais par l’étudiante.
Le chapitre 1 « Description of the spruce budworm (Choristoneura fumiferana)
network at a non epidemic stage: the importance of plants diversity” en collaboration
avec Audrey Nisole, Valérie Fournier et Véronique Martel, est destiné à être soumis
dans la revue Canadian Journal of Forest Research d’ici 2023, sous la forme d’une
note scientifique. L’élaboration du protocole, la collecte des échantillons, les
analyses en laboratoire, l’interprétation des résultats et la rédaction du manuscrit ont
été réalisées par l’étudiante. Le financement provient de Ressources naturelles
Canada, du Fonds de recherche du Québec – Nature et technologies (FRQNT) et
de la bourse Wladimir-A.-Smirnoff, attribuée par Ressources Naturelles Canada, la
Société d’entomologie du Québec et la Société de protection des plantes du
Québec.
Le chapitre 2 « Evaluation of the non-target impacts of an insecticide-based
population control for eastern spruce budworm (Lepidoptera: Tortricidae) » en
collaboration avec Audrey Nisole, Sara Edwards, Sébastien Bélanger, Robert C.
Johns, Abdelmadjid Djoumad, Michel Cusson, Valérie Fournier et Véronique Martel
a été soumis à la revue The Canadian Entomologist le 9 mai 2022. L’article a été
accepté le 20 octobre 2022. La collecte des échantillons, les analyses moléculaires,
les analyses statistiques, l’interprétation des résultats ainsi que la rédaction du
manuscrit ont été effectuées par l’étudiante, avec l’aide de biologistes du Service
canadien des forêts. Le financement provient de Ressources naturelles Canada, du
Fonds de recherche du Québec – Nature et technologies (FRQNT) et de la bourse
Wladimir-A.-Smirnoff, attribuée par Ressources Naturelles Canada, la Société
d’entomologie du Québec et la Société de protection des plantes du Québec.
xINTRODUCTION GÉNÉRALE
La tordeuse des bourgeons de l’épinette (Choristoneura fumiferana Clemens;
Lepidoptera : Tortricidae; TBE), qui s’attaque au sapin baumier (Abies balsamea (L.)
Mill.) et aux épinettes (Picea sp. Mill.) (MacLean 1984), est le défoliateur le plus
important des forêts boréales d’Amérique du Nord (MacLean 1980). Elle a des
conséquences économiques (Chang et al. 2012), des impacts sur le taux de
croissance des arbres (MacLean 2016), sur les écoservices (Schowalter 2012) et
sur la communauté de lépidoptères (Scriber 2004). Son réseau trophique comporte
plus d’une centaine de parasitoïdes (Eveleigh et al. 2007) qui offrent un contrôle
naturel des populations (Régnière et Nealis 2007). Ce réseau change en fonction du
stade de l’épidémie (Eveleigh et al. 2007) : la guilde de parasitoïdes au stade
épidémique n’est pas la même qu’au déclin de l’épidémie. Son réseau trophique a
peu été étudié à un stade non-épidémique malgré l’importance de celui-ci.
Effectivement, une meilleure connaissance de ce stade permettrait de mieux
comprendre les processus du déclenchement d’une épidémie. La stratégie
d'intervention hâtive contre la TBE, qui vise à prévenir les épidémies, est à l’étude
au Nouveau-Brunswick depuis 2014 (Johns et al. 2019). Malgré la spécificité élevée
des insecticides utilisés (Bacillus thuringiensis var. kurstaki (Btk) et tébufénozide),
les traitements pourraient avoir des impacts sur les chenilles d’autres espèces
présentes lors de ceux-ci, qui sont des hôtes alternatifs aux parasitoïdes. Un impact
sur la communauté de chenilles pourrait perturber la communauté de parasitoïdes
qui est nécessaire au contrôle de la TBE de l’épinette les années qui suivent le
traitement. Une meilleure compréhension des impacts des traitements permettra
d’ajuster les protocoles afin d’en minimiser les effets non-ciblés.
Cette étude comportait deux objectifs ayant en commun le réseau trophique de la
TBE : améliorer la compréhension de son réseau trophique à un stade non-
épidémique et définir si les traitements de la stratégie d’intervention hâtive ont des
impacts non-ciblés en étudiant la communauté de chenilles ainsi que son taux de
parasitisme. Une meilleure compréhension de son réseau trophique et des impacts
1des traitements permettra d’ajuster les protocoles afin d’en minimiser les effets non
ciblés.
2ÉTAT DES CONNAISSANCES
Écosystèmes forestiers et perturbations
Les écosystèmes forestiers qui représentent 40 % du territoire canadien sont à la
base de nombreux services écosystémiques, environnementaux, sociaux et
économiques (Ressource naturelles Canada 2021). Les provinces de l’est du
Canada telles que le Québec, les maritimes (Nouveau-Brunswick, Nouvelle-Écosse
et Île du Prince-Edward) et Terre-Neuve et Labrador sont des provinces dominées
par des forêts boréales composées de sapins et d’épinettes. Les forêts de feuillus
qui représentent une faible proportion du territoire forestier se retrouvent à basses
latitudes, particulièrement au sud du Québec et à l’ouest du Nouveau-Brunswick
(Beaudoin et al. 2014). Celles-ci sont dominées par les bouleaux et les érables. Plus
précisément, au Québec, l’écosystème forestier est séparé en trois zones de
végétation : la zone arctique, la zone boréale et la zone tempérée nordique
(Morneau 2021). Chacune des zones est subdivisée en domaines bioclimatiques
créant un gradient latitudinal passant du domaine de l’érablière à caryer cordiforme
au domaine de la toundra arctique herbacée. Au Nouveau-Brunswick, l’écosystème
forestier est séparé en sept écorégions passant d’une forêt de feuillus diversifiée à
l’ouest à une forêt de type boréale à l’est (SGS Belgium 2018).
Les perturbations forestières sont définies comme des évènements ponctuels qui
jouent un rôle dans la dynamique des écosystèmes en changeant les attributs
écologiques, la composition, la structure et l’organisation spatiale des zones
touchées (Pickett et White 1985, MacLean et al. 2001). Au Canada, environ 16 %
du territoire forestier a subi une perturbation dans la dernière décennie (Ressource
naturelles Canada 2021). Il existe trois types de perturbations naturelles sévères
dans les forêts nord-américaines : les feux, les épidémies d’insecte et les chablis.
Les feux et les épidémies d’insectes sont les perturbations principales des
peuplements dominés par les conifères alors que les peuplements de feuillus sont
plutôt affectés par les chablis.
3Au Canada, la superficie affectée par les feux de forêt dépend des conditions
climatiques et fluctue entre les années variant de 0,63 million d’hectares (en 2000)
à 7,38 millions d’hectares (en 1995), avec une moyenne de 2,48 millions d’hectares
affectés entre 1990 et 2019 (National Forestry Database 2022a, Fig. 1). À l’opposé,
la superficie affectée par la récolte varie peu entre les années, avec un minimum de
0,64 million d’hectares (en 2009), un maximum de 1,11 million d’hectares (en 2005)
et une moyenne de 0,91 million d’hectares, toutes méthodes de récolte confondues,
entre 1990 et 2019 (National Forestry Database 2022b). La perturbation affectant la
plus grande superficie forestière est celle causée par les insectes forestiers
(défoliation modérée à grave), soit entre 4,33 millions d’hectares (en 1997) et
42,05 millions d’hectares (en 1992) et en moyenne 16 millions d’hectares. Les
principaux insectes sont le dendroctone du pin ponderosa (Dendroctonus
ponderosae Hopkins, Coleoptera : Curculonidae), ravageant l’Ouest canadien, la
livrée des forêts (Malacosoma disstria Hübner, Lepidoptera : Lasiocampidae) et la
tordeuse des bourgeons de l’épinette (Choristoneura fumiferana Clemens,
Lepidoptera : Tortricidae, TBE), tous deux présents sur la majorité du territoire
canadien (Fig. 2). La TBE est responsable d’une défoliation modérée à grave de
1,89 million d’hectares en moyenne annuellement entre 1990 et 2019 avec un
maximum de 9,73 millions d’hectares défoliés en 1992 (National Forestry Database
2022c).
4Figure 1 : Superficie perturbée annuellement d’intensité modérée ou grave au
Canada entre 1990 et 2019 (National Forestry Database 2022a, 2022b, 2022c)
Figure 2 : Superficie perturbée d’intensité modérée ou grave par les principaux
insectes forestiers au Canada entre 1990 et 2019 (National Forestry Database
2022c).
5La tordeuse des bourgeons de l’épinette
En termes de superficie perturbée, la TBE (C. fumiferana) est le défoliateur le plus
important des forêts boréales d'Amérique du Nord (MacLean 1980; Fig. 3). Cette
espèce univoltine passe l’hiver à son deuxième stade larvaire, soit dans l’écorce,
dans les écailles de bourgeons ou dans les fleurs de l’arbre hôte dans un abri appelé
hibernaculum. Dès l’arrivée du printemps, soit quelques jours avant le débourrement
des aiguilles, aux mois d’avril et de mai, les chenilles émergent de leur diapause.
Elles se nourrissent des bourgeons et des aiguilles de l’année, en ordre de
préférence, du sapin baumier (Abies balsamea (L.) Mill.), de l’épinette blanche
(Picea glauca (Moench) Voss), de l’épinette rouge (Picea rubens Sarg.) et de
l’épinette noire (Picea mariana (Mill.)) (Hennigar et al. 2008). Le sapin baumier est
son hôte préféré dû au synchronisme entre son débourrement et l’émergence de la
TBE, suivi environ trois jours plus tard par les épinettes (Hennigar et al. 2008). Les
chenilles vont défolier les aiguilles et peuvent même attaquer les fleurs et cônes
mâles. Elles se nourrissent pendant environ huit semaines après la sortie de
diapause tout en se faisant un abri avec des fils de soie. Elles cessent de s’alimenter
après le sixième stade larvaire et se métamorphosent en chrysalide, pour une durée
d’environ 10 jours, puis le papillon adulte émerge. L’adulte peut se disperser sur plus
de 100 km grâce aux vents (Anderson et Sturtevant 2011, Garcia et al. 2022). Après
l’accouplement, les papillons femelles pondent leurs masses d’œufs en groupes de
15 à 20 sur les aiguilles des arbres hôtes en juillet et début aout, particulièrement
sur les aiguilles au soleil, au bout des branches (Régnière et Nealis 2007). Les œufs
éclosent entre sept et 14 jours plus tard et les jeunes chenilles passeront l’hiver au
deuxième stade larvaire. La limite de distribution septentrionale de la TBE est définie
par l’absence de ses plantes hôtes alors que sa limite méridionale est limitée par le
climat, soit des étés trop chauds (Candau et Fleming 2005).
6Figure 3 : Tordeuse des bourgeons de l’épinette (Choristoneura fumiferana
Clemens) : papillon adulte en haut et chenille au sixième stade larvaire en bas.
Source : Ressources naturelles Canada.
Depuis le 16e siècle, 11 épidémies de TBE ont eu lieu, les épidémies se produisant
aux 30-40 ans (Boulanger et Arseneault 2004) et durant en moyenne 10,2 +/- 3,8
ans (Boulanger et al. 2012). Une épidémie de TBE cause des retards de croissance
et, après 4-5 ans de défoliation, peut causer la mort de l’arbre (MacLean 1980). Au
pic de l’épidémie, la TBE peut causer jusqu’à 85 % de mortalité, et modifie ainsi la
composition en espèces, la structure d’âge et la dynamique des zones affectées soit
par la réinitialisation des peuplements ou par perturbation partielle (MacLean et al.
2001, Bouchard et al. 2006). Ses impacts sont plus importants dans les peuplements
de conifères que dans les peuplements mixtes (Bergeron et al. 1995, Bouchard et
al. 2005).
Pour la majorité des peuplements qui sont réinitialisés par l’épidémie la composition
de la régénération est mixte ou résineuse, seulement une faible proportion des
peuplements réinitialisés se régénérant en feuillus. Dans les territoires de zones de
perturbation partielle, il y a une ouverture du couvert qui mène à une diversité de
peuplements et change leur structure d’âge : les ouvertures causées par la TBE
permettent la croissance de la régénération menant à une forêt avec une structure
d’âge inéquienne (Bouchard et al. 2006, Chabot et al. 2014). Ces trouées créées
7par les épidémies donnent des caractéristiques de vieux peuplements soit la
formation de structure irrégulière et importante présence de bois mort (Bouchard et
al. 2006, Chabot et al. 2014). Au niveau du paysage, la combinaison des territoires
réinitialisés et partiellement réinitialisés crée une mosaïque hétérogène de
peuplement d’âge, de structure d’âge, de superficie et de composition différents.
Ces caractéristiques sont propres au territoire touché par des épidémies de TBE
(Leblanc et Bélanger 2000, Chabot et al. 2014).
Une épidémie d’insecte a plusieurs impacts sur les services écosystémiques de la
forêt (Schowalter 2012). Dans le cas d’une épidémie de TBE, les services
d’approvisionnement en bois sont affectés et des conséquences économiques sur
le secteur forestier surviennent par le ralentissement de la croissance et la mortalité.
Chang et al. (2012), à l’aide d’un modèle équilibré généralisé, estiment qu’une
épidémie sévère ou modérée entre 2012 et 2041 au Nouveau-Brunswick, sans
interventions, pourrait avoir des impacts économiques de 3,3 à 4,7 milliards de
dollars canadiens. Les services culturels et récréatifs sont aussi affectés : la qualité
visuelle des paysages affectés ou des lieux récréatifs moins sécuritaires pourrait
mener à une diminution de la fréquentation de ses lieux par la population
(Schowalter 2012).
Le réseau trophique de la TBE
Le réseau trophique de la TBE contient de nombreuses espèces de différents
niveaux trophiques : les plantes hôtes, des insectes herbivores, ainsi que de
nombreux ennemis naturels comme des entomopathogènes, des parasitoïdes et
des prédateurs (Eveleigh et al. 2007b).
Parasitoïdes
Un parasitoïde est un organisme qui pond ses œufs dans un hôte (endoparasitoïde)
ou sur un hôte (ectoparasitoïde), et qui cause inévitablement la mort de l’hôte lors
de son développement (Waage et Hassell 1982). Alors que les parasitoïdes
primaires attaquent des herbivores, les parasitoïdes secondaires attaquent plutôt les
8parasitoïdes primaires (Muller et al. 1999). Suivant la même logique, les parasitoïdes
tertiaires pondent leurs œufs dans des parasitoïdes secondaires. Les parasitoïdes
qui attaquent d’autres parasitoïdes sont appelés hyperparasitoïdes. Les
parasitoïdes sont donc un facteur de mortalité chez les insectes, offrant un service
écosystémique de lutte biologique (Costanza et al. 1997). Certains parasitoïdes sont
multivoltins, c’est-à-dire qu’ils complètent plusieurs générations en une saison. Ces
parasitoïdes vont avoir plusieurs espèces d’hôtes, des hôtes alternatifs, afin que
toutes les générations aient des hôtes disponibles, et ce tout au long de la saison.
Dans le réseau trophique de la TBE, plus d’une centaine de parasitoïdes sont
connus (Fernandez-Triana et Huber 2010), mais moins d’une dizaine sont fréquents
(Bouchard et al. 2018). Ils exercent un contrôle naturel de la population de TBE à
différents moments de son cycle épidémique. Lorsqu’une population épidémique est
en déclin, les parasitoïdes y contribuent (Régnière et Nealis 2007). Ils jouent aussi
un rôle lorsque les densités de population sont faibles : le taux de mortalité causé
par les parasitoïdes suffit pour maintenir les populations à de faibles densités
(Bouchard et al. 2018, Régnière et al. 2019). Cependant, à ma connaissance, le
réseau trophique entre deux épidémies n’a jamais été décrit dans son ensemble,
probablement due à la faible densité de population de TBE qui rend l’étude de son
réseau complexe.
Les réseaux trophiques décrits au pic et au déclin d’une épidémie indiquent que
ceux-ci sont dynamiques : la composition de la guilde de parasitoïdes change en
fonction du stade de l’épidémie, avec une plus grande diversité d’espèces de
parasitoïdes au pic de l’épidémie qu’au déclin. Ainsi, avec une augmentation de la
densité de TBE, il y a une augmentation du parasitisme par les parasitoïdes
spécialistes et les parasitoïdes secondaires et tertiaires, mais une diminution du
parasitisme par les parasitoïdes généralistes (Eveleigh et al. 2007 b). De plus,
comme les hôtes alternatifs limitent l’abondance des parasitoïdes, les parasitoïdes
univoltins sont généralement plus abondants durant les épidémies (McGugan et
Blais 1959, Maltais et al. 1989, Thireau et Régnière 1995). Par exemple, Apanteles
fumiferanae Viereck (Hymenoptera : Braconidae) et Glypta fumiferanae Viereck
9(Hymenoptera : Ichneumonidae) sont deux parasitoïdes univoltins attaquant la TBE
en grand nombre au moment où les densités de TBE augmentent et au pic de
l’épidémie (McGugan et Blais 1959, Miller 1963, Miller et Renault 1976). Apanteles
fumiferanae est un endoparasitoïde commun qui attaque les chenilles au premier ou
deuxième stades larvaires et cause leur mort au cinquième stade larvaire (Miller et
Renault 1976). Son taux de parasitisme augmente avec la densité de population de
TBE (Legault et James 2018) malgré un léger décalage (Miller et Renault 1976). À
la suite de plusieurs années d’épidémie, le parasitisme par A. fumiferanae diminue
dû à l’augmentation de la compétition par les autres espèces de parasitoïdes qui
sont présents à de fortes densités de population de TBE (Legault et James 2018).
Apanteles fumiferanae continue toutefois d’être un parasitoïde important au déclin
des populations (Miller 1963). Glypta fumiferanae est aussi un endoparasitoide
commun spécialiste de la TBE qui attaque les chenilles au deuxième stade larvaire
avant la diapause et qui les tue au sixième stade larvaire (Blais 1960, Dodge 1961,
Legault et James 2018). Le pic de population de G. fumiferanae est synchronisé
avec celui de la TBE (Miller 1963, Miller et Renault 1976). Ainsi, au pic de population
de TBE, G. fumiferanae cause entre 10 à 15 % de parasitisme puis le taux de
parasitisme diminue drastiquement lorsque l’épidémie décline (Miller et Renault
1976, Legault et James 2018).
À l’effondrement de l’épidémie, en plus de A. fumiferanae, G. fumiferanae, Meteorus
trachynotus Viereck (Hymenoptera : Braconidae) et Actia interrupta Curran
(Diptera : Tachinidae) sont abondants (Jaynes et Drooz 1952, McGugan et Blais
1959, Blais 1960). Meteorus trachynotus est un parasitoïde koinobionte, c’est-à-dire
que les chenilles parasitées continuent de s’alimenter et de se développer après
avoir été parasitées. Ce parasitoïde attaque les chenilles de TBE de stades cinq et
six et, à cause de sa dépendance à la densité (Régnière et al. 2021a), est le
parasitoïde le plus important lors des déclins des populations de TBE (Dowden et
al. 1950, Jaynes et Drooz 1952, McGugan et Blais 1959, Blais 1960, Régnière et
Nealis 2007), causant jusqu’à 40 % de parasitisme (Miller et Renault 1976).
Meteorus trachynotus est multivoltin, pouvant produire de une à quatre générations
complètes par été (Régnière et al. 2021a), et passe possiblement l’hiver dans la
10tordeuse à bandes obliques (Choristoneura rosaceana Harr, Lepidoptera :
Tortricidae) (Maltais et al. 1989). Actia interrupta attaque la TBE lorsqu’elle est à ses
derniers stades larvaires et est présent après la chute de densité des populations
de TBE (Cusson et al. 2002). Actia interrupta peut produire entre deux et quatre
générations par saison selon son emplacement géographique, se synchronisant
avec l’hôte dans lequel il passe l’hiver (Régnière et al. 2021 b).
À faible densité de population de TBE, les parasitoïdes généralistes comme
Elachertus cacoeciae Howard (Hymenoptera : Eulophidae), Tranosema rostale
Brischke (Hymenoptera : Ichneumonidae) et Phryxe pecosensis Townsend
(Diptera : Tachinidae) prennent de l’importance, empêchant les populations de TBE
d’augmenter (McGugan et Blais 1959, Blais 1960, Miller et Renault 1976, Miller
1990, Cusson et al. 1998, Fidgen et Eveleigh 1998, Eveleigh et al. 2007b, Bouchard
et al. 2018a). Elachertus cacoeciae est un ectoparasitoide attaquant la TBE à partir
de son quatrième stade larvaire. C’est un des parasitoïdes d’importance, pouvant
être responsable de jusqu’à 32 % du parasitisme à basse densité de population
(Miller 1963). Tranosema rostrale, un des parasitoïdes présents entre deux
épidémies (Miller et Renault 1976, Cusson et al. 1998, Eveleigh et al. 2007 b,
Bouchard et al. 2018a), attaque les chenilles de TBE aux stades larvaire trois à six
(Seehausen et al. 2016). Tranosema rostrale peut effectuer de une à quatre
générations par saison (Régnière et al. 2020) en se synchronisant avec ses hôtes,
différentes espèces de tortricidé (Seehausen 2017). Il pourrait passer l’hiver dans
un de ses hôtes alternatifs, la tordeuse à bandes obliques. Phryxe pecosensis est
un diptère généraliste, reconnu pour attaquer la TBE et d’autre espèce de
Choristoneura (O’Hara 2005). Il attaque les chenilles aux stades larvaires quatre et
cinq et émerge du sixième stade larvaire. Il a un taux de parasitisme variable chez
la TBE qui peut atteindre 15 % (Blais 1960, Dowden 1950).
Hôtes alternatifs
La majorité des parasitoïdes connus de la TBE étant multivoltins, la présence
d’hôtes alternatifs est nécessaire. Effectivement, la TBE est disponible comme hôte
11au début de la saison, mais les parasitoïdes des générations suivantes devront
trouver d’autres espèces hôtes, afin de compléter leur cycle de vie. Par exemple,
l’hôte d’hivernation du parasitoïde de la TBE, Meteorus trachynotus, est la tordeuse
à bandes obliques, une espèce de lépidoptère sans laquelle M. trachynotus ne peut
compléter son cycle (Maltais et al. 1989). Les hôtes alternatifs sont d’autant plus
importants pour le contrôle des populations exercé par les parasitoïdes de la TBE
dû au principe de compétition apparente (Holt et Lawton 1993). Effectivement, dans
le cas d’un parasitoïde multivoltin, l’augmentation d’un hôte va augmenter la
population de parasitoïde et peut ainsi diminuer la population de l’autre hôte. Un
exemple de compétition apparente a été observé par Pfannenstiel et al. (2010) dans
des vergers et par Settle et Wilson (1990) : le parasitisme par Anagrus spp.
(Hymenoptera : Mymaridae) est augmenté au point de réduire les populations de
cicadelles dans les vignes lorsque son hôte d’hivernation est présent.
De manière générale, une diversité de plantes dans l’écosystème forestier diminue
l’herbivorie causée par des insectes (Jactel et Brockerhoff 2007), et c’est également
le cas pour la TBE : la présence de feuillus diminue la sévérité des épidémies
(MacKinnon et MacLean 2003, Colford-Gilks et al. 2012, Zhang et al. 2018, 2020a).
Cet effet peut s’expliquer par l’hypothèse de la fragmentation de l’habitat qui stipule
que les individus auraient plus de difficulté avec la recherche de nourriture, la
dispersion et l’accouplement dans un habitat diversifié (Zhang et al. 2020b). Une
autre explication est l’hypothèse des ennemis naturels qui stipule qu’une plus grande
diversité de plantes favorise la présence de parasitoïdes (Cappuccino et al. 1998,
Quayle et al. 2003, Eveleigh et al. 2007b, Legault et James 2018, Greyson-Gaito et
al. 2022). Effectivement, une plus grande diversité de plantes offre une diversité de
niches plus importante et peut donc accueillir plus d’espèces d’hôtes alternatifs
(Cappuccino et al. 1998, Siemann et al. 1998, Eveleigh et al. 2007 b). Aussi, la
présence plus importante de fleurs dans les peuplements diversifiés augmente la
ressource nutritive pour les parasitoïdes adultes, par leur pollen et nectar, et va
influencer positivement et directement la présence et l’activité des parasitoïdes
(Simmons et al. 1975, Russell 1989, Bradette 1994, Cappuccino et al. 1999).
Dépendamment des traits d’histoire de vie des espèces, notamment de leur besoin
12pour des hôtes alternatifs ou non, la présence de feuillus augmente ou diminue leur
parasitisme (Cappuccino et al. 1998, Legault et James 2018). Par exemple, A.
fumiferanae et les Tachinidae (tels que A. interrupta et P. pecosensis) sont favorisés
par des peuplements mixtes alors que ces derniers ont un effet négatif sur G.
fumiferanae (Simmons et al. 1975, Legault et James 2018).
Autres ennemis naturels
La TBE compose une partie importante de l’alimentation des oiseaux forestiers,
principalement la paruline obscure (Leiothlypis peregrina Wilson), la paruline tigrée
(Dendroica tigrina Gmelin) et la paruline à poitrine baie (Setophaga castanea
Wilson) (McMartin et al. 2002). Une épidémie a donc un effet positif pour les oiseaux
(Venier et Holmes 2010). Leur rôle dans la régulation des populations de TBE varie
en fonction du stade de l’épidémie. Lorsque les densités de population de TBE sont
élevées, le taux de prédation est faible, soit de 7 % (Venier et Holmes 2010).
Toutefois, lorsque les populations de TBE sont en déclin, la prédation par les
oiseaux est une source de mortalité significative à partir du quatrième stade larvaire
et jusqu’aux papillons (Régnière et al. 2021 c). Puis, lorsque les densités de
population sont faibles, la consommation de chenilles de TBE contribue à maintenir
les populations de TBE à de faibles densités (Morris 1963, Venier et Holmes 2010).
En plus de la prédation par les oiseaux, de la prédation par des amphibiens, des
petits mammifères et des arthropodes tels que des coléoptères, des guêpes, des
fourmis et des araignées a aussi été observée (Blais 1960, Jennings et Houseweart
1984, Jennings et al. 1999, Bouchard et al. 2018a).
Plusieurs virus, champignons, bactéries et microsporidies infectent la TBE, le plus
commun étant la microsporidie Nosema fumiferanae (Thomson) (Wilson 1977).
Nosema fumiferanae cause des retards de développement chez les chenilles et les
chrysalides, des retards d’émergence et plus de maladies chez les papillons, et se
transmet de la femelle à sa progéniture jusqu’à atteindre un niveau d’infection mortel
(van Frankenhuyzen et al. 2007, Eveleigh et al. 2007a). Le taux d’infection
13augmente avec le stade de l’épidémie pour se rendre jusqu’à 15 % de mortalité
annuellement (Eveleigh et al. 2012).
Dynamique de population
Pureswaran et al. (2016) colligent les trois théories développées sur la dynamique
de population de la TBE : l’hypothèse de la sylviculture, de l’oscillation et du multi-
équilibre. L’hypothèse de la sylviculture se base sur la structure de la forêt,
influencée par nos pratiques d’aménagement forestier, pour expliquer les
épidémies. Cette théorie, supportée par Kettela (1983), suppose que les populations
de TBE restent à de faibles densités tant que la ressource alimentaire n’est pas
optimale : faible disponibilité et bonne vigueur des sapins baumiers. Lorsque les
sapins baumiers sont davantage disponibles, plus vieux, et donc moins vigoureux,
jumelé à une diminution de l’impact des ennemis naturels de la TBE, les populations
de TBE augmentent en densité jusqu’à ce que les ressources deviennent limitées.
Cette hypothèse a été critiquée par plusieurs puis reprise par Cappuccino et al.
(1998) qui la remanient et proposent qu’une composition forestière diversifiée
permette une plus grande diversité de parasitoïdes, expliquant le plus faible impact
des épidémies de TBE dans les peuplements mixtes (MacKinnon et MacLean 2003,
Colford-Gilks et al. 2012, Robert et al. 2012). Cette théorie recommende un
changement dans les pratiques de sylviculture pour une augmentation de la
présence de feuillus (Pureswaran et al. 2016).
La théorie de l’oscillation, proposée par Royama (1984), suppose que les épidémies
sont cycliques et se base sur la théorie de la dépendance à la densité entre les
ennemis naturels et la TBE. À basse densité de population de TBE, le contrôle est
exercé par les ennemis naturels. Toutefois, lorsque les populations de TBE sont très
faibles, les populations d’ennemis naturels déclinent. La diminution d’abondance et
la diminution de l’impact des ennemis naturels permet aux populations de TBE de
devenir épidémiques. Les épidémies sont synchronisées au niveau régional, dû à
des climats régionaux favorables et à la dispersion des papillons. Les chenilles de
TBE étant abondantes, les populations de parasitoïdes augmentent, ce qui
14augmente le taux de mortalité chez les chenilles et, conjugué avec un déclin des
ressources, ramène les populations à un niveau non épidémique.
La théorie du multi-équilibre (ou double équilibre) décrite par Clark et al (1979)
suppose, comme son nom l’indique, qu’il y a deux équilibres dans la dynamique de
population de la TBE. Au premier équilibre, lorsque la population est de faible
densité, les TBE ont de la difficulté à trouver des partenaires, n’ont pas un bon
succès de dispersion (effet Allee) (Régnière et al. 2013) et le contrôle exercé par les
ennemis naturels est suffisant pour maintenir les populations à cet équilibre, comme
pour la théorie de l’oscillation (Bouchard et al. 2018b). Pour des raisons
stochastiques (bonne reproduction, climat favorable, ressources abondantes, etc.),
le taux de mortalité diminue ou le taux d’immigration augmente, ce qui mène à une
augmentation rapide de la population qui échappe au contrôle naturel. La population
atteint alors le deuxième équilibre, à haute densité de population, proche de la
capacité de soutien de la forêt. À ce moment, le taux de survie de la population de
TBE est élevé et stable sauf à l’occasion de quelques chutes desquelles elle se
remet. Ces diminutions dans la survie peuvent être causées par une maladie, un
manque de nourriture, la mortalité lors de l’émigration ou autre. Le vrai déclin de la
population se produit directement après l’une de ces chutes de laquelle la population
ne se remet pas. La densité de population de TBE décline jusqu’au point où les
ennemis naturels peuvent reprendre le contrôle (Régnière et Nealis 2007). Selon
cette théorie, les épidémies sont soudaines, se développent localement et sont
contagieuses via la migration des papillons, un facteur clé de la propagation de
l’épidémie (Larroque et al. 2020). L’atteinte du deuxième équilibre, épidémique,
étant facilité par l’immigration d’individus d’une épidémie à proximité (Régnière et al.
2019), un contrôle des populations en visant les populations locales pourrait limiter
les épidémies régionales (Pureswaran et al. 2016, Johns et al. 2019).
La théorie de l’oscillation est la théorie qui reçoit le plus de confirmation. Toutefois,
des études récentes continuent de justifier la théorie du multi-équilibre. Selon
Pureswaran et al. (2016), peu d’études ont été réalisées sur la transition entre des
populations de TBE de faibles densités à des populations en croissance. Des études
15à de basses densités permettraient de compléter les connaissances sur la
dynamique de population.
Lutte contre la TBE
La lutte contre la TBE peut être biologique ou chimique. Une option de lutte
biologique contre la TBE à l’aide de Trichogramma minutum Riley (Hymenoptera:
Trichogrammatidae), un parasitoïde indigène en Amérique du Nord qui attaque les
œufs, a été étudiée en Ontario et au Québec (Smith et al. 1990, Martel et al. 2021).
Les résultats des lâchers de T. minutum lors de l’épidémie de 1980 indiquent que la
méthode est efficace avec un taux de parasitisme de 83 %, réduisant les populations
de TBE et pouvant ainsi prévenir la défoliation des conifères (Smith et al. 1990). Les
études plus récentes ont démontré l’efficacité des drones pour l’épandage de ces
parasitoïdes (Martel et al. 2021).
Toutefois, la majorité des traitements effectués pour lutter contre la TBE se font à
l’aide d’insecticides. À l’échelle du Canada, l’insecte forestier à l’origine des plus
grandes aires traitées est la TBE, avec un total d’environ cinq millions d’hectares de
forêt traités entre 1990 et 2019, suivi par l’arpenteuse de la pruche, avec environ
650 000 hectares de forêt traités (National Forestry Database 2022d). Au Québec,
une épidémie est en cours depuis 2006, atteignant plus de 12 millions d’hectares,
dont quatre millions de défoliation modérée ou grave en 2021 (Ministère des Forêts
de la Faune et des Parcs 2021) touchant plusieurs régions administratives, dont la
région de la Gaspésie-Îles-de-la-madeleine. Cette épidémie a mené au traitement
de 186 000 hectares, à l’aide de Btk, en 2018, dans une gestion d’épidémie de type
protection du feuillage (National Forestry Database 2022d). Adjacent à cette
province, bien que le Nouveau-Brunswick fasse face à des migrations de papillons
de TBE en provenance du Québec depuis plusieurs années, les populations restent
sous contrôle et la défoliation est presque inexistante : en 2021, 85 000 hectares ont
été traités contre la TBE, à l’aide de Btk et de tébufénozide (D. Carleton,
communication personnelle).
16Vous pouvez aussi lire

















































