Étude du réseau de projections transzonales durant la folliculogenèse et de la méthylation dans la stabilisation des ARN messagers
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Étude du réseau de projections transzonales durant la
folliculogenèse et de la méthylation dans la stabilisation
des ARN messagers
Mémoire
Karine Dubuc
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Karine Dubuc, 2021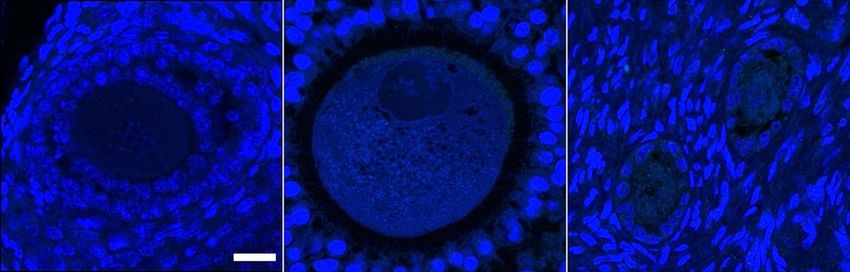
Étude du réseau de projections transzonales durant la
folliculogenèse et de la méthylation dans la
stabilisation des ARN messagers
Mémoire
Karine Dubuc
Sous la direction de :
Claude Robert, directeur de recherche
Géraldine Delbès, codirectrice de recherche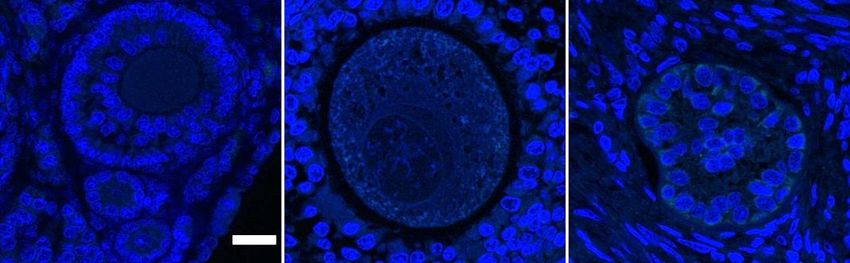
Résumé
La croissance ovocytaire comprend l’établissement de projections transzonales formant un
réseau de communication entre le gamète et les cellules somatiques entourant ce dernier.
Une accumulation des ARNm dans le cytoplasme se produit lors des premiers stades de
croissance de l’ovocyte afin de soutenir la maturation durant le silence transcriptionnel qui
perdure jusqu’à l’activation du génome embryonnaire. Selon l’espèce, cette période peut
durer des jours voir des semaines nécessitant une grande stabilité des ARNm maternels. Il
est connu que la communication entre l’ovocyte et les cellules somatiques ainsi que
l’utilisation des réserves d’ARNm sont essentielles pour l’acquisition des compétences de
l’ovocyte afin de soutenir la fécondation et le développement embryonnaire précoce. Des
modifications chimiques connues comme ayant un rôle stabilisateur, tel que la méthylation,
ont également été détectées sur les ARNm.
L’hypothèse est que des modifications chimiques de l’ARNm, soit l’ajout de groupements
méthyles, sont impliquées dans la stabilisation des transcrits au sein de l’ovocyte durant le
silence transcriptionnel. Le réseau de communication pourrait jouer un rôle en faisant le
transfert de protéines permettant la méthylation de l’ARNm. Les objectifs sont de détecter
les différentes modifications chimiques ayant lieu dans le transcriptome de l’ovocyte ainsi
que localiser et caractériser les protéines ayant pour rôle de catalyser, détecter et inverser
les modifications chimiques identifiées sur l’ARN.
Différents types de modifications post-transcriptionnelles ont été détectées au sein de
l’ovocyte et les résultats montrent que m6A et m5C sont les modifications les plus
abondantes. La caractérisation des protéines impliquées dans ces modifications dans
l’ovaire et l’ovocyte de la souris, du porc et du bovin montre une activité de méthylation
différente selon l’espèce durant la folliculogenèse et l’ovogenèse. De plus amples
recherches devront être réalisées afin de bonifier la compréhension des mécanismes
impliqués dans la production d’un ovocyte de qualité. Les travaux présentés contribuent à
l’avancement de ces connaissances.
ii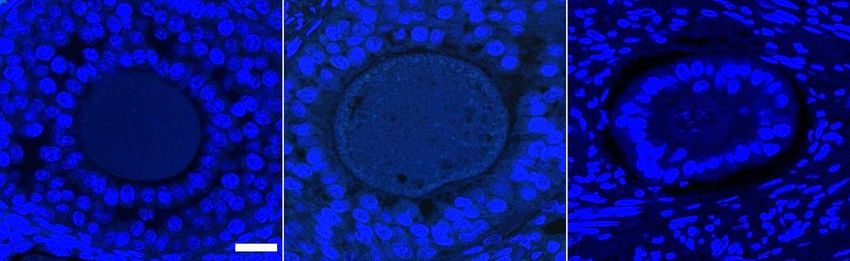
Abstract
Oogenesis, which occurs within the ovarian follicle, is a process closely link to
folliculogenesis. The oocyte growth mainly involves the establishment of a communication
network between the gamete and surrounding somatic cells. During the early stages of
oocyte growth, the gamete accumulates mRNAs in its cytoplasm to support maturation and
first cell division during transcriptional silencing until the embryonic genome activation.
This period can last from days to weeks depending on the species requiring long stability
from maternal mRNAs. It is known that communication between oocyte and somatic cells
and use of mRNA reserves are essential for oocyte competence acquisition to sustain
fertilization and early embryogenesis. Chemical modifications known to have stabilizing
role, such as methylation, have also been detected on the mRNAs.
The main hypothesis is the chemical modifications of mRNA, by the addition of methyl
groups, that are involved in stabilizing and managing transcripts within oocyte during
transcriptional silencing. The communication network could have a role in enabling
proteins transfer from cumulus cells allowing mRNA methylation. Objectives are to detect
various chemical modifications taking place in oocyte transcriptome and to locate and
characterize proteins having role of managing the chemical modifications identified on
RNA.
Many post-transcriptional modifications have been detected within the oocyte and results
have shown that m6A and m5C are modifications with higher abundant expression. Proteins
characterization involved in these post-transcriptional modifications in ovary and oocyte
of mouse, swine and bovine showed methylation activity during folliculogenesis and
oogenesis. More research is needed to improve understanding of mechanisms involved in
oocyte competences acquisition, but the present study has contributed to the advancement
of this knowledge.
iii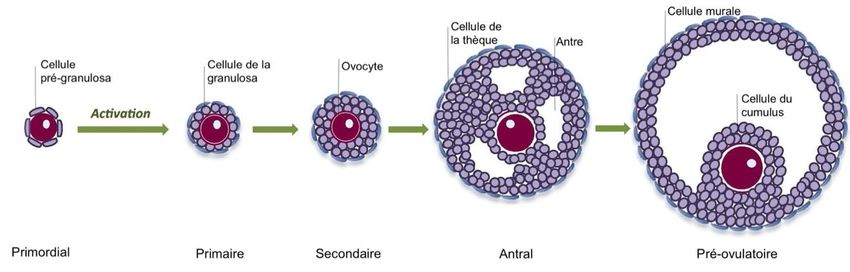
Table des matières
Résumé................................................................................................................................ ii
Abstract .............................................................................................................................. iii
Table des matières.............................................................................................................. iv
Liste des tableaux .............................................................................................................. vii
Liste des figures ............................................................................................................... viii
Liste des abréviations .......................................................................................................... x
Remerciements .................................................................................................................. xii
Avant-propos.................................................................................................................... xiii
Introduction ......................................................................................................................... 1
CHAPITRE 1 : Revue de littérature ................................................................................... 3
1. Folliculogenèse ........................................................................................................... 3
1.1 Folliculogenèse in vitro....................................................................................... 6
2. Ovogenèse ................................................................................................................... 8
2.2 Culture in vitro .................................................................................................. 11
3. Production d’ovocytes in vivo et cycle œstral bovin ................................................ 12
4. Facteurs affectant la folliculogenèse et l’ovogenèse ................................................ 13
4.1 Cellules du cumulus .......................................................................................... 14
3.1 Projections transzonales .................................................................................... 15
5. Principes du domaine neuronal appliqués à l’ovaire ................................................ 18
5.1 Gène Fragile X mental retardation 1 (FMR1)................................................... 18
5.2.1 FMRP ........................................................................................................ 20
6. Finalité de la folliculogenèse : produire un ovocyte de qualité ................................ 20
6.1 Maturation moléculaire et activité transcriptionnelle ....................................... 21
6.2 Maturation cytoplasmique ................................................................................ 22
6.2.1 Mitochondries ........................................................................................... 23
6.2.2 Lipides....................................................................................................... 24
6.2.3 Granules corticaux .................................................................................... 24
6.3 Maturation méiotique ........................................................................................ 25
6.3.1 Adénosine monophosphate cyclique (AMPc) et guanosine monophosphate
cyclique (GMPc) ....................................................................................................... 25
7. Qualité et compétence au développement de l’ovocyte ............................................ 27
8. Nouvelles considérations en lien avec l’épitranscriptome ........................................ 29
iv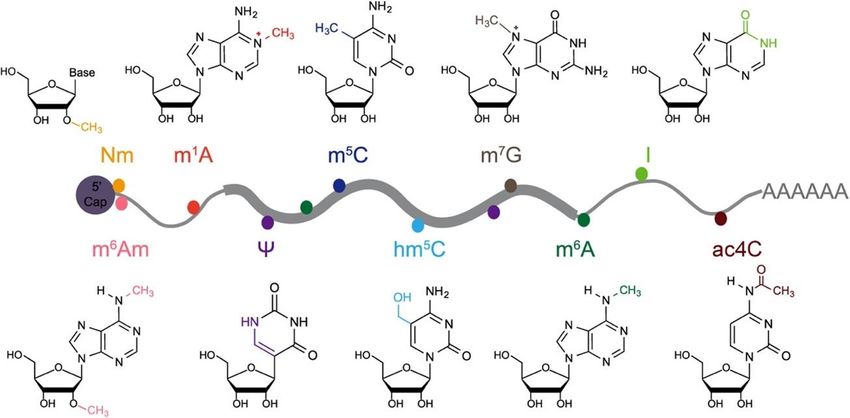
8.1 Acide ribonucléique (ARN) .............................................................................. 29
8.1.1 Stabilisation et dormance des ARNm maternels ...................................... 30
8.2 5-methylcytosine (m⁵C) .................................................................................... 32
8.3 N6-methyladenosine (m6A) ............................................................................... 32
8.4 Writers............................................................................................................... 34
8.4.1 Wilms’ tumor 1-associating protein (WTAP)........................................... 34
8.4.2 ADN Méthyltransférase 2 (DNMT2)........................................................ 35
8.5 Readers ............................................................................................................. 36
8.5.1 Methyl binding domain 2 (MBD2) ........................................................... 36
8.5.2 YTH-domain family member 2 (YTHDF2) ............................................... 37
8.6 Erasers .............................................................................................................. 38
8.6.1 Fat mass and obesity-associated protein (FTO) ....................................... 38
8.6.2 AlkB homolog 5 (ALKBH5) ..................................................................... 38
Conclusion ........................................................................................................................ 39
Liste des ouvrages cités .................................................................................................... 40
CHAPITRE 2: Distribution of epi-transcriptome-modifying proteins in mammalian
oocytes .............................................................................................................................. 48
Résumé.............................................................................................................................. 48
Abstract ............................................................................................................................. 49
2.1 Introduction ....................................................................................................... 50
2.2 Materials and Methods ...................................................................................... 52
2.2.1 Animals ..................................................................................................... 52
2.2.2 RNA isolation ........................................................................................... 52
2.2.3 RNA enzymatic digestion and LC-MS/MS analysis ................................ 52
2.2.4 Ovary section immunohistofluorescence .................................................. 53
2.2.5 Whole-mount oocyte immunofluorescence .............................................. 54
2.2.6 Ovary and oocyte imaging ........................................................................ 54
2.3 Results ............................................................................................................... 55
2.3.1 Identification and measurement of RNA modifications by LC-MS/MS
analysis 55
2.3.2 Detection of RNA-modifying proteins ..................................................... 55
2.3.3 Detection of writer proteins during folliculogenesis ................................ 55
2.3.4 Detection of reader proteins ...................................................................... 56
2.3.5 Detection of RNA methylation eraser proteins ......................................... 57
2.4 Discussion ......................................................................................................... 57
v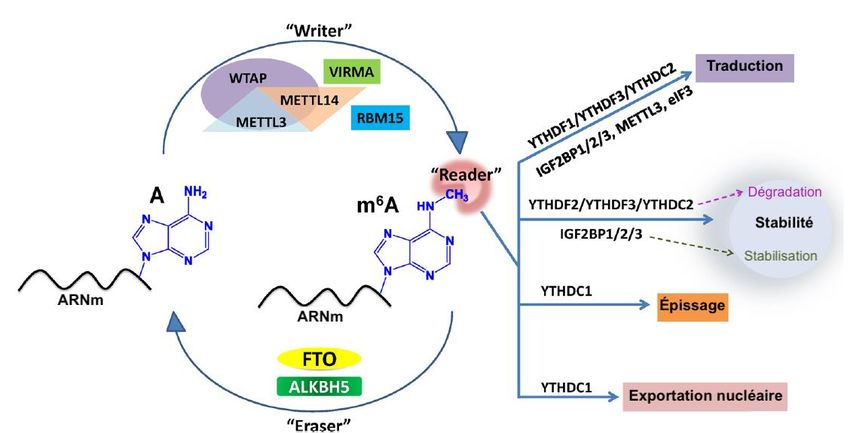
2.5 Conclusion ........................................................................................................ 62
2.6 Figures and legends........................................................................................... 63
2.7 Supplementary Data .......................................................................................... 72
Conclusion générale .......................................................................................................... 77
References ......................................................................................................................... 79
vi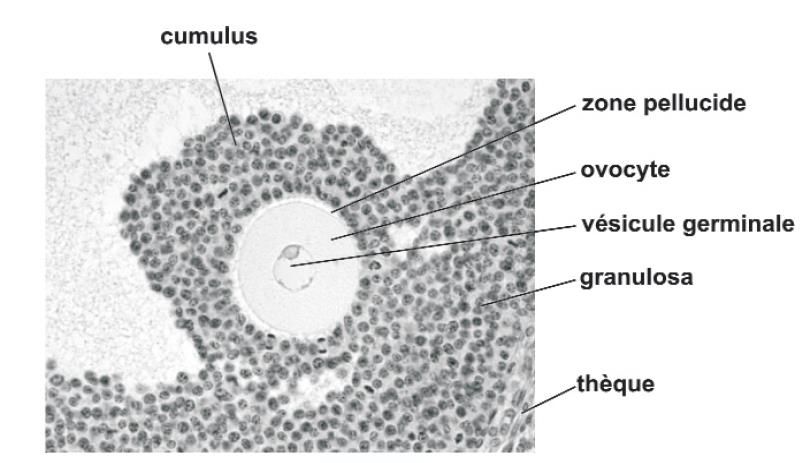
Liste des tableaux
Table 2.1 Localisation of RNA-modifying enzyme protein in early antral follicles and
mature oocytes .................................................................................................. 70
Table 2.2 Stage and diameter of the follicles used in this study and the corresponding
chromatin stage and transcription activity reported in the literature ................ 71
Table 2.3 Description of the antibodies used for immunofluorescence in this study ....... 72
vii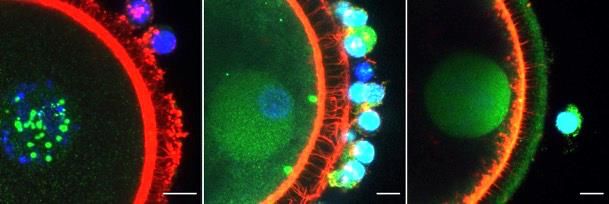
Liste des figures
Figure 1.1 Développement folliculaire chez les mammifères. Adapté de Sánchez and Smitz
(2012). .............................................................................................................. 3
Figure 1.2 Stades et la durée (jours) de la folliculogenèse au sein de l’ovaire adulte chez
l'humain. Adapté de Gougeon (1986) .............................................................. 5
Figure 1.3 Maturation d’un ovocyte de mammifère. .......................................................... 9
Figure 1.4 Relation entre la taille des follicules et ovocytes durant leur développement chez
la vache. Adapté de Fair (2003). .................................................................... 10
Figure 1.5 Coupe histologique d'un follicule ovarien au stade antral chez la souris. Adapté
de Monniaux et al. (2009). ............................................................................. 14
Figure 1.6 Ovocyte bovin et projections transzonales marquées à la rhodamine phalloidine.
Adapté de Macaulay et al (2014). .................................................................. 15
Figure 1.7 Mutation et prémutation du gène FMR1. Adapté de Hagerman et al. (2009)..
........................................................................................................................ 19
Figure 1.8 Modifications chimiques principales se retrouvant sur des ARNm. Adapté de
Song and Yi (2020). ....................................................................................... 30
Figure 1.9 Méthylation de l'adénosine en position N6 sur un ARNm. Adapté de Deng et al.
(2018). ............................................................................................................ 34
Figure 2.1 Relative expression of modifications of porcine somatic and germinal cell RNA,
based on mass spectrometry (A) N1-methyladenosine, (B) N6-
methyladenosine, (C) 5-methylcytidine, (D) 7-methylguanosine .................. 63
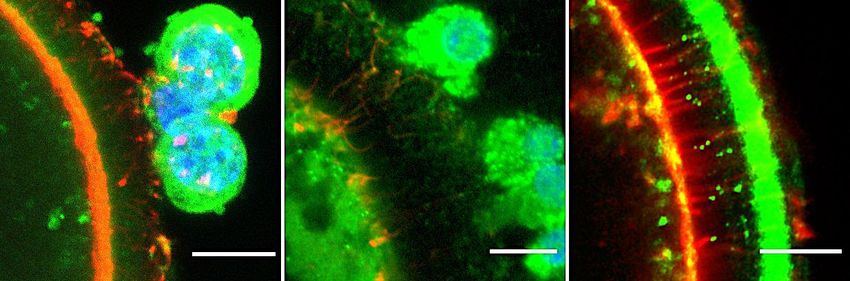
Figure 2.2 Representative z-stack confocal images of TRDMT1 and WTAP writer protein
distribution in murine, porcine and bovine germinal tissue obtained using
fluorescent antibodies (green). A – Sections of ovary showing secondary (S),
early antral (Ea) and antral (An) follicle stages (bar = 20 m). B – Whole
oocyte mounts (bar = 10 m). Blue is due to Hoechst DNA stain and red is due
to Acti-Stain 555 actin stain. .......................................................................... 65
Figure 2.3 Representative z-stack confocal images of MBD2, YTHDF2 and FMRP reader
protein distribution in murine, porcine and bovine germinal tissue obtained
using fluorescent antibodies (green). A – Sections of ovary showing primary
(Pr), secondary (S) and early antral (Ea) follicle stages (bar = 20 m). B –
Whole oocyte mounts (bar = 10 m). Blue is due to Hoechst DNA stain and
red is due to Acti-Stain 555 actin stain. ......................................................... 67
Figure 2.4 Representative z-stack confocal images of ALKBH5 and FTO eraser protein
distribution in murine, porcine and bovine germinal tissue obtained using
fluorescent antibodies (green). A – Sections of ovary showing primary (Pr),
secondary (S) and early antral (Ea) and antral (An) follicle stages (bar = 20
m). B – Whole oocyte mounts (bar = 10 m). Blue is due to Hoechst DNA
stain and red is due to Acti-Stain 555 actin stain. .......................................... 69
Figure 2.5 Ovarian section in paraffin, negative control without primary antibody, showing
no specific staining by the secondary antibody. DNA is stained with Hoescht
33342 (blue). Bar = 20 µm. ............................................................................ 74
viii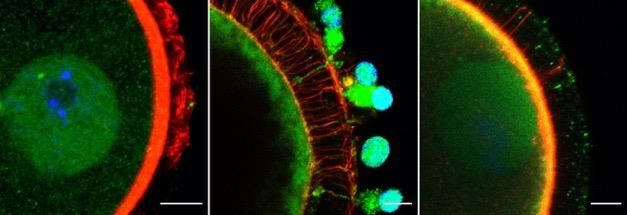
Figure 2.6 Oocyte whole mount, negative control without primary antibody, showing no
specific staining by the secondary antibody. DNA is stained with Hoescht
33342 (blue) and actin with Acti-stain 555 (red). Bar = 20 µm (same
magnification for pig and cow). ..................................................................... 76
ixListe des abréviations
3’UTR 3’ Untranslated Transcribed Region
5’UTR 5’ Untranslated Transcribed Region
A Adénine
Ac4C N4-acetylcytidine
ADN Acide désoxyribonucléique
ALKBH5 AlkB homologue 5
AMPc Adénosine monophosphate cyclique
ARN Acide ribonucléique
ARNm Acide ribonucléique messager
ARNr Acide ribonucléique ribosomal
ARNt Acide ribonucléique de transfert
ATP Adénosine-TriphosPhate
BMP15 Bone morphogenetic protein-15
BMP7 Bone morphogenetic protein-7
BTG4 BTG Anti-Proliferation Factor 4
C Cytosine
COC Complexe ovocyte cumulus
DNA Deoxyribonucleic acid
DNMT2 DNA methyltransferase 2
EGF Epidermal growth factor
FAO Fatty beta-oxidation
FMR1 Fragile X mental retardation 1
FMRP Fragile X mental retardation protein
FSH Hormone folliculostimulante
FTO Fat mass and obesity-associated protein
FXPOI fragile X premature ovarian insufficiency
FXS Syndrome du X fragile
G Guanine
GFRA1 Family receptor alpha-1
GMPc Guanosine monophosphate cyclique
GV Germinal vesicle
GVBD Germimal vesicle breakdown
hm5C 5-Hydroxymethylcytidine
I Inosine
LC-MS/MS Liquid Chromatography with tandem mass spectrometry
LH Hormone lutéinisante
lncRNAs Long non-coding RNA
m1 A N1-methyladenosine
m⁵C 5-methylcytosine
m6A N6-methyladenosine
m6 Am 2’-O-dimethyladenosine en position N6
m7 G 7-methylguanosine
xMBD Methyl-CpG binding domain
MBD2 Methyl binding domain 2
MBD3 Methyl binding domain 3
METTL3 Methyl-transferase like 3
METTL14 Methyl-transferase like 14
mRNA Messenger Ribonucleic acid
NES Nuclear Export Signal
NLS Nuclear Localization Sequence
Nm 2’-O-Methylation
PBS Paraformaldehyde in phosphate-buffered saline
PBS-PVA Paraformaldehyde in PBS-polyvinyl alcohol
PCOS Polycystic ovary syndrome
PDE Phosphodiestérase
PDE1 Phosphodiestérase 1
PDE3 Phosphodiestérase 3
PDE4 Phosphodiestérase 4
PDE5 Phosphodiestérase 5
PDE8 Phosphodiestérase 8
PDE11 Phosphodiestérase 11
PDE3A Phosphodiestérase 3A
PDE4D Phosphodiestérase 4D
PDE8A Phosphodiestérase 8A
PDE8B Phosphodiestérase 8B
PTZs Projections transzonales
RMPs RNA-modifying proteins
RNA Ribonucleic acid
RNP Ribonucléoprotéine
SAM S-adenosyl methionine
TBST Tris-buffered saline with Tween 20
U Uracile
UTR Untranslated Transcribed Region
TRDMT1 TRNA Aspartic Acid Methyltransferase 1
WEE2 Wee1-like protein kinase 2
WTAP Wilms’ tumor 1-associating protein
WPAP1 Wilms tumor-associated protein 1
XIST X Inactive Specific Transcrit
YBX2 Y-box binding protein 2
YTH YT521-B homology
YTHDF1 YTH-domain family member 1
YTHDF2 YTH-domain family member 2
Ψ Pseudouridine
xiRemerciements
J’aimerais tout d’abord remercier mon directeur de recherche, Claude Robert, pour sa
confiance en mes capacités, ses conseils judicieux et son encadrement. Il a su contribuer à
mon autonomie ainsi qu’au développement de mon esprit critique.
Je tiens à remercier spécialement Isabelle Gilbert et Alexandre Bastien qui m’ont, non
seulement aidée, mais également soutenue et encouragée tout au long de mon parcours. Je
suis reconnaissante envers leur écoute et leur grande patience.
Je voudrais remercier l’aide précieuse d’Isabelle Laflamme et de Mathilde Marchais qui
ont grandement contribué à la réalisation de mes travaux.
À ma famille et spécialement mon conjoint, merci de m’avoir appuyée tout au long de mes
études graduées. Votre présence a sans aucun doute contribué à la réussite de cette maîtrise.
Je vous remercie pour votre écoute durant les moments plus difficiles.
J’aimerais également remercier tous les étudiants des laboratoires de Claude Robert et de
Marc-André Sirard qui sont devenus plus que des collègues de travail, mais bien des amis
qui ont su m’apporter du soutien et de l’entraide lorsque cela devenait plus difficile. Grâce
à vous, je garde d’excellents souvenirs de mes études graduées et notre esprit d’équipe me
manquera certainement.
Finalement, je tiens à remercier ma co-directrice ainsi que tous les partenaires ayant rendu
mon projet de maîtrise possible. Merci au Fonds Québécois de la Recherche sur la Nature
et les Technologies, au Conseil de recherches en sciences naturelles et en génie du Canada,
à Transbiotech, au Réseau Québécois en Reproduction ainsi qu’au Centre de recherche en
reproduction, développement et santé intergénérationnelle.
xiiAvant-propos
Ce mémoire comporte un chapitre rédigé sous forme d’article scientifique en anglais
portant le titre «Distribution of epi-transcriptome-modifying proteins in mammalian
oocytes». En raison de problèmes d’analyse de résultats bioinformatiques, l’article n’a pu
être complété pour être soumis à un journal actuellement. Je suis l’auteure principale de
cet article et les co-auteurs sont Mathilde Marchais, Isabelle Gilbert, Alxandre Bastien,
Karen E. Neonene, Edouard W. Khandjian, Robert S. Viger et Claude Robert qui ont tous
grandement contribué à ce projet de recherche.
xiiiIntroduction
L’ovocyte, une cellule très particulière de l’organisme d’un mammifère, est encore très
méconnue. La qualité ovocytaire est influencée par plusieurs facteurs en débutant par la
folliculogenèse, l’ovogenèse, la structure et la morphologie de l’ovocyte et la contribution de
l’ovocyte pour un développement embryonnaire de qualité. Ces divers facteurs influencent
la compétence de l’ovocyte à compléter son développement afin de former un embryon
viable. À ce jour, de nombreux mécanismes menant à la compréhension des éléments
influençant la compétence au développement d’un ovocyte sont très peu connus.
Le dogme central de la biologie moléculaire accepté assume que l’information génétique
supportée par l’ADN est transcrit en ARN ayant soit une fonction structurale, enzymatique
ou encore de transport. Les ARN dits messagers sont finalement traduits en protéines (Fu et
al., 2014). Pendant la croissance de l’ovocyte, des ressources sont emmagasinées dans le but
de soutenir les premières divisions cellulaires pendant l’absence de transcription nucléaire.
Au moment de l’activation du génome embryonnaire, la transcription est alors reprise.
Cependant, la période de silence transcriptionnel diffère selon l’espèce mammifère. En effet,
la demi-vie de ces transcrits est estimée de quelques heures à plusieurs jours selon les espèces
mammifères. Chez l’espèce murine, l’activité transcriptionnelle reprend lors du stade 1
cellule comparativement aux gros mammifères où la reprise a lieu au stade 4 cellules et 8
cellules chez le porc et le bovin respectivement (Schultz, 1993; Mateusen et al., 2005; (Frei
et al., 1989; Kopecny, 1989; Barnes and First, 1991). Les mécanismes permettant la
stabilisation des transcrits maternels et les différences entre les espèces durant cette période
ne sont pas connus.
En lien avec le domaine de l’épigénétique, des modifications chimiques réversibles peuvent
survenir sur l’ADN ayant d’importants rôles dans la régulation de la différenciation cellulaire
ainsi que dans le développement. Toutefois, une nouvelle voie a été explorée révélant que
des modifications chimiques peuvent également avoir lieu sur les ARN telles que l’ajout de
groupements méthyles (Fu et al., 2014).
1Ainsi, les travaux de recherche de ce mémoire ont pour objectif de caractériser le potentiel
de méthylation des ARNm des ovocytes ainsi que des tissus somatiques de différentes
espèces mammifères. Ce projet permettra d’acquérir des connaissances sur la gestion des
transcrits durant l’ovogenèse et sur la gestion de ceux-ci selon l’espèce animale. L’hypothèse
consiste à ce que l’ajout de groupements méthyles sur l’ARNm influence la capacité de
stabilisation des ARNm durant la période du silence transcriptionnel de l’ovocyte.
Le présent mémoire est divisé en deux chapitres. Dans un premier temps, le Chapitre 1
consiste en une revue de littérature présentant les connaissances actuelles à ce qui a trait du
développement d’un gamète femelle de qualité et les facteurs influençant sa compétence au
développement ainsi que de nouvelles perspectives en lien avec l’épitranscriptome. Le
second chapitre, présenté sous forme d’article scientifique, traitera plus spécifiquement des
différences de la gestion des ARNm entre plusieurs espèces mammifères ainsi que de la
caractérisation des protéines impliquées dans la méthylation de l’ARN durant l’ovogenèse.
2CHAPITRE 1 : Revue de littérature
1. Folliculogenèse
Le follicule ovarien est primordial pour les fonctions physiologiques principales de l’ovaire
(Paulini et al., 2014). C’est dans cette structure que l’ovogenèse se produit, c’est-à-dire, où
l’ovocyte se développe et se différencie. Chez les mammifères femelles, la folliculogenèse
débute avant la naissance de l’animal, soit au stade fœtal de sorte que de centaines de milliers
d’ovocytes se retrouvent dans l’ovaire lors de la naissance de la femelle (Paulini et al., 2014).
Les follicules ont plusieurs stades de développement, soit primordial, primaire, secondaire,
pré-antral, antral et finalement pré-ovulatoire (figure 1.1). Lors du développement prénatal
de l’ovaire, soit pendant la vie fœtale, la migration des cellules germinales dérivant de la
masse cellulaire interne du blastocyste, appelées ovogonies chez la femelle, prolifèrent dans
les cordons ovigères jusqu’à ce qu’elles atteignent la prophase méiotique marquant l’arrêt de
prolifération (Picton, 2001; Monniaux et al., 2009). Ensuite, la prophase méiotique se
poursuit pour s’arrêter au stade diplotène pendant que chaque ovocyte s’associe à une couche
de cellules somatiques, appelées les cellules de la granulosa, dans le but de former des
follicules primordiaux (Picton, 2001; Monniaux et al., 2009).
Figure 1.1 Développement folliculaire chez les mammifères. Adapté de Sánchez and Smitz
(2012).
3Ces follicules primordiaux constituent la réserve ovarienne du nombre d’ovocytes contenu
dans l’ovaire adulte puisque c’est à partir de celle-ci que les follicules effectueront leur
croissance jusqu’aux stades suivants de la folliculogenèse (Picton, 2001; Monniaux et al.,
2009). L’activation de follicules primordiaux est caractérisée par le changement des
quelques cellules de la granulosa entourant l’ovocyte à une forme cuboïde (Fair et al., 1997;
Li et Albertini, 2013).
Par la suite, les cellules de la granulosa prolifèrent afin de former au minimum deux couches
autour de l’ovocyte et la déposition initiale du matériel de la zone pellucide à lieu
caractérisant le stade secondaire de la folliculogenèse (Fair, 2003). Les cellules de la
granulosa prennent de l’expansion en proliférant tout au long de la maturation du follicule
jusqu’à ce que l’ovocyte ait terminé sa croissance (Li et Albertini, 2013). Pendant la
croissance de l’ovocyte, la zone pellucide se développe entre l’ovocyte et les cellules de la
granulosa au sein des follicules du stade pré-antral (Aerts and Bols, 2010a). Ensuite, le stade
antral de la folliculogenèse est caractérisé par l’apparition d’une cavité de fluide, appelée
l’antre, et chez la vache, c’est à ce stade que le follicule atteint plus de 250 cellules de la
granulosa (Li et Albertini, 2013). Chez ce même mammifère, l’antre se forme lorsque le
follicule a atteint 2% de son diamètre pré-ovulatoire et, à ce stade, tous les ovocytes ont une
zone pellucide développée (Fair et al., 1997; Aerts et Bols, 2010a). La majorité des follicules
activés atteignent ce stade, mais la plupart subiront l’apoptose par la suite (Aerts et Bols,
2010a).
Lorsque le follicule atteint le stade pré-ovulatoire, les cellules de la granulosa se distinguent
pour devenir les cellules du cumulus et les cellules murales (Li et Albertini, 2013). Les
cellules du cumulus sont les cellules qui sont en contact direct avec l’ovocyte formant une
couche unique contrairement aux cellules murales qui sont situées en périphérie de la limite
extérieure du follicule (Li et Albertini, 2013). Ces dernières sont impliquées dans la
production de stéroïdes hormonaux (Li et Albertini, 2013). Tout le processus de croissance
folliculaire du stade pré-antral jusqu’au stade pré-ovulatoire prend jusqu’à 85 jours chez
l’humain sans compter l’étape du recrutement tel que présenté en figure 1.2 (Gougeon, 1986).
4ATRÉSIE
120j
secondaire
10 j
1 couche cellules de
la granulosa
OVULATION
Figure 1.2 Stades et la durée (jours) de la folliculogenèse au sein de l’ovaire adulte chez
l'humain. Adapté de Gougeon (1986)
Chez la vache, la durée du développement folliculaire du stade primordial jusqu’au stade pré-
ovulatoire est estimée à 180 jours (Lussier et al., 1987). La folliculogenèse peut se diviser
en deux phases, soit la folliculogenèse basale et la folliculogenèse terminale (Monniaux
et al., 2009). La folliculogenèse basale se caractérise par la croissance du follicule
jusqu’au stade pré-antral en l’absence de l’hormone folliculostimulante (FSH) sécrétée
par l’hypophyse (Monniaux et al., 2009). C’est pendant cette phase qu’une grande partie
de l’ovogenèse s’effectue (Monniaux et al., 2009). Lors de la folliculogenèse terminale,
la FSH stimule la transition du follicule pré-antral à un follicule antral (Monniaux et al.,
2009). Il est donc dépendant de cette hormone durant cette phase jusqu’à sa maturation
5au stade pré-ovulatoire où la présence de l’hormone lutéinisante (LH) est requise
(Monniaux et al., 2009).
En effet, les récepteurs de l’hormone FSH n’ont pas été décelés dans les follicules
primordiaux (Combelles et al., 2004). C’est plutôt dans les follicules au stade secondaire que
ces récepteurs ont été observés sur les cellules de la granulosa avec l’apparition des cellules
de la thèque (Combelles et al., 2004). Finalement, l’ovulation a lieu lors de l’élévation du
taux de LH, appelé pic de LH, par la rupture de la membrane folliculaire et libérant ainsi
l’ovocyte (Li et Albertini, 2013).
1.1 Folliculogenèse in vitro
La culture in vitro de follicules ovariens permet d’étudier davantage le développement
ovocytaire. Durant ces dernières années, des progrès ont été réalisés sur la culture de
follicules primaires et secondaires chez plusieurs espèces (Xiao et al., 2015). Toutefois, isoler
et mettre en culture des follicules primordiaux s’avère être plus complexe en raison de leur
petite taille. La mise en culture des follicules à ce stade précoce permet d’étudier presque
l’intégrité du développement et de la maturation ovocytaire en plus d’être une voie pour de
nombreuses applications thérapeutiques (Telfer et McLaughlin, 2011). Pour ce faire,
diverses méthodologies ont été explorées dont celle de la mise en culture du tissu ovarien,
c’est-à-dire, le cortex étant donné que ce tissu contient principalement des follicules
primordiaux. Cette méthode permettrait de produire des ovocytes compétents pour des
techniques de reproduction assistée (Telfer et McLaughlin, 2011). Appliquée dans un autre
domaine, cette méthode permettrait également de garder la fertilité chez des jeunes femmes
atteintes d’insuffisances ovariennes causées par des traitements de radiothérapie ou de
chimiothérapie (Xiao et al., 2015). Xiao et al. (2015), ont mis en culture in vitro des follicules
ovariens provenant d’humains et les follicules au stade pré-antral qui se sont développés
jusqu’au stade antral et pour la première fois, ont atteint la compétence méiotique en se
développant jusqu’à la métaphase II. Ces grandes avancées sont prometteuses pour la
préservation de la fertilité chez les jeunes femmes ou les jeunes filles prépubères atteintes
d’un cancer (Xiao et al., 2015).
6En ce qui concerne le domaine animal, la culture in vitro de follicules du stade secondaire a
mené à une production d’embryons chez l’espèce caprine et ovine (Paulino et al., 2020). Pour
ce qui est de l’espèce bovine, les résultats se limitent en la formation de follicules au stade
antral à partir de follicules secondaires (McLaughlin et al., 2010; Paulino et al., 2020).
Ainsi, plusieurs études portent sur la supplémentation de milieu de culture des follicules
bovins in vitro afin d’obtenir des résultats permettant de maximiser les conditions de culture.
Ultimement, les ovocytes produits pourraient servir aux techniques de reproduction assistées.
En ce sens, Andrade et al. (2012) ont étudié l’effet de différentes doses d’acide ascorbique
sur des tranches de cortex ovarien bovin en culture in vitro. L’acide ascorbique est une
substance pouvant être ajoutée à un milieu de culture aux stades folliculaires précoces et
possède une propriété protective contre les dommages causés par les espèces réactives de
l’oxygène (Andrade et al., 2012). Cette étude a soulevé que l’ajout d’acide ascorbique à une
concentration de 50 g/ml au milieu de culture augmenterait le taux d’activation des
follicules primordiaux en plus de préserver leur viabilité tout au long des huit jours de culture
(Andrade et al., 2012).
D’autre part, McLaughlin et al. (2010) ont étudié l’impact de supplémenter un milieu de
culture de follicules ovariens bovin in vitro avec de l’activine. Ce composant est connu
comme étant impliqué dans l’activation folliculaire ainsi que dans leur développement au
stade pré-antral (McLaughlin et al., 2010). Leurs résultats suggèrent que l’ajout d’activine à
la culture de follicules bovins au stade pré-antral mène de façon significative à une
augmentation de la croissance folliculaire, de la prolifération des cellules du cumulus en plus
d’une augmentation du taux de formation de l’antre au sein du follicule (McLaughlin et al.,
2010).
Finalement, Paulino et al., (2020) ont étudiés l’impact de la supplémentation d’un milieu de
culture avec l’epidermal growth factor (EGF) ainsi que de la progestérone lors de culture in
vitro de follicules secondaires bovins isolés. L’EGF est connu pour avoir un effet favorable
sur l’expansion des cellules du cumulus ainsi qu’à l’amélioration de la maturation ovocytaire
et du taux de fécondité (Hsieh et al., 2007). Les conclusions tirées sont que l’EGF contribue
7significativement à la croissance folliculaire contrairement à la progestérone et qu’aucune
interaction positive entre ces deux hormones n’a été observée (Paulino et al., 2020). De plus,
aucune élévation du taux de survie des follicules n’a été notée avec les traitements de l’EGF,
de la progestérone et les deux combinées (Paulino et al., 2020).
2. Ovogenèse
L’ovogenèse se définit généralement comme étant le potentiel d’un gamète femelle de croître
en un ovocyte ayant la capacité d’être fécondé et de soutenir le développement de l’embryon
jusqu’au stade de blastocyste (Conti et Franciosi, 2018). Ce processus est étroitement lié avec
celui de la folliculogenèse (Monniaux et al., 2009). Tout comme la folliculogenèse,
l’ovogenèse débute au stade fœtal par la différenciation des cellules germinales primordiales,
se situant dans l’ovaire, en ovogonies qui sont les cellules souches spécifiques de l’ovaire
(Monniaux et al., 2009).
L’ovocyte croit jusqu’au stade de la prophase méiotique caractérisant la formation des
follicules primordiaux et ce stade est atteint de 75 à 80 jours post-conception chez
l’espèce bovine (Erickson, 1966). La prophase méiotique de l’ovocyte est composée des
étapes de la préleptotène, leptotène, zygotène, pachytène et diplotène (Fair, 2003). Ce
dernier stade est marqué par l’arrêt de la première division méiotique survenant près de 170
jours post-conception chez la vache (Baker and Hunter, 1978). Le follicule primordial
contient donc un ovocyte stagné à la prophase I au stade diplotène contenant quatre copies
de cellules d’acide désoxyribonucléique (ADN) composées de deux jeux de chromosomes
diploïdes (4N) tel que présenté à la figure 1.3 (Li et Albertini, 2013). À ce stade, le noyau de
l’ovocyte est connu sous l’appellation de germinal vesicle (GV) (Tan et al., 2009). Ce ne
sera que lors de la puberté, que la croissance de l’ovocyte se poursuivra déclenchée par la
FSH (Monniaux et al., 2009).
8Figure 1.3 Maturation d’un ovocyte de mammifère.
Chez l’espèce bovine, la croissance de l’ovocyte reprend lorsque le follicule contient
minimalement 40 cellules de la granulosa et connaît différents niveaux de modifications de
remodelage de la chromatine au sein du GV (Tan et al., 2009; Aerts et Bols, 2010a). La
croissance de l’ovocyte cesse lorsque le follicule atteint un diamètre de 3 mm pour stagner à
une taille de 120 à 130 m tandis que le follicule poursuit sa croissance jusqu’à une taille se
situant entre 15 et 20 mm de diamètre au stade pré-ovulatoire tel qu’illustré à la figure 1.4
(Fair, 2003).
9Diamètre
Diamètre follicule (mm)
ovocyte
Figure 1.4 Relation entre la taille des follicules et ovocytes durant leur
développement chez la vache. Adapté de Fair (2003).
La reprise de la maturation de l’ovocyte sera déclenchée par le pic de LH qui stimule la
rupture de la vésicule germinale (GVBD) et tout le nucléoplasme qui était contenu dans cette
vésicule se déverse dans le cytoplasme de l’ovocyte (Monniaux et al., 2009; Aerts et Bols,
2010a). C’est également durant cette période que la chromatine de la vésicule germinale est
décondensée et que des activités de transcription ont lieu tel que rapporté par Conti et
Franciosi (2018). Les chromosomes se condensent et un fuseau méiotique se forme
caractérisant ainsi la reprise de la méiose I où les chromosomes se lient et migrent en
périphérie (Li et Albertini, 2013). Ce processus induit la formation d’un domaine cortical
d’actomyosine (Li et Albertini, 2013). Après la méiose I, une première extrusion d’un corps
polaire a lieu (Li et Albertini, 2013). Ensuite, l’ovocyte débute la méiose II pour
s’interrompre, encore une fois, à la métaphase II (Li et Albertini, 2013). Lorsque cet ovocyte
10sera expulsé par la rupture du follicule, dite ovulation, la méiose II sera reprise seulement s’il
y a fécondation avec un spermatozoïde (Li et Albertini, 2013). Uniquement les follicules
exprimant des récepteurs de LH en grand nombre à la surface des cellules de la granulosa
pourront répondre au pic de LH et ovuler (Monniaux et al., 2009). Lors de la fécondation,
une deuxième extrusion d’un corps polaire se produira et il y aura séparation des chromatides
sœurs laissant un chromatide femelle issu de l’ovocyte et un chromatide mâle provenant du
spermatozoïde qui formeront le zygote (Li et Albertini, 2013). Une fois la méiose II terminée,
l’ovogenèse se conclut, mais si aucune fécondation n’a lieu, l’ovocyte dégénère par apoptose
(Li et Albertini, 2013).
2.2 Culture in vitro
La culture in vitro consiste à mettre les ovocytes en maturation dans des milieux de culture
artificiels. La fécondation sera également produite de façon in vitro en laboratoire. Lorsque
cette méthodologie est utilisée, il est primordial de bien contrôler l’environnement, par les
milieux de culture, la température et l’humidité pour les ovocytes, car un moindre désordre
a un impact non négligeable sur la maturation de l’ovocyte en culture (Filatov et al., 2017).
Un défi de grande importance dans les techniques de reproduction assistées est de mettre
l’ovocyte dans les bonnes conditions afin qu’il acquière les compétences développementales
nécessaires à la fécondation (Moussa et al., 2015). En effet, l’application des
gonadotrophines (FSH et LH) ainsi que d’autres additifs permettent de tenter de recréer,
jusqu’à une certaine mesure, les conditions in vivo en culture in vitro tout en gardant en tête
qu’il est impossible de complètement recréer un environnement in vivo en laboratoire.
Toutefois, l’exposition de ces structures à des conditions non physiologiques provoque
régulièrement un développement ovocytaire irrégulier menant à un embryon anormal (del
Collado et al., 2017). En effet, pendant la production d’embryons bovins en condition in
vitro, certaines anomalies métaboliques se produisent dont principalement l’accumulation
anormale de lipides au sein du blastocyste (del Collado et al., 2017). Une haute teneur en
lipides dans le blastomère provoque des impacts négatifs tels qu’un potentiel réduit de
cryoconservation et un faible taux de survie embryonnaire après sa décongélation (del
Collado et al., 2017). Ces effets peuvent ensuite mener à des taux de gestation plus faibles et
11des pertes embryonnaires ce qui doit être évité dans les élevages (del Collado et al., 2017).
Ce type de désordre métabolique se produit lors de la maturation in vitro des ovocytes et non
lorsqu’ils sont produits en milieu in vivo (del Collado et al., 2017). Cette affirmation
soutiendrait le fait que les ovocytes sont de qualité supérieure lorsqu’ils sont produits sous
des conditions physiologiques et non sous des conditions artificielles.
3. Production d’ovocytes in vivo et cycle œstral bovin
Dans une optique d’améliorer le potentiel génétique d’un troupeau de vaches laitières, il est
certain que d’obtenir une seule descendance par gestation de neuf mois n’est pas un processus
rapide. C’est ainsi qu’il y a eu un engouement pour la technologie de reproduction assistée
de la part des producteurs.
Dans un premier temps, le cycle œstral bovin se caractérise par deux à trois vagues
folliculaires et seule la dernière cohorte de follicules résultera en une ovulation (Fair, 2003;
Aerts and Bols, 2010b). Une vague folliculaire évolue en quatre phases (Sirard et al., 2018).
En premier, la phase de recrutement où un groupe de follicules se développe dû à
l’augmentation du taux de FSH (Sirard et al., 2018). Ensuite vient la phase de sélection où la
diminution du taux de FSH mène à la sélection d’un follicule dominant ayant acquis des
récepteurs LH avant les autres follicules de la même vague (Sirard et al., 2018). Ce follicule
dominant sera davantage dépendant de la LH que de la FSH par ses récepteurs qui
maintiennent sa croissance tout en évitant l’atrésie (Sirard et al., 2018). Ce processus
s’effectue lors de la phase dite de dominance (Sirard et al., 2018). Les autres follicules cessent
de croître et terminent en atrésie (Sirard et al., 2018). Finalement, la phase de régression se
produit lorsque le taux de LH n’est pas suffisamment élevé et qu’il y a présence de
progestérone sécrétée par les follicules (Sirard et al., 2018). Le follicule dominant perd sa
dominance et se verra décroitre pour ultimement être en atrésie ce qui activera le recrutement
d’une nouvelle vague folliculaire (Fair, 2003; Sirard et al., 2018). Chez l’espèce bovine, un
seul follicule dominant par vague folliculaire est apte à atteindre le stade de l’ovulation. De
ce fait, plus de 99% des follicules primordiaux présents au sein des ovaires de la génisse à la
naissance terminent en atrésie (Aerts et Bols, 2010b). Lorsque le follicule dominant est près
12d’atteindre sa taille maximale étant de plus de 14 mm, plusieurs changements structurels au
sein de l’ovocyte se produisent (Fair, 2003).
Toutefois, il est possible de produire plusieurs follicules de type dominant à l’aide d’un
soutien exogène de FSH (Sirard et al., 2006). Cette technique se nomme la superovulation et
elle consiste à stimuler l’ovaire à l’aide d’un traitement FSH. D’abord, le follicule dominant
de la vague est aspiré afin d’induire une nouvelle vague folliculaire (Nivet et al., 2012).
L’ovaire est ensuite stimulé avec un protocole comportant plusieurs injections de FSH (Nivet
et al., 2012).
Les ovocytes produits peuvent être récoltés afin de les féconder in vitro. Une autre façon est
de les féconder in vivo par insémination artificielle puis de les récolter pour les implanter
dans des femelles dites receveuses préparées à cet issu. Lorsque la méthodologie de la
fécondation in vitro est utilisée, plusieurs études suggèrent qu’un temps de coasting à la suite
d’une injection de FSH influence la qualité ovocytaire (Sirard et al, 2006 ; Nivet et al., 2012).
En effet, il a été démontré que la diminution de la FSH est importante dans l’acquisition des
compétences de l’ovocyte et que le prélèvement des ovocytes dans une fenêtre de 47 à 61
heures après la dernière poussée de FSH est la période optimale afin d’obtenir des ovocytes
de qualité (Nivet et al., 2012). Cette fenêtre de temps simulerait la phase plateau du follicule
(Sirard et al., 2006).
4. Facteurs affectant la folliculogenèse et l’ovogenèse
Une observation des mécanismes et des structures de l’ovocyte est importante non seulement
pour la compréhension de leur contribution à la qualité ovocytaire, mais également pour
évaluer les effets sur ceux-ci des diverses techniques de reproduction assistée. En effet, la
maturation in vitro et la cryoconservation d’ovocytes font l’objet de nombreuses études. En
fait, les organites cytoplasmiques ovocytaires subissent différentes altérations qui reflètent
les processus physiologiques et le développement ovocytaire (Paulini et al., 2014). Plusieurs
structures et molécules jouent des rôles clés dans le fonctionnement de l’ovocyte tels que les
cellules du cumulus (figure 1.5), les projections transzonales (PTZs) et le pyruvate.
13Figure 1.5 Coupe histologique d'un follicule ovarien au stade antral chez la souris.
Adapté de Monniaux et al. (2009).
4.1 Cellules du cumulus
Les cellules du cumulus sont en contact direct avec l’ovocyte via les PTZs au sein du follicule
ovarien (Li et Albertini, 2013). Ces cellules sont primordiales à l’ovocyte en fournissant des
nutriments et des métabolites en plus de métaboliser l’énergie consommée par l’ovocyte (del
Collado et al., 2017). Elles ont également un grand rôle dans le contrôle de fonctions vitales
telles que l’arrêt et la reprise méiotique de l’ovocyte (Coticchio et al., 2015). En effet, c’est
par leur production d’énergie dans le complexe ovocyte cumulus (COC) qu’elles sont
essentielles pour maintenir la méiose pendant la maturation méiotique de l’ovocyte
(Coticchio et al., 2015). Les cellules du cumulus transfèrent des nutriments comme le
pyruvate et le lactate à l’ovocyte (Paczkowski et al., 2013). Cette communication avec les
cellules du cumulus est indispensable puisqu’il est connu que la fécondation d’un ovocyte
dénudé est altérée et que peu d’embryons sont produits à partir de ceux-ci (Gilchrist, 2011).
De plus, Matsuda et al. (2012) suggèrent que ces cellules initieraient l’atrésie folliculaire
chez les mammifères puisque les cellules du cumulus ont été observées comme population
14Vous pouvez aussi lire

















































