Évaluation du potentiel de compétition entre le caribou migrateur et le boeuf musqué au Nunavik - Mémoire Alexis Brodeur Maîtrise en biologie ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Évaluation du potentiel de compétition entre le caribou
migrateur et le bœuf musqué au Nunavik
Mémoire
Alexis Brodeur
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Alexis Brodeur, 2021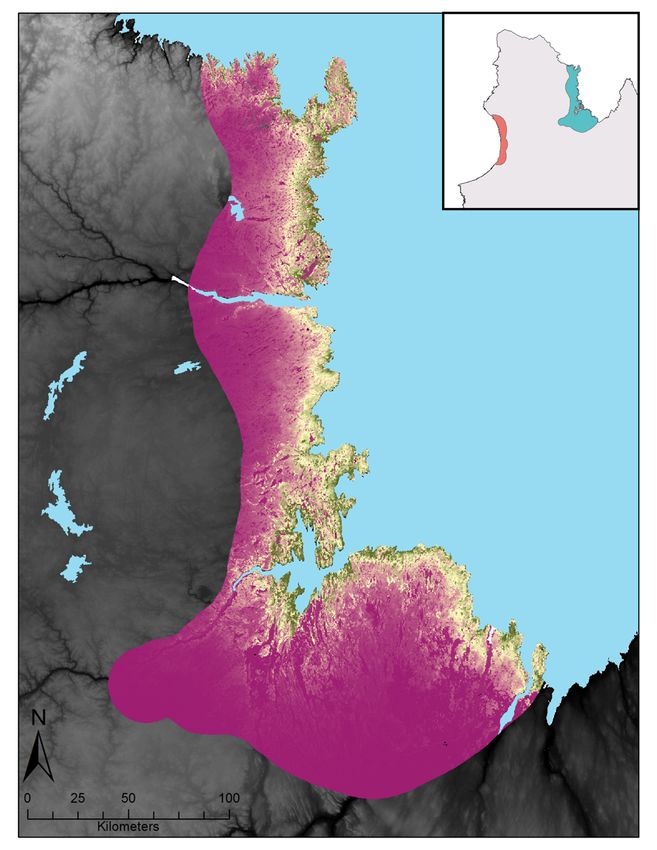
Évaluation du potentiel de compétition entre le caribou
migrateur et le bœuf musqué au Nunavik
Mémoire
Alexis Brodeur
Sous la direction de :
Steeve D. Côté, directeur de recherche
Joëlle Taillon, codirectrice de recherche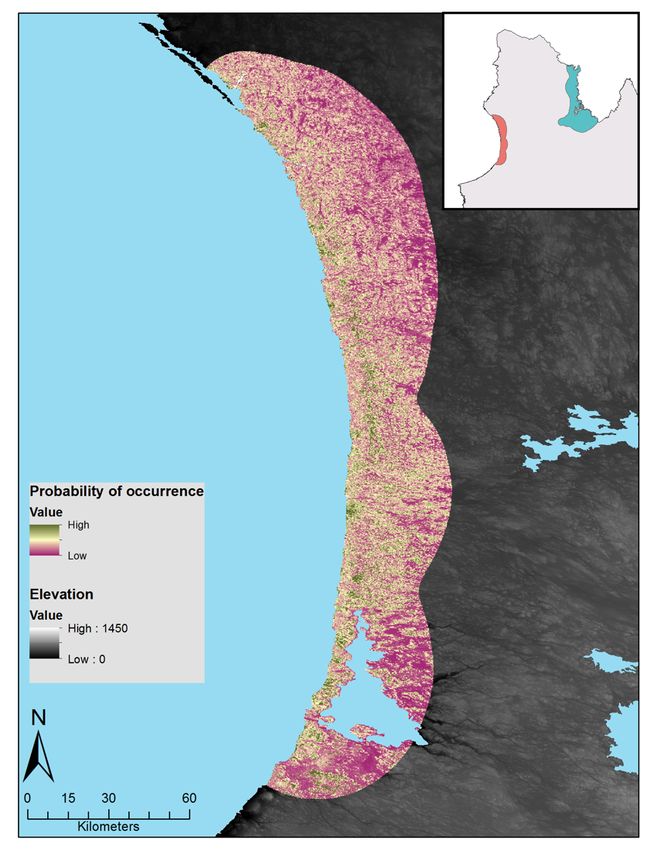
Résumé
Dans le Nord-du-Québec et au Labrador, les troupeaux de caribous migrateurs Rivière-George et Rivière-aux-
Feuilles sont en déclin, respectivement depuis les années 1990 et 2000. Plus particulièrement, le déclin marqué
du troupeau Rivière-aux-Feuilles qui a coïncidé avec la croissance de la population de bœuf musqué, une
espèce introduite dans le Nord-du-Québec, a mené les communautés inuites à s’interroger sur le potentiel de
compétition entre les deux espèces. En réponse à ce questionnement, nous avons combiné trois approches
pour évaluer le potentiel de compétition interspécifique : (1) quantifier le chevauchement spatial, (2) comparer
les patrons de sélection d’habitats des deux espèces dans leur aire de répartition commune et (3) évaluer le
chevauchement des régimes alimentaires. Pour ce faire, nous avons utilisé des données provenant de suivis
télémétriques et récolté des échantillons de fèces pour les analyser avec la méthode du codage à barre d’ADN.
À l’échelle annuelle, le chevauchement spatial était faible et avait lieu principalement au printemps sur la côte
de la baie d’Hudson et à l’été sur la côte de la baie d’Ungava. Dans leur aire de répartition commune, le bœuf
musqué sélectionne les milieux arbustifs et productifs situés près des côtes alors que le caribou sélectionne des
milieux de toundra avec substrat rocheux et des habitats de faible altitude. À une échelle spatiale plus fine, cela
fait en sorte que les probabilités de co-occurrence sont faibles. Toutefois, notre analyse du régime alimentaire
a montré des similarités, puisque les familles de plantes détectées les plus fréquemment (Ericacea, Salicaceae,
Betulaceae) étaient les mêmes pour les deux espèces à l’été et à l’hiver. Dans leur ensemble, ces résultats
suggèrent que le potentiel de compétition est faible entre le caribou migrateur et le bœuf musqué dans le Nord-
du-Québec. Cependant, plusieurs facteurs dont l’expansion possible de l’aire de répartition du bœuf musqué et
les interactions avec les prédateurs, dans un contexte de changement climatique, pourraient éventuellement
influencer les interactions entre ces deux espèces d’ongulés.
ii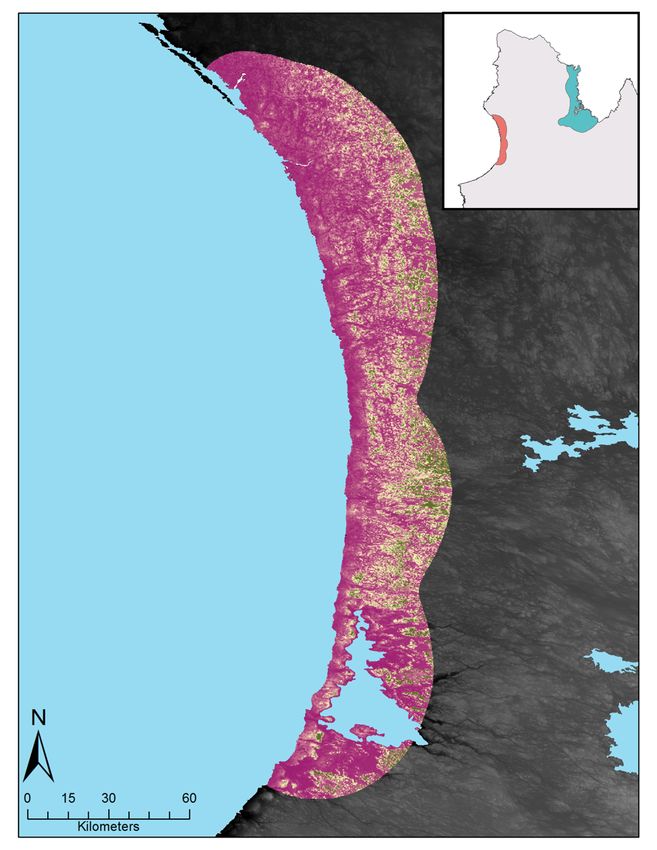
Abstract
In northern Quebec and Labrador, the Rivière-George and Rivière-aux-Feuilles caribou herds have been
declining since the 1990s and 2000s, respectively. In particular, marked decline of the Rivière-aux-Feuilles
caribou herd coinciding with the demographic growth of muskoxen, an introduced species in northern Quebec,
has led local Inuit communities to question the potential for interspecific competition. In response to these
concerns we used three methods to evaluate the potential for interspecific competition at different scales: (1) we
quantified spatial overlap, (2) we compared both species habitat selection patterns in their overlapping range
and (3) we evaluated diet overlap. We used telemetric data and collected fecal samples of both species for DNA
barcoding analysis. At the annual scale, spatial overlap was low and limited in time, mostly in spring on the coast
of Hudson Bay and in summer on the coast of Ungava Bay. Within their overlapping range, migratory caribou
and muskoxen had mostly different habitat selection patterns, particularly in regard to habitat types and proximity
to the coast. At a finer spatial scale, this led to low probability of co-occurrence across both coastal areas. Our
dietary analysis of fecal samples suggested that both species had a similar diet based on the frequency of the
most detected plant families (Ericaceae, Salicaceae et Betulaceae) in summer and winter. Combined, our results
suggest that the potential for competition between migratory caribou and muskoxen is low in our study area.
However, several factors such as the possible expansion of the muskoxen range and interactions with predators,
in the context of climate change, could potentially influence the interactions between these two ungulate species.
iii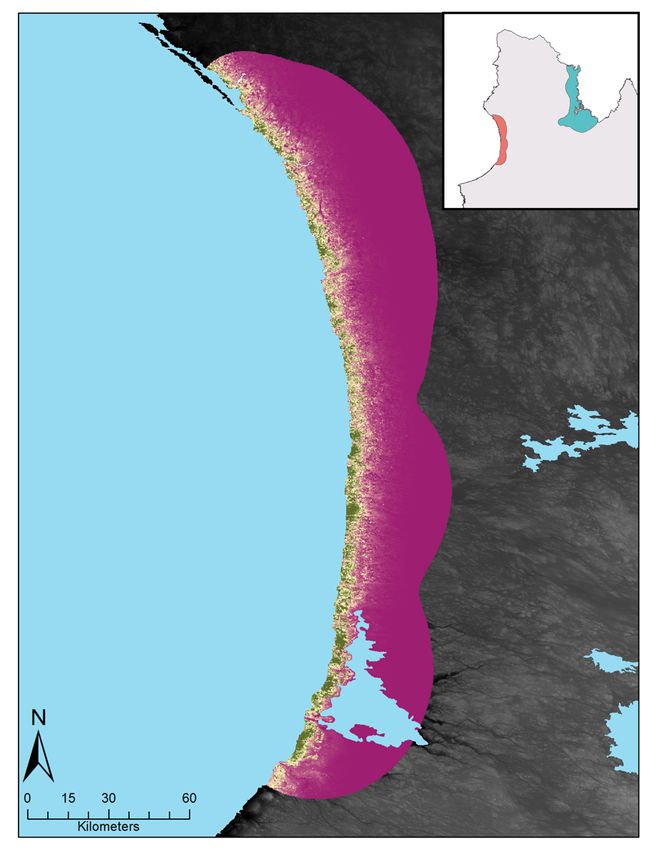
Table des matières
Résumé ................................................................................................................................................................ii
Abstract................................................................................................................................................................ iii
Table des matières ..............................................................................................................................................iv
Liste des tableaux ................................................................................................................................................ v
Liste des figures...................................................................................................................................................vi
Remerciements................................................................................................................................................... vii
Avant-propos .......................................................................................................................................................ix
Introduction .......................................................................................................................................................... 1
Contexte théorique .......................................................................................................................................... 1
L’étude de la compétition interspécifique en milieu naturel ............................................................................. 2
Compétition pour l’espace : utilisation et sélection d’habitat ........................................................................... 3
Compétition pour les ressources alimentaires ................................................................................................ 4
Contexte de l’étude : potentiel de compétition entre le caribou migrateur et le bœuf musqué ....................... 5
Historique de la problématique entourant la compétition entre le caribou et le bœuf musqué ....................... 6
Écologie du caribou migrateur au Nord-du-Québec ........................................................................................ 7
Écologie du bœuf musqué au Nord-du-Québec.............................................................................................. 8
Objectifs .......................................................................................................................................................... 9
Chapitre 1: Investigating the potential for competition between migratory caribou and introduced muskoxen . 11
1.1 Résumé ................................................................................................................................................... 11
1.2 Abstract ................................................................................................................................................... 12
1.3 Introduction.............................................................................................................................................. 13
1.4 Methods................................................................................................................................................... 14
Study area................................................................................................................................................. 14
Study populations ..................................................................................................................................... 16
Animal capture and monitoring ................................................................................................................. 17
Space use ................................................................................................................................................. 17
Habitat selection ....................................................................................................................................... 18
1.5 Results .................................................................................................................................................... 21
Space use ................................................................................................................................................. 21
Habitat selection ....................................................................................................................................... 22
Diet............................................................................................................................................................ 27
1.6 Discussion ............................................................................................................................................... 29
Acknowledgements ....................................................................................................................................... 33
1.7 References .............................................................................................................................................. 34
1.8 Supplementary material .......................................................................................................................... 39
Conclusion ......................................................................................................................................................... 44
Chevauchement spatial ................................................................................................................................. 44
Sélection d’habitat ......................................................................................................................................... 46
Régime alimentaire ....................................................................................................................................... 47
Évaluation du potentiel de compétition.......................................................................................................... 48
Limites de l’étude .......................................................................................................................................... 49
Perspectives de recherche ............................................................................................................................ 52
Bibliographie ...................................................................................................................................................... 53
iv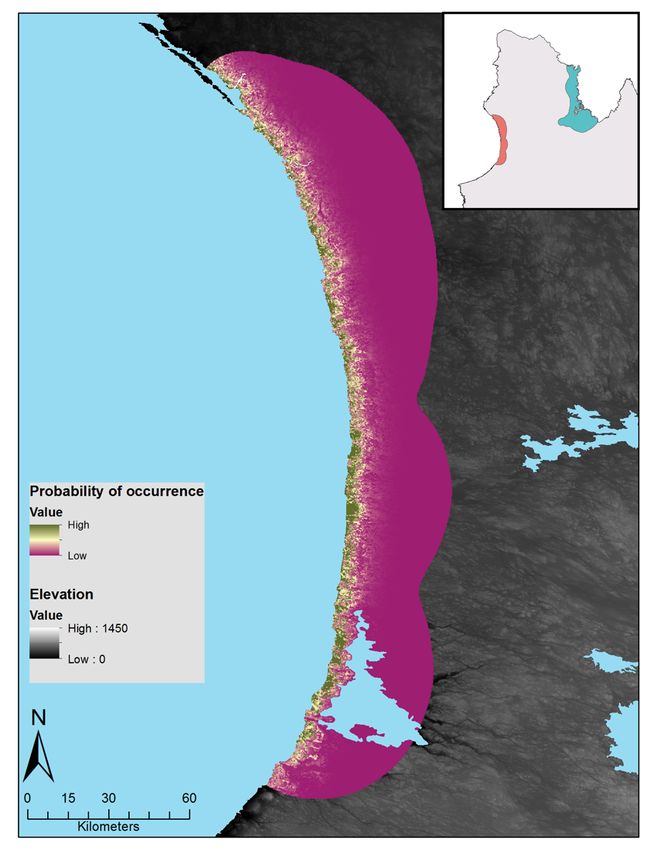
Liste des tableaux
Tableau 1 : Contribution des auteurs de l’article scientifique présenté dans le mémoire. ...................................ix
Table 2 : Caribou and muskoxen telemetry locations recorded in each study area per year. The number of
individuals is indicated in parentheses. ............................................................................................................. 23
Table 3 : Candidate models and model selection results for spring (Hudson Bay; 2017-2018) and summer
(Ungava Bay; 2018) habitat selection of the Rivière-aux-Feuilles caribou herd and muskoxen in Northern
Québec. We chose the most parsimonious models for each species and season (lower AICc value and ∆AICc
> 2) to predict co-occurrence probabilities on both species overlapping range. Models are described in table S2
in supplementary material.................................................................................................................................. 24
Table 4 : Resource selection estimates ± [95 % confidence intervals] for the Rivière-aux-Feuilles caribou herd
and muskoxen in spring (Hudson Bay ; 2017-2018) and summer (Ungava Bay ; 2018) in Northern Québec... 26
Table S1 : Habitat class description (based on Leboeuf et al. (2018)) and availability (%) in each study area. 40
Table S2 : Candidate mixed-effects models describing seasonal resource selection by migratory caribou and
muskoxen on their overlapping range in Northern Québec (Spring : Hudson Bay study area and Summer :
Ungava Bay study area). ................................................................................................................................... 43
v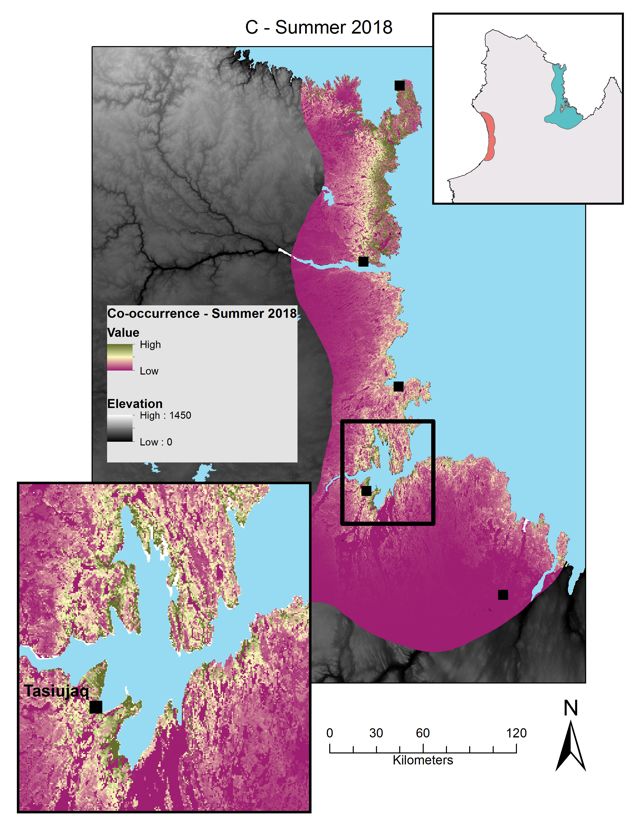
Liste des figures
Figure 1 : Annual (dashed outline), winter (light gray) and summer (light green) ranges of the Rivière-aux-Feuilles
caribou herd, and muskoxen ranges in Hudson Bay (red) and Ungava Bay (blue) in northern Québec, Canada.
We delineated annual caribou range using a 100% minimum convex polygon and seasonal caribou ranges using
95% minimum convex polygons of telemetry data from 2015 to 2019. Muskoxen range delineation is described
in the methods section (study area)................................................................................................................... 16
Figure 2 : Annual overlap (% of area) between caribou and muskoxen in northern Québec from 2015 to 2019
(*caribou data collected until November 2019). We measured spatial overlap as the proportion of the annual
migratory caribou range overlapping the muskoxen ranges in Northern-Quebec. Caribou range varied annually
while the muskoxen range was kept constant throughout the period. ............................................................... 22
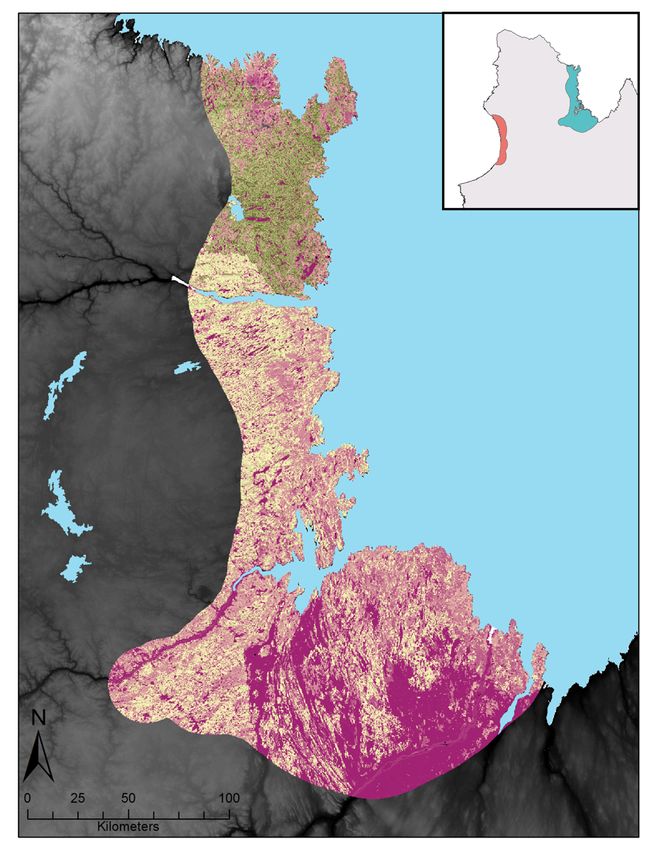
Figure 3 : Relative probabilities of spring and summer caribou-muskoxen co-occurrence in northern Québec.
Spring co-occurrence in the Hudson Bay region in (A) 2017 and (B) 2018 and summer co-occurrence in the
Ungava Bay region in (C) 2018. Relative probabilities were estimated by multiplying caribou relative probabilities
of occurrence and muskoxen relative probabilities of occurrence. Each map color scale is derived from data
standard deviation. ............................................................................................................................................ 27
Figure 4 : Heat map illustrating the frequency of occurrence of each plant families detected in the fecal samples
of caribou and muskoxen in northern Québec. Mosses families were grouped. The darker the color, the higher
the frequency of occurrence. ............................................................................................................................. 28
Figure 5 : Non-metric multidimensional scaling (NMDS) of presence-absence based on Bray-Curtis dissimilarity
of samples. Dashed (muskox) and grey (caribou) ellipses in panel A represent 95% confidence intervals. Only
families detected in more than 5 samples were included in the analyses. Symbols distinguish caribou (open) and
muskoxen (filled), and seasons: winter (squares), late winter (circles) and summer (triangles). ...................... 29
Figure S1 : Caribou telemetry locations in spring (2017 to 2019) overlapping the Hudson Bay study area (red).
The overlapping locations were used in the resource selection analyses. We delineated the Hudson Bay study
area by generating a 95 % kernel polygon around all muskox observations compiled by the Government of
Québec since 1988 (MFFP, unpublished data). ................................................................................................ 39
Figure S2 : Caribou telemetry locations in summer (2017 to 2019) overlapping the Ungava Bay study area (blue).
The overlapping locations were used in the resource selection analyses. We delineated the Ungava Bay study
area by creating a 95 % kernel polygon around all muskox observations recorded during an extensive aerial
survey conducted in March 2019 (MFFP, unpublished data). ........................................................................... 39
Figure S3 : Map of the fecal samples collected for the dietary analyses using DNA metabarcoding ................ 41
Figure S4 : Relative probability of occurrence of caribou (top panels) and muskoxen (bottom panels) during
spring and summer. These maps were produced using variables identified in the top model for each species
(table 3). Each map color scaled is derived from data standard deviation. ....................................................... 42
vi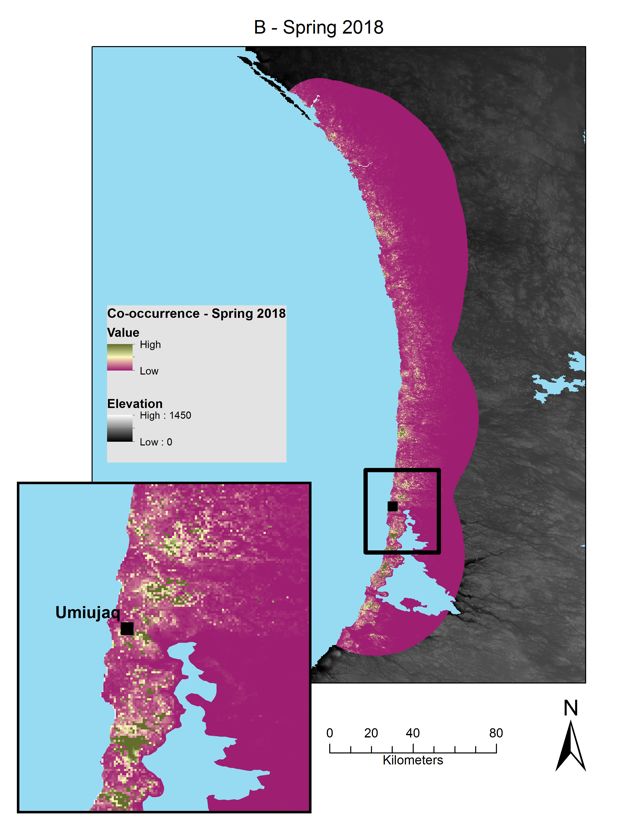
Remerciements
La réalisation d’un projet de maîtrise n’est jamais la réalisation que d’une seule personne. Je tiens à remercier
tous ceux qui, de près ou de loin, auront contribué à la réalisation de ce projet. Tout d’abord, merci à mon
directeur de recherche, Steeve D. Côté, de m’avoir donné la chance de venir étudier à Québec et de faire partie
de la grande famille du labo Côté-Tremblay. Mon intérêt pour le projet et pour l’équipe m’aura amené à
déménager considérablement plus loin que ce que j’avais envisagé quand j’ai décidé de poursuivre à la maîtrise,
une décision que je prendrais à nouveau demain matin. Je serai toujours reconnaissant d’avoir pu travailler sur
ce projet de maîtrise. De plus, les opportunités de travail sur le terrain et les présentations dans plusieurs
congrès, ici comme à l’étranger, ont été très formatrices et surtout, seront des souvenirs pour la vie. Je tiens
également à remercier Joëlle Taillon, ma codirectrice, pour son support tout au long de mon parcours. Tu as su
me poser les bonnes questions au bon moment et pour que je puisse améliorer mon projet et pour que je puisse
acquérir les connaissances et l’expertise qui me seront utiles dans ma vie professionnelle. Merci aussi de m’avoir
donné l’occasion de partager les résultats de mon projet de maîtrise à de nombreuses occasions et pour
l’opportunité d’aller sur le terrain capturer des bœufs musqués.
Je voudrais aussi remercier Mathieu Leblond pour ses judicieux conseils et sa grande disponibilité. Tu n’as
jamais été découragé de répondre à mes questions et faire de nombreuses relectures. Ton support a
grandement contribué à la réussite de mon projet de maîtrise. J’ai beaucoup apprécié travailler avec toi et
j’espère que ça pourra continuer dans le futur. Merci de m’avoir fait sourire avec des commentaires parfois très
drôles dans le manuscrit, peut-être même sans l’avoir réalisé, quand j’en avais besoin.
Merci à Julien et Cathy pour votre grande disponibilité à répondre à mes questions et à tous problèmes
logistiques liés à l’envoi de colis et de données télémétriques. Je voudrais aussi remercier tous ceux qui ont
contribué au projet pour le Ministère des Forêts, de la Faune et des Parcs et tout particulièrement Vincent
Brodeur.
Je tiens à remercier spécialement Delphine, Évelyne, Florent, Julien, Martin, Michaël et Rachel pour la relecture
de différentes parties de ce document, de résumés et pour les sessions de brainstorming ! Merci beaucoup à
Sabrina, Michaël et Martin, vous êtes pour moi de grandes sources d’inspiration et l’incarnation du scientifique
moderne. Votre aide a réellement été inestimable et j’ai grandement apprécié travailler avec vous.
Merci à tous membres du labo Côté-Tremblay et plus particulièrement Florent, Rachel, Évelyne, Delphine, Linda,
Michaël, Catherine, Barbara, Sophiane, Jérémie, Olivier, Sabrina, Émilie, Laurent, Béatrice, Mathilde, Frédéric,
Marianne, Martin, Antoine, Maël. Les bières, les discussions du midi, les parties de aki et les soirées karaoké
ont définitivement un rôle à jouer dans la complétion de mon projet ! Mention spéciale groupe des maitriseux,
vii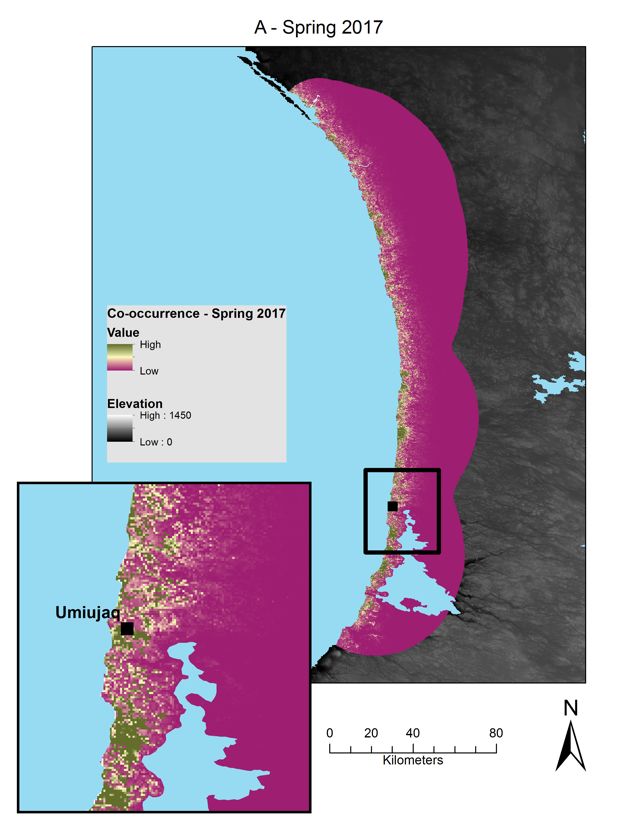
merci pour le support en temps réel. Merci à la gang du 881 de m’avoir hébergé à la fin de mon séjour à Québec.
Merci à tous ceux qui ont déjà arrosé mes plantes à tous ceux avec qui j’ai fait du sport à Québec.
Je voulais également prendre le temps de remercier mes parents, mes sœurs, membres de la famille et de la
belle-famille pour votre présence, votre support et pour tous les encouragements au cours des dernières
années. Ce n’était pas une décision facile à prendre d’aller à Québec, mais ce fut définitivement plus facile et
agréable grâce à vous. Merci, Maman, de m’avoir cuisiné des plats à emporter, la valeur d’un chili à réchauffer
le lundi midi est inestimable. Merci à mes parents pour votre support tout au long de mon parcours scolaire.
Sara-Maude et Anne-Sophie, j’espère que ça ne vous aura pas trop fait peur ; c’est OK d’être encore à l’école
à 26 ans… Merci à vous deux pour votre joie de vivre. Merci, François et Sonia (et Claudia, Gabriel, Gabrielle,
Raffael, Camille et Daniel), de m’avoir accueilli dans la famille et supporté toutes ces années, c’est un bonheur
de faire partie de la famille. Merci spécial pour l’hébergement et l’accueil (en tout temps, mais surtout le vendredi
soir), ce fut grandement apprécié.
Finalement, merci Marie-Lou. Si je suis arrivé à réaliser ce projet et à écrire ce mémoire, c’est beaucoup grâce
à toi. Merci d’avoir fait le trajet vers Québec si souvent, même si ça voulait dire de côtoyer des étrangers dans
un amigo. Merci de m’avoir soutenu dans les hauts et les bas et au travers des nombreuses périodes de remises
en question. Je pense que tu aurais pu écrire ce mémoire à ma place à force de m’avoir entendu présenter mon
projet ! J’aurais pu écrire un roman ici alors je vais simplement te dire merci pour tout. Et le tout dernier mais
non le moindre, Albert, mon compagnon de confinement sans qui le travail à la maison aurait été beaucoup plus
difficile.
viii
Avant-propos
Ce mémoire de maîtrise sur l’étude du potentiel de compétition interspécifique entre le caribou migrateur et le
bœuf musqué a comme objectif de répondre à une préoccupation locale et d’apporter des informations qui
pourront contribuer à la gestion de ces deux espèces dans le Nord-du-Québec. Le mémoire comporte trois
sections : une introduction générale, un chapitre principal sous la forme d’un article scientifique rédigé en anglais
et une conclusion générale.
L’article scientifique présenté ici comme chapitre principal sera soumis à une revue scientifique. J’ai réalisé les
analyses statistiques, l’interprétation des résultats et la rédaction de l’article en tant que premier auteur en
collaboration avec ma codirectrice Joëlle Taillon (biologiste au Ministère des Forêts, de la Faune et des Parcs),
Mathieu Leblond (chercheur à Environnement et Changement Climatique Canada), Vincent Brodeur (biologiste
au Ministère des Forêts, de la Faune et des Parcs) et mon directeur Steeve D. Côté (professeur titulaire au
Département de biologie de l’Université Laval). La contribution des auteurs au projet est présentée dans le
tableau 1.
Tableau 1 : Contribution des auteurs de l’article scientifique présenté dans le mémoire.
A. BRODEUR M. LEBLOND J. TAILLON V. BRODEUR S.D. CÔTÉ
CONCEPTION DU PROJET X X X X
FINANCEMENT X X X X
COLLECTE DE DONNÉES X X X X
ANALYSE STATISTIQUES X
INTERPRÉTATION X X X X
DES RÉSULTATS
RÉDACTION X
RÉVISION X X X X X
APPROBATION FINALE X X X X X
ixIntroduction
Contexte théorique
En milieu naturel, la dynamique de population peut notamment être influencée par des mécanismes de
régulation dépendants de la densité (Solberg et al. 2001), par des variations ponctuelles ou récurrentes des
conditions environnementales (Chan et al. 2005, Frederiksen et al. 2008) ou par des interactions avec d’autres
espèces (Gurnell et al. 2004). Parmi ces interactions, la compétition interspécifique est le sujet d’un nombre
croissant d’études, entre autres à la suite d’introductions d’espèces (O’Brien et al. 2019) et du déplacement des
aires de répartition en lien avec les changements climatiques (Alexander et al. 2015, Fitt et Lancaster 2017) et
anthropiques (Brown 2007, Buchmann et al. 2013). La compétition interspécifique peut engendrer des
répercussions sur des individus, telles qu’une augmentation du niveau de stress (Formenti et al. 2018), un
déplacement du domaine vital (Focardi et al. 2006), une détérioration de la condition corporelle (Wauters et al.
2002), une diminution de la capacité reproductive (Wauters et al. 2000) ou une augmentation de la probabilité
de mortalité (Merkle et al. 2009). Ces conséquences peuvent, ensuite, entrainer des effets négatifs à l’échelle
de la population (Gurnell et al. 2004).
Les mécanismes de compétition entre deux espèces peuvent être directs ou indirects. Plusieurs conditions
doivent être rencontrées pour que des individus de différentes espèces soient en compétition : (1) les espèces
doivent vivre en sympatrie, (2) les espèces doivent partager plusieurs ressources (p. ex. nourriture, habitats,
abris), et (3) les ressources qui sont partagées doivent être limitées dans l’environnement (De Boer et Prins
1990). La compétition par interférence implique des interactions directes qui ont généralement lieu lors de
l’utilisation d’une ressource située au même endroit et disponible simultanément. Ainsi, l’accès à la ressource
commune peut devenir limité pour l’espèce présentant une moins grande capacité compétitive (Putman 1996).
Par exemple, Ferretti et al. (2010) ont utilisé le mécanisme d’interférence pour expliquer l’effet négatif du
chevreuil (Capreolous capreolus) sur le daim (Dama dama) en Italie. Dans toutes les interactions observées, le
chevreuil était dominant sur le daim et occupait les habitats favorables comparativement au daim qui réagissait
en augmentant son niveau de vigilance et en diminuant son utilisation de ces habitats.
En contrepartie, la compétition par exploitation est indirecte et a lieu par l’utilisation de ressources communes,
mais à différents moments. Ce type de compétition implique que l’utilisation d’une ressource par une espèce
réduit sa disponibilité ultérieure pour l’autre espèce en compétition (Pianka 1981). Le mécanisme de compétition
par exploitation nécessite que la ressource partagée soit limitée dans l’environnement (De Boer et Prins 1990).
Pour une espèce, un accès restreint à une ressource essentielle peut avoir de multiples répercussions,
notamment sur la condition corporelle des individus et sur différents processus démographiques populationnels
1(Gurnell et al. 2004). Par exemple, les travaux de Richard et al. (2010) ont révélé que les densités élevées de
cerfs élaphes (Cervus elaphus) avaient une influence négative sur la masse des faons de chevreuils par la
compétition par exploitation. Selon cette étude, la réduction de la masse des faons serait liée à une alimentation
de moins bonne qualité par les femelles chevreuils pendant la gestation. Cette diminution serait causée par la
baisse de la disponibilité des ressources pour les femelles chevreuils dans les habitats où l’on retrouve des
densités élevées de cerfs élaphes qui peuvent consommer les mêmes ressources. Dans des habitats où la
régénération de la végétation est lente, l’impact de densité élevées d’un compétiteur comme le cerf élaphe peut
réduire la qualité d’habitat du chevreuil pendant plusieurs saisons et ainsi avoir des effets sur plusieurs cohortes
de faons (Richard et al. 2010).
La compétition apparente est un autre type de compétition interspécifique indirecte dont le mécanisme agit par
le biais d’un consommateur commun (ex. prédateur ou parasite ; Hatcher et al. 2006). Ce type de compétition
est, entre autres, observé entre l’orignal (Alces alces) et le caribou forestier (Rangifer tarandus) par leurs
interactions avec le loup (Canis lupus). L’augmentation des populations d’orignaux génère une réponse
numérique de leur principal prédateur, le loup, ce qui affectera également les populations d’autres proies
potentielles présentes sur le territoire, dont le caribou (Wittmer et al. 2005).
L’étude de la compétition interspécifique en milieu naturel
Pour qu’il y ait compétition interspécifique, il doit y avoir un chevauchement des niches écologiques de deux
espèces (Pianka 1981, Alley 1982). Toutefois, les niches écologiques d’espèces similaires ont tendance à
diverger au cours de l’évolution afin de favoriser un partitionnement temporel ou spatial des ressources et de
diminuer le potentiel de compétition. Le processus de spécialisation écologique est donc, dans certains cas, le
résultat de la compétition passée entre deux espèces (Fischer et Gates 2005, Ale et al. 2011). Ainsi, des
espèces peuvent utiliser des habitats similaires sans être en compétition en n’utilisant qu’une partie de leur
niche écologique fondamentale (Schoener 1974). Ce partitionnement de la niche peut se manifester
temporellement (Kotler et al. 1993) ou spatialement (Fischer et Gates 2005). En contrepartie, des espèces
peuvent être compétitrices dans des situations où les ressources deviennent limitées ou lors d’introduction
d’espèces (Gurnell et al. 2004).
L’étude du potentiel de compétition entre deux espèces doit considérer les variations spatiales et temporelles
de plusieurs aspects de la niche écologique, entre autres, par rapport à la sélection d’habitat et au régime
alimentaire (Putman 1996, Jung et al. 2015). Cette temporalité peut être importante à l’échelle saisonnière,
principalement en raison de la variation de la quantité de ressources disponibles qui peut mener à la compétition
lorsqu’elles sont limitées (DuBowy 1988). L’aspect temporel de la compétition peut également se manifester à
l’échelle journalière. En effet, certaines espèces, dont l’impala (Aepyceros melampus) et le grand koudou
2(Tragelahus strepsiceros), peuvent changer le moment du jour pendant lequel elles utilisent les points d’eau en
saison sèche afin de réduire la compétition par interférence avec l’éléphant (Valeix et al. 2007). De plus,
l’hétérogénéité spatiale de la distribution des ressources peut également influencer le potentiel de compétition.
Des habitats de moins bonne qualité peuvent servir de refuges pour l’espèce ayant de moins grandes capacités
compétitives (Atkinson et Shorrocks 1984). Une étude de Davies et al. (2021) en Afrique du Sud a montré que
l’hétérogénéité spatiale du paysage était un élément clé de la coexistence entre le lycaon (Lycaon pictus) et le
lion (Panthera leo). Cette hétérogénéité permet au lycaon de sélectionner des habitats évités par le lion afin de
réduire le risque d’interactions négatives.
Compétition pour l’espace : utilisation et sélection d’habitat
La sélection d’habitat opère par le fait qu’un animal utilise de manière disproportionnée (c.-à-d., sélectionne ou
évite) une ressource de son habitat comparativement à sa disponibilité (Johnson 1980). La sélection d’habitat
est un processus hiérarchique qui lie les individus et les populations aux différentes conditions biotiques et
abiotiques de leur environnement. Ainsi, Johnson (1980) a défini plusieurs ordres de sélection : (1) l’aire de
répartition d’une espèce, (2) le domaine vital d’un individu, (3) les habitats utilisés à l’intérieur du domaine vital
et (4) le choix des items alimentaires dans les sites d’alimentation. Cette hiérarchisation permet de relier les
études sur l’habitat à celles sur l’alimentation d’une espèce, puisque des facteurs variés dictent les choix des
individus à différentes échelles de sélection. Par exemple, des études chez le caribou forestier et l’orignal ont
montré que l’évitement de la prédation était le facteur le plus important à large échelle, alors que la sélection
d’habitat offrant des ressources alimentaires abondantes était déterminante à une échelle plus fine (Rettie et
Messier 2000, Dussault et al. 2005). Toutefois, il est possible que la sélection d’habitat d’une espèce puisse être
principalement expliquée par le même facteur à plusieurs échelles de sélection. C’est notamment le cas du
bœuf musqué (Ovibos moschatus) pour qui, en hiver, la sélection d’habitat à large et fine échelles est influencée
positivement par un faible couvert de neige et une disponibilité élevée de graminoïdes (Schaefer et Messier
1995). Boyce et al. (2003) ont suggéré que cela s’expliquerait par l’environnement relativement homogène dans
lequel on retrouve l’espèce en hiver.
Plusieurs études ont caractérisé la sélection d’habitat pour évaluer le potentiel de compétition (Fischer et Gates
2005, Wereszczuk et Zalewski 2015, Jung et al. 2018) et mesurer la similarité de niches écologiques entre deux
espèces (Lowrey et al. 2018). Les fonctions de sélection de ressources (RSF) constituent l’une des principales
méthodes pour évaluer la sélection d’habitat (Manly et al. 2002). Une RSF est définie comme étant toute fonction
proportionnelle à la probabilité d’utilisation d’une unité de ressource (Boyce et McDonald 1999). En ce sens,
une unité de ressource peut être considérée comme un type d’habitat, une quantité de nourriture disponible ou
un ensemble de variables environnementales (Lele et al. 2013). Les fonctions de sélection de ressources sont
utilisées, entre autres, pour mesurer l’impact de perturbations anthropiques (Sawyer et al. 2009, Skarin et al.
32018), quantifier les pertes d’habitat (Boulanger et al. 2012, Plante et al. 2018) ou caractériser les relations
interspécifiques (Courbin et al. 2009). À cet effet, Fischer et Gates (2005) ont comparé la sélection d’habitat, en
hiver, du caribou forestier à celle du bison des bois (Bison bison athabascae) et ont déterminé que cette dernière
différait à plusieurs échelles spatiales, ce qui limitait le potentiel de compétition entre les deux espèces.
Compétition pour les ressources alimentaires
L’étude des régimes alimentaires est fréquemment utilisée pour évaluer le potentiel de compétition à fine échelle
(Putman 1996). Les premières études visant à caractériser le régime alimentaire des herbivores ont été réalisées
avec la technique de la microhistologie (Putman 1984) alors que de nouvelles méthodes, comme le codage à
barre d’ADN (angl. metabarcoding) (Valentini et al. 2009), sont de plus en plus utilisées (Pompanon et al. 2012).
Parmi les avantages que cette méthode offre par rapport aux analyses microhistologiques, on note l’amélioration
de la résolution taxonomique et de la rapidité des analyses (Newmaster et al. 2013, Nielsen et al. 2018). De
manière générale, les étapes pour caractériser le régime alimentaire avec le codage à barre d’ADN consistent
d’abord à extraire l’ADN végétal d’échantillons provenant des fèces ou du système digestif de l’herbivore. Par
la suite, des séquences ciblées sont amplifiées et assignées à un taxon selon plusieurs critères (Fazekas et al.
2012, Creer et al. 2016). L’utilisation du codage à barre d’ADN est toutefois limitée par l’accès à des données
de référence fiables, idéalement provenant du même site d’étude ou d’un biome similaire, et par le manque
d’études comparatives avec d’autres méthodes pour valider l’estimation quantitative du contenu des régimes
alimentaires (Nielsen et al. 2018).
En plus d’être utilisée pour caractériser et quantifier le régime alimentaire chez les herbivores (Valentini et al.
2009, Rayé et al. 2011, Clare 2014) cette méthode a notamment permis d’évaluer le chevauchement de niche
alimentaire entre des espèces potentiellement compétitrices (Brown et al. 2014, Soininen et al. 2015). Par
exemple, Christopherson et al. (2019) ont utilisé cette technique pour étudier une population menacée de
caribou montagnard et une population d’orignaux qui étaient possiblement en compétition pour des ressources
alimentaires communes. L’étude a permis de déceler des différences marquées entre les régimes alimentaires
des deux espèces et de confirmer un mécanisme de compétition apparente par le biais de prédateurs communs
plutôt que par compétition par interférence ou exploitation.
L’étude de la compétition interspécifique doit impérativement considérer la variation temporelle du régime
alimentaire et de la niche écologique. En effet, Jung et al. (2015) ont étudié une communauté de 7 espèces
d’herbivores au Yukon et déterminé que l’influence de la saison doit être considérée pour évaluer le potentiel
de compétition par exploitation. Par exemple, le chevauchement du régime alimentaire entre les caribous et les
bisons était plus élevé en été qu’en hiver, en raison de la consommation accrue de graminoïdes par les caribous
en été. En contrepartie, le chevauchement du régime alimentaire entre les bisons et les chevaux semi-sauvages
4(Equus ferus caballus) est demeuré élevé en été et en hiver, suggérant un potentiel de compétition plus élevé
entre ces deux herbivores. De plus, dans leurs revues respectives, Connell (1983) et Schoener (1983) ont
rapporté que dans des études sur la compétition interspécifique ayant été conduites sur plusieurs années, la
présence de variabilité temporelle dans la compétition était démontrée dans plus d’une étude sur deux. En
moyenne, parmi ces études, les espèces étaient en compétition une année sur trois, possiblement en raison de
la variation temporelle dans la disponibilité des ressources (Wiens 1997, dans Connell 1983). En ce sens, la
variabilité interannuelle de l’abondance de ressources communes à deux ou plusieurs espèces peut influencer
les interactions interspécifiques et entrainer de la compétition lorsque ces ressources deviennent limitantes.
Dans le même ordre d’idées, Kotler et al. (1993) ont étudié le rôle de la variabilité spatiale et temporelle dans la
coexistence de deux espèces de gerbilles (Gerbillus pyramidum et G. allenbyi) aux niches écologiques
similaires. Leurs résultats indiquent que le partitionnement temporel de l’utilisation des ressources alimentaires
assure la coexistence des deux espèces dans les mêmes habitats. En effet, chaque espèce possède une
période pendant laquelle elle est plus efficace que l’autre dans la quête alimentaire, en fonction de la densité de
nourriture disponible (Kotler et al. 1993). Dans certains systèmes, la temporalité de la compétition est liée aux
patrons de déplacement, comme les migrations saisonnières. Une espèce migratrice peut se retrouver,
temporairement et à forte densité, dans les mêmes habitats qu’une espèce résidente (Milner-Gulland et al.
2011). Par exemple, une étude réalisée au Mexique sur l’utilisation de l’espace et les ressources alimentaires
de la paruline à calotte rousse (Basileuterus rufifrons) a montré qu’en saison humide, les individus s’alimentent
d’insectes dans la canopée et dans la strate arbustive, en proportions relativement égales. Toutefois, en saison
sèche, des espèces migratrices occupent principalement la canopée et les parulines à calotte rousse changent
leur utilisation de l’habitat en s’alimentant principalement dans la strate arbustive (Jedlicka et al. 2006). De plus,
en Suisse, Anderwald et al. (2015) ont noté qu’une population migratrice de cerf élaphe entre en compétition, à
la fois pour l’espace et pour les ressources, avec deux espèces résidentes, le chamois (Rupicapra rupicapra) et
le bouquetin (Capra ibex), lorsqu’elle se déplace dans les mêmes habitats en été.
Contexte de l’étude : potentiel de compétition entre le caribou
migrateur et le bœuf musqué
À l’échelle circumpolaire, le déclin des troupeaux de caribous/rennes migrateurs est généralisé et partiellement
synchronisé (Vors et Boyce 2009). Ce déclin serait attribué à une multitude de facteurs, incluant l’épuisement
des ressources alimentaires (Couturier et al. 2009, Newton et al. 2014, Newton et al. 2015), la prédation (Ballard
et al. 1997), les perturbations anthropiques (Plante et al. 2017, Plante et al. 2018) et les changements
climatiques (Sharma et al. 2009, Taillon et al. 2016). Les changements climatiques peuvent engendrer une
asynchronie trophique (Post et Forchhammer 2008), une modification de l’habitat par le phénomène
d’arbustification (Myers-Smith et al. 2011, Fauchald et al. 2017) et une plus grande fréquence d’évènements
5climatiques extrêmes (Stocker et al. 2013). L’accumulation de ces impacts et les interactions entre ces facteurs
peuvent avoir des effets négatifs exacerbés pour le caribou. Par exemple, la présence d’infrastructures linéaires
peut diminuer les habitats de qualité pour le caribou et également faciliter l’accès des prédateurs comme le loup
à leurs proies (DeMars et Boutin 2018). Le caribou/renne migrateur a peu de compétiteurs naturels, toutefois,
dans certaines parties de son aire de répartition, il partage des habitats avec le bœuf musqué. Le potentiel de
compétition entre les deux espèces a fait l’objet de quelques études (Wilkinson et al. 1976, Ihl et Klein 2001)
tout en étant source de préoccupations locales, notamment au Nord-du-Québec (Guimond et al. 2010).
Historique de la problématique entourant la compétition entre le
caribou et le bœuf musqué
Les inquiétudes concernant le potentiel de compétition entre le caribou migrateur et le bœuf musqué sont
présentes dans plusieurs régions de l’Arctique, où certaines populations endémiques de bœuf musqué ont
atteint des densités élevées (p. ex. > 60 000 individus sur l’île de Banks, Jenkins et al. 2011, Cuyler et al. 2020)
et où des populations introduites sont considérées en croissance (p. ex. Péninsule du Taymir [Russie]
Sheremetev et al. 2014, Cuyler et al. 2020). Cette préoccupation a d’abord été rapportée dans les années 1970
par les chasseurs de la communauté de Sachs Harbor sur l’île de Banks aux Territoires du Nord-Ouest
(Canada). Ces derniers estimaient que la croissance et l’expansion de la population du bœuf musqué sur l’île
de Banks étaient néfastes au caribou de Peary (Rangifer tarandus pearyi; Vincent et Gunn 1981).
Les premières études sur les interactions entre le bœuf musqué et le caribou ont conclu que des différences
dans l’utilisation de l’habitat et dans la composition du régime alimentaire des deux espèces réduisaient le
potentiel de compétition (Parker et Ross 1976, Wilkinson et al. 1976, Shank et al. 1978). Toutefois, des analyses
subséquentes ont noté une similitude des régimes alimentaires, soulignant l’importance du saule (Salix spp.)
dans le régime alimentaire des deux espèces d’ongulés (Klein 1992, Larter et Nagy 1997, 2004). Cette similitude
pourrait, toutefois, être liée à la densité élevée de bœuf musqué sur l’île de Banks pendant les années 1970. À
densité élevée, le broutement par le bœuf entrainerait une baisse de la disponibilité de ses ressources préférées
(c.-à-d. Carex spp) causant une augmentation de la consommation de ressources alimentaires secondaires
comme le saule, particulièrement en hiver (Larter et al. 2002). Pour leur part, Ihl et Klein (2001) ont étudié la
sélection d’habitat du caribou et du bœuf musqué en Alaska et ont conclu que, malgré l’utilisation de sites
d’alimentation similaires, les deux espèces n’étaient pas en compétition puisqu’elles sélectionnaient, à fine
échelle, des ressources alimentaires différentes. Les conditions environnementales, comme la quantité de neige
au sol, pourraient également avoir un impact sur le chevauchement de niche écologique entre le bœuf musqué
et le caribou. En hiver, des accumulations importantes de neige pourraient mener à la compétition pour l’accès
à la nourriture de qualité dans les secteurs où la végétation demeure disponible, comme les milieux élevés
(Larter et Nagy 1997).
6Il est néanmoins possible que ces deux espèces soient en compétition apparente par le biais de leur guilde de
prédateurs communs comme le loup et le grizzly, respectivement présents dans l’ensemble et dans l’ouest de
l’Amérique du Nord (Heard 1992, Lent 1999, Mech 2010, Arthur et Del Vecchio 2017). Le mécanisme de
compétition apparente pourrait également agir via la présence et l’évitement de parasites communs (Hughes et
al. 2009, Kutz et al. 2013). Hughes et al. (2009) ont rapporté que sur l’île Victoria au Nunavut, l’infection du
caribou par des nématodes pouvait mener à une diminution de la masse corporelle chez des femelles de deux
ans et plus. De plus, les auteurs ont émis l’hypothèse qu’un changement de l’aire d’hivernage du troupeau de
caribous pourrait être lié à l’évitement d’habitats dans lesquels le risque d’être parasité pourrait être plus grand
à la suite de leur utilisation par les bœufs musqués. Une étude de Van der Wal et al. (2000) sur le renne de
Svalbard (Ranfiger tarandus platyrhynchus) a montré que les individus pourraient éviter les habitats dans
lesquels les densités de fèces sont élevées, pour diminuer le risque d’infection par des larves de nématodes.
Les deux troupeaux de caribous migrateurs (Rangifer tarandus caribou) qui occupent le Nord-du-Québec et le
Labrador, le troupeau Rivière-aux-Feuilles (TRAF) et le troupeau Rivière George (TRG), sont en déclin
démographique depuis respectivement 2 à 3 décennies. Ces deux troupeaux font l’objet d’un suivi à long terme
depuis plus de 40 ans qui a permis de monitorer plusieurs centaines de caribous par télémétrie, de mener à des
inventaires aériens et de récolter divers indicateurs biologiques. Ces efforts ont été maintenus par les
gouvernements du Québec (TRAF et TRG) et de Terre-Neuve-et-Labrador (TRG). Depuis 2008, le programme
de recherche Caribou Ungava vise à mieux comprendre l’ensemble des facteurs qui influencent la dynamique
de population de ces troupeaux. C’est dans cette perspective que l’on s’intéresse aux relations interspécifiques
entre le caribou migrateur et l’autre espèce de grand herbivore présente au Nunavik, le bœuf musqué. Cette
problématique est d’autant plus intéressante que le bœuf musqué n’est pas endémique au Nord-du-Québec
(voir la section Écologie du bœuf musqué au Nord-du-Québec). Cette espèce introduite a présenté une
expansion de sa répartition et une croissance démographique qui ont alimenté les questionnements par rapport
à son impact potentiel sur le caribou migrateur, notamment en ce qui a trait au chevauchement spatial et au
chevauchement de leurs régimes alimentaires (Guimond et al. 2010, Kutz et al. 2017). Les communautés inuites
pour qui le caribou migrateur est une composante centrale à leur culture, leur approvisionnement alimentaire et
leur économie (Hummel et Ray 2008), ont d’ailleurs exprimé plusieurs préoccupations concernant cette
potentielle compétition (Guimond et al. 2010, Kutz et al. 2017).
Écologie du caribou migrateur au Nord-du-Québec
Le caribou migrateur est un écotype présent sur l’ensemble du continent nord-américain (Festa-Bianchet et al.
2011). Il se démarque des écotypes de caribou forestier et montagnard par l’utilisation d’aires de mise bas et
d’estivage dans la toundra arctique et par l’agrégation des individus en groupes de plusieurs milliers pour les
migrations printanières et automnales (Festa-Bianchet et al. 2011). Les individus complètent leur cycle annuel
7Vous pouvez aussi lire

















































