Faire la lumière sur l'horloge circadienne de l'épinoche à trois épines - Mémoire Marie-Pier Brochu Maîtrise en biologie - avec mémoire
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Faire la lumière sur l’horloge circadienne de l’épinoche à trois épines Mémoire Marie-Pier Brochu Maîtrise en biologie - avec mémoire Maître ès sciences (M. Sc.) Québec, Canada © Marie-Pier Brochu, 2021
Faire la lumière sur l’horloge circadienne de l’épinoche à trois épines Mémoire Marie-Pier Brochu Sous la direction de : Nadia Aubin-Horth, directrice de recherche
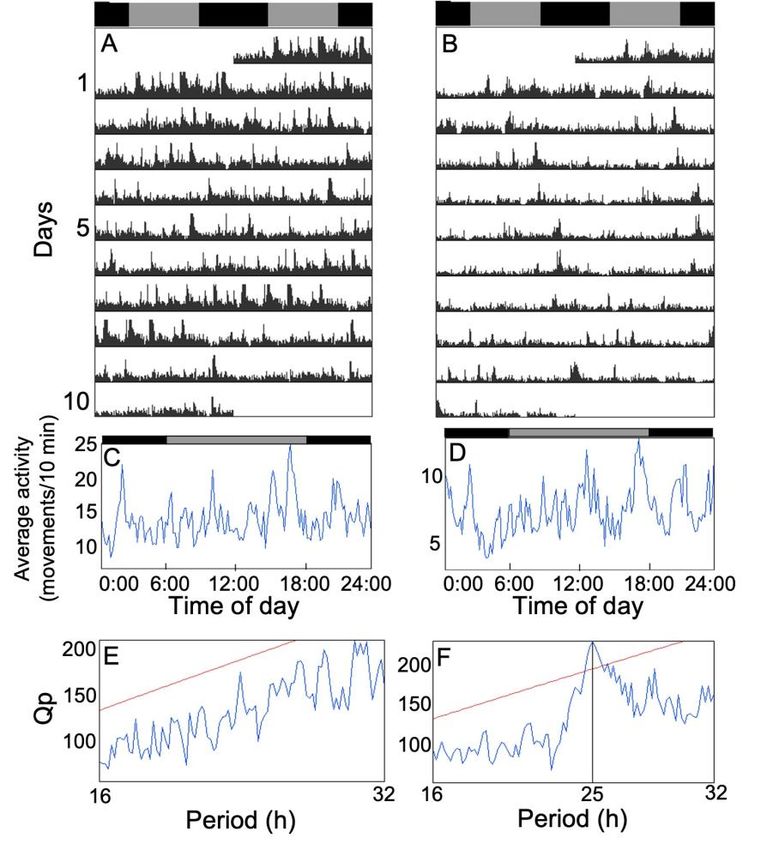
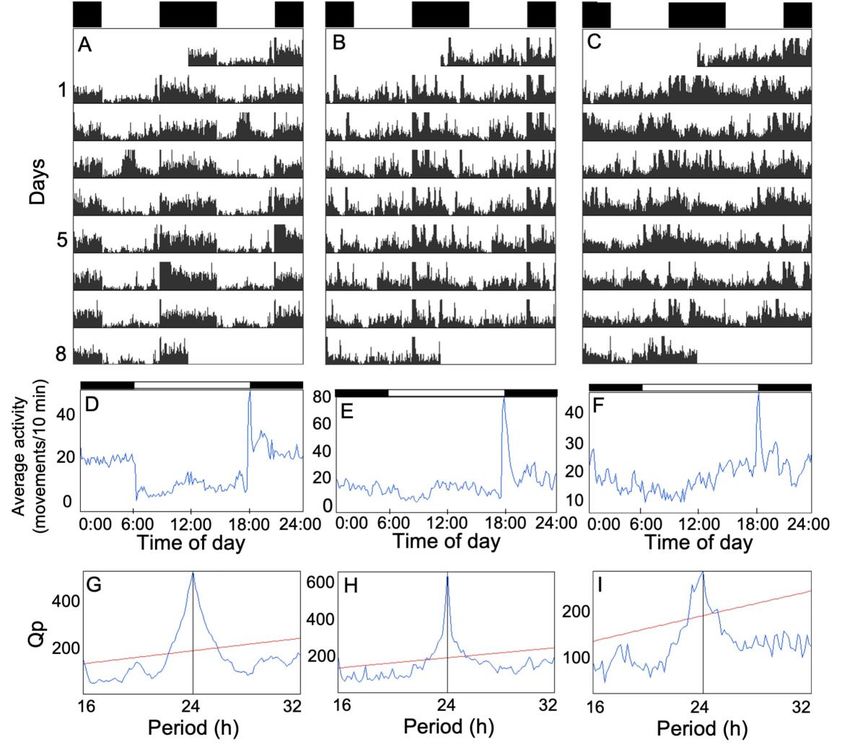
Résumé L’horloge circadienne est un système interne présent chez presque tous les organismes vivants et permettant de coordonner les fonctions biologiques entre elles et avec l’environnement. Dans les dernières décennies, l’horloge circadienne a majoritairement été étudiée en laboratoire à l’aide d’organismes modèles. Les connaissances concernant l’importance fonctionnelle et l’évolution des rythmes circadiens en nature sont donc limitées. L’épinoche à trois épines (Gasterosteus aculeatus) montre plusieurs caractéristiques intéressantes pour étudier l’horloge circadienne du point de vue de l’écologie et de l’évolution. Toutefois, ses rythmes circadiens n’ont presque jamais été étudiés et l’existence d’une horloge circadienne n’a jamais été démontrée chez cette espèce. À l’aide d’épinoches sauvages que nous avons rapportées au laboratoire, nous avons donc étudié l’horloge circadienne de cette espèce aux niveaux comportemental et moléculaire. Notre premier objectif était de déterminer si le rythme journalier de l’activité locomotrice est contrôlé par l’horloge circadienne. En quantifiant l’activité locomotrice sous un cycle lumière-noirceur (LD) et en noirceur constante (DD), nous avons montré que les épinoches ont bel et bien une horloge circadienne, mais que l’activité est faiblement contrôlée par celle-ci. Notre deuxième objectif était de déterminer la phase de l’activité des épinoches en LD. Nos résultats indiquent qu’elles sont majoritairement nocturnes, mais nous avons aussi observé une grande variation interindividuelle. Notre troisième objectif était de décrire l’oscillation moléculaire circadienne de cinq gènes de l’horloge (bmal1a, clock1b, clock2, per1b et cry1b) dans le cerveau des épinoches en DD en utilisant la PCR quantitative en temps réel. Nous n’avons pas détecté de rythmicité circadienne dans l’expression des gènes, ce qui pourrait soit indiquer que l’oscillateur moléculaire est très dépendant de la lumière ou qu’il y avait bel et bien une oscillation, mais que nous avons été incapables de la détecter. Dans l’ensemble, notre étude permet de mieux comprendre le rôle et les mécanismes de l’horloge circadienne chez l’épinoche et ouvre la voie à une exploration plus approfondie des rythmes circadiens chez cette espèce. ii
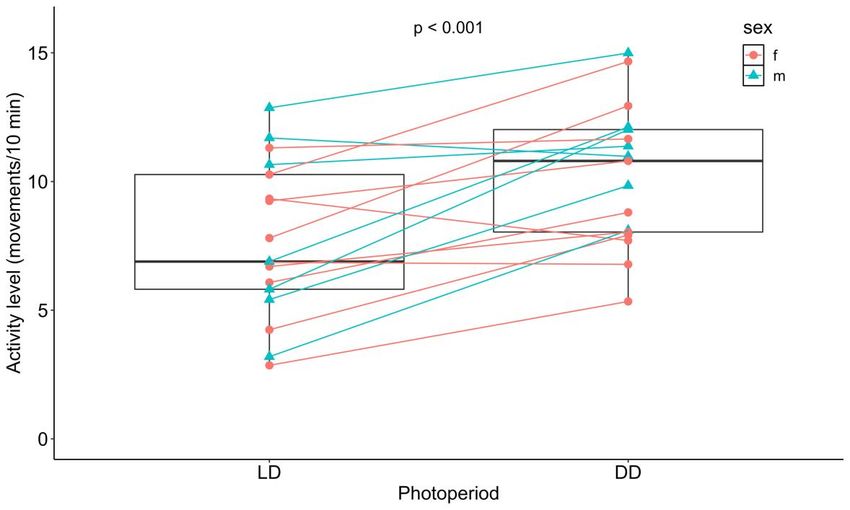
Abstract The circadian clock is an internal timekeeping system shared by almost all living organisms. In the last decades, the circadian clock has been mostly studied with model organisms in the laboratory, so knowledge about the functional importance and evolution of circadian rhythms in natural environments is limited. The threespine stickleback (Gasterosteus aculeatus) shows many interesting characteristics to study the circadian clock from an ecological and evolutionary perspective. However, its circadian rhythms have hardly ever been studied and the existence of a circadian clock has never been demonstrated in this species. In this study, using wild-caught threespine sticklebacks, we investigated the circadian clock of this species at the behavioral and molecular levels. Our first objective was to determine if the daily rhythm of locomotor activity is under circadian clock control. By quantifying locomotor activity in individual fish under a light-dark cycle (LD) and under constant darkness (DD), we showed that sticklebacks own an internal timekeeping system, but that locomotor activity is only weakly controlled by the clock. Our second objective was to determine the phase of activity of sticklebacks under LD. Our results indicate that they are mostly nocturnal, but we also observed large inter-individual variation. Our third objective was to describe the circadian molecular oscillation of five clock genes (bmal1a, clock1b, clock2, per1b and cry1b) in the brain of sticklebacks under DD using quantitative real-time PCR. We reported a lack of circadian rhythmicity for the five genes, which could either indicate that the clock molecular oscillator is highly light-dependent or that there was an oscillation but that we were unable to detect it. Overall, our study allows a better understanding of the role and mechanisms of the stickleback circadian clock and paves the way for further exploration of circadian rhythms in this species. iii
Table des matières Résumé .................................................................................................................................. ii Abstract ................................................................................................................................ iii Table des matières .............................................................................................................. iv Liste des tableaux ................................................................................................................ v Liste des figures .................................................................................................................. vi Liste des abréviations ........................................................................................................ vii Remerciements .................................................................................................................... ix Avant-propos ....................................................................................................................... xi Introduction ........................................................................................................................... 1 Les rythmes biologiques ................................................................................................. 1 L’horloge circadienne ..................................................................................................... 3 Les limites des organismes modèles ............................................................................. 9 L’épinoche à trois épines : une nouvelle espèce pour étudier l’horloge circadienne .. 10 Que savons-nous sur les rythmes journaliers de l’épinoche ? .................................... 11 Objectifs et hypothèses du projet ................................................................................. 13 Approches techniques .................................................................................................. 15 Chapitre 1: Shedding Light on the Threespine Stickleback Circadian Clock ............. 17 1.1 Résumé ...................................................................................................................... 18 1.2 Abstract....................................................................................................................... 19 1.3 Introduction ................................................................................................................. 20 1.4 Materials and methods ............................................................................................... 23 1.5 Results ........................................................................................................................ 28 1.6 Discussion .................................................................................................................. 30 1.7 Acknowledgments ...................................................................................................... 37 1.8 Tables ......................................................................................................................... 38 1.9 Figures ........................................................................................................................ 41 Conclusion .......................................................................................................................... 48 Limites de l’étude .......................................................................................................... 49 Les prochaines questions............................................................................................. 51 Bibliographie ....................................................................................................................... 53 Annexe 1 .............................................................................................................................. 64 iv
Liste des tableaux Table 1: The four core genes of the transcription-translation feedback loop (TTFL) of the clock molecular oscillator in mammals, the zebrafish and the stickleback and the references for the phylogenetic analyses. The five stickleback genes that we investigated in this study are in bold in the table ………………………………………………………………………...…. 38 Table 2: Characteristics of the primers used to quantify clock gene expression. Gene name, transcript sequence accession number from the Ensembl Genome Browser (Ensembl Transcript ID), forward and reverse primer sequences, qPCR amplicon size in base pairs (bp) and efficiency of the primer pair (%) ……………………………………………………… 39 Table 3: Characteristics of the primers used in the search for the optimal reference gene. Gene name, transcript sequence accession number from the Ensembl Genome Browser (Ensembl Transcript ID), forward and reverse primer sequences, qPCR amplicon size in base pairs (bp), efficiency of the primer pair (%) and reference if primer sequences were previously published. Using the NormFinder software, ubc was identified as the most stable gene between time points and was used as the reference gene (shown in bold) ………… 40 v
Liste des figures Figure I : La boucle rétroactive de transcription et de traduction de l’oscillateur moléculaire circadien dans une cellule ……………………………………………………………………..... 5 Figure 1: Experimental design …………………………………………………………………. 41 Figure 2: Under LD, sticklebacks display significant daily rhythmicity, but show variable activity patterns ………………………………………………………………………………….. 42 Figure 3: Under DD, most individuals are arrhythmic and only a few individuals show circadian rhythmicity …………………………………………………………………………….. 43 Figure 4: Under LD, sticklebacks are mostly nocturnal but show large inter-individual variation in the phase of activity ………………………………………………………….…….. 44 Figure 5: Under LD, sticklebacks show large inter-individual variation in the total daily activity …………………………………………………………………………………………….. 45 Figure 6: Under LD, the most nocturnal fish are also the least active ……………..……… 46 Figure 7: The expression of clock genes does not significantly vary during a 24-hour period in the brain of sticklebacks in DD ………………………………………………………………. 47 Figure S1: Position of the infrared photoelectric sensor on an experimental tank ……….. 64 Figure S2: Sensors position optimization …………………………………………….………. 65 Figure S3: Sticklebacks are less active during the light phase in LD than during the subjective light phase in DD ……………………………………….…………….……………... 66 vi
Liste des abréviations bp – base pairs cDNA – complementary deoxyribonucleic acid Cq – quantification cycle DD – dark-dark DNase – deoxyribonuclease h – hour ID – identification IDH – isocitrate dehydrogenase LED – light emitting diode LD – light-dark min – minute ng – nanogram NSC – noyau suprachiasmatique PCR – polymerase chain reaction qPCR – quantitative real-time PCR RIN – RNA integrity number RNA – ribonucleic acid s – second SCN – suprachiasmatic nucleus SEM – standard error of the mean TTFL – transcription-translation feedback loop µL, L – microliter, liter µM, nM – micromolar, nanomolar °C – degree Celsius vii
À ma mère Diane et à mon père Jules viii
Remerciements J’aimerais avant tout remercier ma directrice de recherche, Nadia Aubin-Horth. Nadia m’a fait entièrement confiance pour étudier un nouveau sujet dans son laboratoire : l’horloge circadienne. Elle m’a encouragée à sortir des sentiers battus et à poser de nouvelles questions sur un vieux modèle (l’épinoche à trois épines), ce qui a mené à plusieurs belles expériences et découvertes. Encore plus important, Nadia a marché à mes côtés tout au long de ce périple scientifique, que ce soit pour tenter (sans succès) de trouver la glande pinéale dans un minuscule cerveau d’épinoche, pour échantillonner des poissons sur le terrain ou pour me conseiller abondamment sur les manipulations de biologie moléculaire. Merci Nadia d’avoir été une directrice de recherche exceptionnelle. Ensuite, j’aimerais remercier Nicolas Cermakian qui a pris le temps de discuter avec moi de mon projet à plusieurs reprises. Ce fut très rassurant d’obtenir les conseils d’un expert dans le domaine de la chronobiologie. Un merci tout spécial à Dany Turcotte de la compagnie Westburne qui m’a grandement aidée à trouver le bon modèle de capteur pour mesurer l’activité de nos poissons et qui a bénévolement offert de son temps pour programmer le contrôleur. De plus, un grand merci à tous les membres du Laboratoire de Recherches en Sciences Environnementales et Médicales (LARSEM) de l’Université Laval qui veillent au bien-être des poissons et qui gèrent les installations aquatiques de notre centre de recherche. Je vous suis très reconnaissante pour toute l’aide technique que vous m’avez apportée et pour votre intérêt envers mon projet. Merci également à Julie Turgeon, Nicolas Cermakian et Nadia qui ont accepté d’évaluer ce mémoire. Vos commentaires et suggestions seront grandement appréciés. ix
J’aimerais aussi remercier les membres du laboratoire Aubin-Horth qui m’ont tous aidée à de nombreuses reprises durant ma maîtrise et que j’ai eu beaucoup de plaisir à côtoyer pendant quelques années. Merci à Florent Sylvestre, Verônica A. Alves, Chloé S. Berger et Sann Delaive pour leurs commentaires sur mes présentations orales et sur ce manuscrit. J’ai également une pensée toute spéciale pour Verônica A. Alves et Morgane Philippe qui m’ont aidée avec le travail de terrain et pour Sann Delaive qui a accepté de se lever en plein milieu de la nuit pour m’assister lors des dissections. Finalement, je n’aurais pas pu terminer cette maîtrise sans le soutien et l’amour inconditionnel de ma famille et de mes amis. Merci à ma mère Diane, à mon père Jules, à mon frère Raphaël, à mes merveilleuses amies Rébecca, Karine et Jennifer, à mes biologistes préférés Mireille et Louis-Philippe, à mes amis du Club l’Épaulé et à tous ceux que j’oublie. Une mention spéciale à ma maman qui est ma correctrice de toujours et qui, depuis le début de mes études universitaires, a lu tous mes travaux et demandes de bourse avec beaucoup d’attention. Merci aussi à mon amoureux Yohan qui m’a soutenue sans relâche dans mon aventure aux cycles supérieurs et qui a été à mes côtés dans les bons comme dans les moins bons moments. Merci à tous, je vous aime. x
Avant-propos Le chapitre principal de ce mémoire est présenté sous forme d’article scientifique et est intitulé « Shedding Light on the Threespine Stickleback Circadian Clock ». Cet article sera éventuellement soumis à la revue Journal of Experimental Biology. M.-P.B. a conçu le projet avec la contribution de N.A.-H. M.-P.B. a réalisé les expériences et analysé les données. M.-P.B. et N.A.-H. ont écrit le manuscrit. Auteurs : Marie-Pier Brochu et Nadia Aubin-Horth xi
Introduction Les rythmes biologiques Plusieurs comportements et processus physiologiques des organismes vivants sont rythmiques. Ces rythmes biologiques peuvent être induits par des cycles environnementaux qui sont eux-mêmes engendrés par la relation de la Terre avec le Soleil et la Lune (Refinetti 2008). Par exemple, la révolution annuelle de la Terre autour du Soleil et l’inclinaison de son axe de rotation causent les saisons et engendrent les rythmes circannuels, c’est-à-dire les rythmes qui ont une période approximative d’un an. Parmi ceux-ci, les rythmes migratoires, d’hibernation et de reproduction sont bien connus (Helm & Lincoln 2017). De plus, la rotation journalière de la Terre génère l’alternance entre le jour et la nuit, créant les rythmes circadiens (du latin circa diem, environ un jour). Le rythme de sommeil-éveil chez les humains et chez plusieurs animaux est certainement un des exemples les mieux établis de rythme circadien (Keene & Duboue 2018). De son côté, l’attraction gravitationnelle que la Lune exerce sur les masses d’eau produit les marées et engendre des rythmes circatidaux, c’est-à-dire des rythmes d’environ une demi-journée lunaire (12,4 h), dans le comportement de plusieurs espèces aquatiques occupant la zone intertidale (Tessmar- Raible et al. 2011). L’influence de la Lune sur la Terre provoque également, chez certaines espèces marines, des rythmes circalunaires et circasemilunaires correspondant à un cycle lunaire (29,5 jours) et à un demi-cycle lunaire (14,8 jours), respectivement. Ces deux types de rythmes sont surtout impliqués dans les événements de reproduction comme la ponte en masse des coraux (Tessmar-Raible et al. 2011). Tous ces cycles environnementaux créent une alternance prévisible entre des périodes favorables et moins favorables pour les organismes vivants. Les rythmes biologiques sont donc une réponse des organismes pour tirer profit – ou pour limiter les effets négatifs – de chaque période. Par exemple, dans les régions tempérées, plusieurs animaux profitent de l’été pour faire des réserves d’énergie et se reproduire alors que leurs activités sont beaucoup plus limitées pendant la saison froide (Foster & Kreitzman 2009). De façon similaire, les espèces diurnes recherchent de la nourriture et des partenaires sexuels le jour alors qu’elles se mettent à l’abri la nuit parce que leur système sensoriel est moins adapté pour l’exploitation de cette niche temporelle (Hut et al. 2012). En zone intertidale, certains 1
crustacés exploitent la marée basse pour trouver de la nourriture et s’enfouissent dans le sable à l’abri des prédateurs lorsque la marée monte à nouveau (de la Iglesia & Hsu 2010). La caractéristique commune des rythmes circannuels, circadiens, circatidaux et circalunaires est qu’ils ne constituent pas simplement une réponse directe à un environnement donné, mais bien qu’ils permettent aux organismes vivants d’anticiper les événements cycliques. En effet, l’arrivée d’une nouvelle saison, le début du jour et le changement de marée étant prévisibles, les individus peuvent se préparer à ces événements. Cette anticipation est essentielle, notamment parce que les changements comportementaux, physiologiques et même morphologiques qui surviennent au cours d’un cycle peuvent prendre plusieurs heures, jours ou mois à se mettre en place. C’est d’ailleurs ce qui rend les rythmes biologiques aussi intéressants : la notion d’anticipation implique que les organismes vivants possèdent un ou plusieurs mécanismes endogènes pour garder une trace du temps. L’existence de telles horloges internes n’est plus à prouver, bien que, pour certains types de rythmes, les mécanismes exacts de chronométrage ne soient pas encore entièrement compris. Généralement, démontrer l’origine endogène d’un rythme est assez simple. Il suffit de sélectionner des organismes qui sont rythmiques dans leur milieu naturel, de les exposer à un environnement constant en laboratoire, par exemple en conditions de noirceur et de température constante, et de vérifier s’ils conservent leur rythmicité. C’est de cette manière que des études pionnières en chronobiologie ont dévoilé l’origine endogène des rythmes circannuels (ex. : Pengelley & Fisher 1957, Gwinner & Dittami 1990), circadiens (ex. : Richter 1922, Aschoff & Wever 1962), circatidaux (ex. : Isaac & Jarvis 1973, Beentjes & Williams 1986) et circalunaires (ex. : Hauenschild 1960, Neumann 1966). Lorsqu’un comportement ou un processus physiologique conserve sa rythmicité en conditions constantes, la période du rythme (c.-à-d. la durée d’un cycle) est en libre cours et peut soit être un peu plus longue ou un peu plus courte que la période en conditions normales. La période libre cours correspond à la véritable période de l’horloge interne (Floessner & Hut 2017). C’est d’ailleurs pour cette raison que seuls les rythmes qui ont une origine endogène sont qualifiés de « circa » (« environ » en latin). Par exemple, un rythme journalier n’est pas nécessairement circadien, c’est-à-dire qu’il n’est pas nécessairement contrôlé par une horloge interne. En chronobiologie, lorsqu’un individu répond directement à son environnement sans qu’une horloge interne ne soit impliquée, on dit que le signal environnemental a un effet masquant sur l’organisme (Mrosovsky 1999). L’effet masquant 2
correspond donc à une simple synchronisation d’un comportement ou d’un processus physiologique par l’environnement. Lorsqu’un mécanisme interne est impliqué, on dit plutôt qu’il y a entraînement du rythme en question par le signal environnemental (Bittman 2020). L’horloge circadienne Parmi les rythmes biologiques, les rythmes circadiens sont sans contredit les plus étudiés. En raison de leur période de 24 h, ils sont plus pratiques à observer que les rythmes circannuels. Ils sont également beaucoup plus communs que les rythmes circatidaux ou circalunaires. En fait, les rythmes circadiens sont présents chez la plupart des organismes vivants. Des cyanobactéries aux animaux en passant par les plantes et les champignons, presque toutes les formes de vie peuvent présenter de la rythmicité circadienne (Bhadra et al. 2017). De plus, la perturbation des rythmes circadiens est reliée à plusieurs problèmes de santé chez les humains. Notamment, un décalage entre le rythme circadien interne et l’environnement, qui survient par exemple lors du travail de nuit, est associé à un risque plus élevé de développer un cancer (Pariollaud & Lamia 2020). Le système endogène responsable des rythmes circadiens est aujourd’hui bien connu : il s’agit de l’horloge circadienne. Le cœur de l’horloge circadienne est constitué d’un oscillateur moléculaire qui a une période intrinsèque d’environ 24 h et qui est autonome au niveau de la cellule. Depuis que le premier gène de l’horloge a été identifié par clonage moléculaire chez la drosophile (Drosophila melanogaster) en 1984 (Bargiello et al. 1984, Bargiello & Young 1984, Reddy et al. 1984, Zehring et al. 1984), notre compréhension des mécanismes moléculaires circadiens n’a pas cessé de s’améliorer. Nous savons maintenant que l’oscillateur moléculaire est composé d’une boucle rétroactive de transcription et de traduction (transcription-translation feedback loop, TTFL) comprenant des éléments négatifs et positifs (Figure I). Bien que ces éléments peuvent varier entre les règnes du vivant, le fonctionnement de la TTFL est universel, suggérant une évolution convergente des mécanismes moléculaires de l’horloge circadienne (Bell-Pedersen et al. 2005). Chez les mammifères, les protéines BMAL1 (aussi connue sous le nom d’ARNTL) et CLOCK (et son paralogue NPAS2) sont des éléments positifs. Il est intéressant de noter que BMAL1 a aussi un paralogue nommé BMAL2 (ou ARNTL2), mais, contrairement à BMAL1, la protéine BMAL2 n’est pas suffisante pour maintenir l’oscillation moléculaire au niveau cellulaire (Xu 3
et al. 2015). Les protéines BMAL1 et CLOCK forment un hétérodimère et activent la transcription des gènes period (per1, per2, per3) et cryptochrome (cry1, cry2) qui codent pour les protéines PER et CRY, des éléments négatifs. Lorsque les protéines PER et CRY sont présentes en quantité suffisante dans le cytoplasme, elles forment un complexe avec d’autres molécules et retournent dans le noyau pour interagir avec BMAL1-CLOCK et réprimer leur propre transcription. Pendant que la transcription diminue, les niveaux de PER et CRY déclinent. Lorsque les éléments négatifs ne sont plus présents en quantité suffisante, l’activité de BMAL1-CLOCK peut reprendre et le cycle se répète (Figure I). Évidemment, il s’agit d’une description très simplifiée de l’oscillateur moléculaire et plusieurs articles faisant une revue détaillée des connaissances sont disponibles (ex. : Takahashi 2017, Rosensweig & Green 2020). De plus, malgré qu’il puisse y avoir des différences fonctionnelles, les gènes impliqués dans l’oscillateur moléculaire mentionnés ci-dessus, aussi appelés les gènes de l’horloge, sont conservés chez les animaux comme chez les insectes (Tataroglu & Emery 2015), les poissons (Vatine et al. 2011) et les oiseaux (Cassone 2014). Lorsqu’on désire étudier l’oscillateur moléculaire d’une cellule, d’un tissu ou d’un organe, c’est généralement le rythme circadien d’expression des gènes de l’horloge qui est utilisé, c’est-à-dire qu’on mesure le niveau d’expression des gènes à différents moments de la journée pendant au minimum un cycle de 24 h (ex. : Mure et al. 2018). On s’intéresse surtout à l’amplitude (différence entre le niveau moyen d’expression et la valeur maximale ou minimale de l’expression) et à l’acrophase (moment auquel l’expression est maximale) du rythme (Refinetti et al. 2007). En règle générale, on s’attend à ce que le niveau d’expression des gènes qui codent pour les éléments positifs de la TTFL soit en antiphase avec le niveau d’expression des gènes qui codent pour les éléments négatifs. En d’autres mots, lorsque l’expression de bmal1 et clock est à son plus haut, l’expression de per et cry est à son plus bas et vice-versa (Takahashi 2017). Cela dit, il y des exceptions. Par exemple, l’expression de clock est constante dans certains tissus chez les mammifères (Oishi et al. 1998, Shearman et al. 2000). 4
Figure I : La boucle rétroactive de transcription et de traduction de l’oscillateur moléculaire circadien dans une cellule. Chez les mammifères, les protéines BMAL1 et CLOCK sont des éléments positifs (en jaune sur la figure) qui forment un hétérodimère et activent la transcription des gènes period (per) et cryptochrome (cry). Les protéines PER et CRY sont des éléments négatifs (en bleu sur la figure) qui, lorsqu’ils sont présents en quantité suffisante dans le cytoplasme, retournent dans le noyau pour interagir avec BMAL1-CLOCK et réprimer leur propre transcription. Pendant que la transcription diminue, les niveaux de PER et CRY déclinent. Lorsque les éléments négatifs ne sont plus présents en quantité suffisante, l’activité de BMAL1-CLOCK peut reprendre et le cycle se répète. Cet oscillateur moléculaire a une période intrinsèque d’environ 24 h et est autonome au niveau de la cellule (Takahashi 2017, Rosensweig & Green 2020). Chez les animaux, l’oscillateur moléculaire est retrouvé dans la plupart des tissus et des organes (insecte : Plautz et al. 1997, poisson : Whitmore et al. 1998, oiseau : Chong et al. 2003, mammifère : Yoo et al. 2004). Tous ces oscillateurs autonomes doivent donc être coordonnés pour que l’organisme fonctionne de manière optimale. De plus, bien qu’il ait une période intrinsèque d’environ 24 h, l’oscillateur moléculaire est entraîné par l’environnement pour que sa phase et sa période soient en harmonie avec, par exemple, le cycle lumière- 5
noirceur. Chez les mammifères, la coordination des oscillateurs entre eux et l’entraînement par l’environnement se font par l’intermédiaire d’un oscillateur maître situé dans l’hypothalamus, le noyau suprachiasmatique (NSC) (revue dans Reppert & Weaver 2002, Welsh et al. 2010). Sommairement, les cellules rétiniennes captent les signaux lumineux et envoient des informations au NSC par le tractus rétinohypothalamique. Les cellules neuronales du NSC, qui contiennent chacune un oscillateur moléculaire autonome, interagissent et sont entraînées par l’environnement lumineux. Par l’intermédiaire de stimulus nerveux et humoraux, le NSC envoie ensuite des signaux aux autres tissus et aux organes pour coordonner les oscillateurs périphériques et pour que tout l’organisme soit en harmonie avec le cycle lumière-noirceur. Par exemple, le NSC induit la sécrétion nocturne de la mélatonine dans la glande pinéale via la noradrénaline qui est libérée par des fibres nerveuses sympathiques. La mélatonine est importante pour entraîner les rythmes circadiens de nombreux oscillateurs périphériques puisque ses récepteurs se retrouvent dans plusieurs tissus. Les récepteurs de la mélatonine se retrouvent aussi dans le NSC; il peut donc y avoir une rétroaction de la mélatonine en circulation sur l’oscillateur maître (Pevet & Challet 2011). Ainsi, par l’intermédiaire de la mélatonine et de plusieurs autres signaux, le NSC entraîne des rythmes circadiens dans les comportements et les processus physiologiques comme l’activité locomotrice, la pression sanguine, la température corporelle, la sécrétion d’hormone, le métabolisme, etc. On dit d’ailleurs que le NSC est l’oscillateur maître de l’horloge circadienne des mammifères puisqu’il est nécessaire et suffisant pour la génération de plusieurs rythmes circadiens. Par exemple, l’ablation du NSC élimine les comportements circadiens (Stephan & Zucker 1972, Rusak 1977) alors que sa transplantation les restaure (Lehman et al. 1987, Ralph et al. 1990). Les signaux environnementaux pouvant entraîner l’horloge circadienne et les rythmes circadiens sont appelés des zeitgebers. Le cycle lumière-noirceur est un des zeitgebers principaux de l’horloge circadienne. Comme expliqué précédemment, l’entraînement des rythmes circadiens à ce signal environnemental se fait par l’intermédiaire du NSC chez les mammifères. Cela dit, plusieurs autres zeitgebers peuvent entraîner l’horloge circadienne indépendamment du NSC. En effet, comme l’oscillateur moléculaire se retrouve dans la plupart des tissus et des organes, ceux-ci peuvent être entraînés par des signaux non lumineux. Par exemple, la disponibilité de la nourriture et les glucocorticoïdes peuvent entraîner les rythmes circadiens dans des organes comme le cœur et le foie (Balsalobre et 6
al. 2000, Damiola et al. 2000, Stokkan et al. 2001). Les oscillateurs périphériques reçoivent donc des informations temporelles locales en plus des signaux provenant de l’oscillateur maître. Il semble également que les oscillateurs périphériques envoient à leur tour des signaux au NSC, mais les mécanismes permettant cette communication bidirectionnelle sont encore mal connus (revue dans Richards & Gumz 2012). Chez les autres animaux, l’organisation hiérarchique de l’horloge circadienne montre certaines différences importantes par rapport aux mammifères. Chez les oiseaux, il n’y a pas que le NSC qui joue le rôle d’oscillateur maître : la glande pinéale est également un oscillateur central du système circadien. Comme chez les mammifères, la glande pinéale reçoit des signaux nerveux du NSC, mais, en plus, cette structure est photoréceptrice chez les oiseaux et elle contient un oscillateur moléculaire qui peut être entraîné par la lumière (Natesan et al. 2002). L’oscillateur moléculaire des cellules de la glande pinéale contrôle l’expression rythmique du gène qui code pour l’enzyme responsable de la synthèse de la mélatonine (Bernard et al. 1997a, Bernard et al. 1997b). Ainsi, chez les oiseaux, le rythme de sécrétion de la mélatonine dans la glande pinéale est contrôlé non seulement par le NSC (comme chez les mammifères), mais aussi localement par l’environnement lumineux. De cette façon, les rythmes circadiens des oiseaux peuvent être contrôlés par la glande pinéale indépendamment du NSC. Cela dit, les interactions entre la glande pinéale et le NSC semblent essentielles pour la stabilisation et le maintien de l’oscillation circadienne de chacune de ces structures (revue dans Cassone 2014). Chez les poissons, un groupe pour lequel les connaissances proviennent en majorité du poisson-zèbre (Danio rerio), l’organisation de l’horloge circadienne est encore plus décentralisée que chez les oiseaux. Bien que les poissons possèdent un NSC, il ne semble pas avoir un rôle d’oscillateur maître, notamment parce qu’il n’est pas nécessaire pour instaurer l’oscillation moléculaire circadienne chez les embryons de poissons-zèbres (Noche et al. 2011). Pour ce qui est de la glande pinéale, elle est similaire chez les poissons et les oiseaux. En effet, la glande pinéale des poissons est photoréceptrice, elle contient un oscillateur moléculaire indépendant et elle sécrète de la mélatonine (Ekström & Meissl 1997). Cependant, bloquer génétiquement la glande pinéale diminue l’amplitude du rythme circadien de l’activité locomotrice chez les poissons-zèbres, mais cela n’affecte pas la 7
période du rythme d’activité ni le rythme circadien des oscillateurs moléculaires périphériques (Livne et al. 2016). De plus, une mutation qui élimine la production de mélatonine chez les poissons-zèbres empêche la régulation circadienne du sommeil, mais cela ne perturbe pas les autres rythmes circadiens (Gandhi et al. 2015). Ces résultats suggèrent donc que la glande pinéale n’est pas le seul oscillateur impliqué dans l’entraînement des rythmes circadiens chez les poissons. D’ailleurs, comme chez les autres animaux, la plupart des cellules des poissons contiennent un oscillateur moléculaire autonome (Whitmore et al. 1998). La différence avec les mammifères et les oiseaux est que tous les tissus et les organes des poissons étudiés jusqu’à maintenant sont sensibles à la lumière (Whitmore et al. 2000). Ainsi, lorsqu’on place, par exemple, des cellules de cœur en culture et qu’on les expose à un cycle lumière-noirceur, l’oscillateur moléculaire est entraîné (Whitmore et al. 2000). On ne sait pas encore exactement comment les cellules des poissons captent le signal lumineux, mais trois photorécepteurs principaux ont été proposés jusqu’à maintenant : les opsines non visuelles (Davies et al. 2015), les cryptochromes (Cermakian et al. 2002) et les oxydases contenant de la flavine (Hirayama et al. 2007). Le signal lumineux capté par la cellule induit la transcription de gènes de l’horloge sensibles à la lumière (per2 et cry1a chez le poisson-zèbre) et ainsi entraîne l’oscillateur moléculaire (Ziv et al. 2005, Tamai et al. 2007). De cette façon, chaque tissu et chaque organe peut, en théorie, agir comme oscillateur maître et entraîner de façon indépendante les rythmes circadiens. En réalité, il est plus probable que les rythmes circadiens émergent des interactions entre plusieurs oscillateurs distribués dans différents tissus de l’organisme (revue dans Frøland Steindal & Whitmore 2019). Pour ce qui est des insectes, les connaissances proviennent en grande partie des études sur la drosophile (Drosophila melanogaster). Comme chez les poissons, des oscillateurs moléculaires sont présents dans presque tous les tissus de la drosophile et peuvent être directement entraînés par la lumière (Plautz et al. 1997). Cela dit, un ensemble de neurones dans le cerveau de la drosophile possède une fonction similaire à celle du NSC des mammifères. En effet, ces neurones sont nécessaires et suffisants pour entraîner le rythme circadien de l’activité locomotrice (Renn et al. 1999, Blanchardon et al. 2001, Grima et al. 2004) et sont impliqués à différents niveaux dans le contrôle des rythmes circadiens physiologiques (ex. : Erion et al. 2016). Certains oscillateurs périphériques semblent fonctionner de manière indépendante (ex. : Tanoue et al. 2004), mais en général, ils ont un 8
certain niveau de dépendance aux oscillateurs dans les neurones du cerveau (revue dans Dubowy & Sehgal 2017). Les limites des organismes modèles Notre connaissance des mécanismes moléculaires et de l’organisation hiérarchique de l’horloge circadienne a été acquise majoritairement en laboratoire avec l’aide d’organismes modèles comme la souris (Mus musculus) et d’autres espèces de rongeurs, le poisson- zèbre (Danio rerio) et la drosophile (Drosophila melanogaster). À une époque où les outils moléculaires étaient moins accessibles, ces espèces ont été essentielles entre autres pour étudier la fonction des gènes de l’horloge par l’intermédiaire de manipulations génétiques. Cependant, les organismes modèles présentent certaines limites, notamment pour répondre à des questions en écologie et en évolution (Kronfeld-Schor et al. 2013, Schwartz et al. 2017). Premièrement, ils représentent une très faible diversité d’organismes vivants qui ont été choisis entre autres parce qu’ils présentaient des rythmes circadiens robustes. Cela est problématique parce qu’il y a de plus en plus d’évidences suggérant que le rôle de l’horloge circadienne (c.-à-d. son niveau d’implication dans le contrôle des rythmes biologiques) peut grandement varier dépendamment de la biologie de l’espèce (revue dans Bloch et al. 2013, Hazlerigg & Tyler 2019). Par exemple, chez certains herbivores qui doivent s’alimenter à de nombreuses reprises pendant la journée, les activités quotidiennes ne sont pas entraînées par l’horloge circadienne, mais présentent plutôt un rythme ultradien, c’est-à-dire un rythme avec une période plus courte que 24 h (Daan & Slopsema 1978, Liu et al. 2007). Chez les oiseaux, les périodes de migration sont parfois associées à une diminution drastique du sommeil et à une perte de rythmicité circadienne pendant plusieurs jours (Rattenborg et al. 2004, Yadav et al. 2021). Chez les insectes eusociaux, les différentes castes présentent souvent des rythmes circadiens distincts : les individus qui prennent soin des larves sont actifs jour et nuit alors que ceux qui recherchent de la nourriture à l’extérieur de la colonie montrent des rythmes circadiens de l’activité très robustes (Moore et al. 1998, Mildner & Roces 2017). Deuxièmement, une autre limite des organismes modèles provient du fait qu’ils sont issus de lignées de laboratoire qui ont été élevées dans des conditions optimales et contrôlées depuis de nombreuses générations. Cela n’est pas représentatif du milieu naturel dans lequel les organismes doivent faire face à plusieurs défis, menaces et opportunités et ajuster leur patron d’activités en conséquence. Par exemple, certaines espèces changent la phase de leur activité journalière (c.-à-d. le 9
moment de la journée pendant lequel elles sont actives) en réponse à la présence de parasites (Zukl et al. 1993), à la compétition interspécifique (Gutman & Dayan 2005), aux contraintes thermo-énergétiques (van der Vinne et al. 2014), à la disponibilité de la nourriture (Ware et al. 2012), aux opportunités d’accouplement (Lesku et al. 2012), au risque de prédation (Pellman et al. 2015), etc. Ce changement de phase peut être occasionné par un effet masquant ou par l’entraînement de l’horloge circadienne. Tous ces exemples soulignent la nécessité de s’intéresser à une grande diversité d’espèces sauvages qui ont évolué dans des contextes écologiques variés pour mieux comprendre l’importance fonctionnelle de l’horloge circadienne dans le milieu naturel (c.-à-d. les avantages qu’elle confère à une espèce dans un environnement donné) et, par le fait même, quelles pressions de sélection peuvent façonner l’évolution des rythmes circadiens. L’épinoche à trois épines : une nouvelle espèce pour étudier l’horloge circadienne L’épinoche à trois épines (Gasterosteus aculeatus) présente des caractéristiques écologiques et évolutives très intéressantes. À l’origine, ce petit poisson était confiné au milieu marin, mais lors de la fin de la dernière époque glaciaire, l’épinoche a colonisé à répétitions de nombreux habitats d’eau douce, menant à une radiation adaptative massive de l’espèce (Bell & Foster 1994, McKinnon & Rundle 2002). Aujourd’hui, on retrouve des épinoches en abondance partout à travers l’hémisphère Nord autant en eau douce (lacs, ruisseaux, marais, rivières) qu’en eau salée en plus des populations anadromes (Fang et al. 2018). Depuis des dizaines d’années, des chercheurs s’intéressent aux traits qui ont permis aux épinoches de s’adapter à autant d’habitats variés. Étant donné que la forme ancestrale marine existe toujours, il est possible d’évaluer quels traits ont évolué récemment et s’ils sont des adaptations à un milieu particulier. De cette façon, les traits morphologiques, physiologiques, d’histoire de vie et comportementaux sont largement décrits chez les différents écotypes d’épinoches (Ostlund-Nilsson et al. 2007). Les bases génétiques de ces traits sont également de mieux en mieux comprises (ex. : Cresko et al. 2004, Shapiro et al. 2004, Kitano et al. 2010). Or, malgré toute les connaissances que nous avons sur la biologie de l’épinoche (Bell & Foster 1994, Ostlund-Nilsson et al. 2007), très peu d’informations sont disponibles sur ses rythmes circadiens. En fait, l’existence d’une horloge circadienne n’a jamais été démontrée chez cette espèce. 10
L’épinoche est une espèce prometteuse pour répondre à des questions d’ordre écologique et évolutive sur l’horloge circadienne. Comme les écotypes d’épinoches font face à des environnements très différents (variations dans la présence de prédateurs et de parasites, le type de nourriture, l’intensité et la composition spectrale de la lumière, etc.), il serait intéressant d’éventuellement pouvoir comparer leurs rythmes circadiens. On pourrait évaluer, par exemple, si certains comportements ou processus physiologiques sont plus ou moins contrôlés par l’horloge circadienne entre les écotypes, s’il y a des différences de phase dans les rythmes circadiens et si ces variations ont une valeur adaptative dans un contexte écologique spécifique. D’ailleurs, dans une expérience de jardin commun, il a déjà été rapporté que le gène de l’horloge cry1ab était surexprimé dans le foie d’épinoches provenant d’un ruisseau par rapport aux épinoches provenant d’un lac à proximité (Hanson et al. 2017). La fonction biologique de cette différence n’est pas connue et il faudrait probablement s’intéresser au rythme circadien d’expression du gène chez les deux populations pour en apprendre davantage. Néanmoins, ce résultat suggère que les mécanismes moléculaires circadiens peuvent varier entre les écotypes. De plus, les épinoches sont faciles à échantillonner en nature et sont assez petites pour être rapportées en grand nombre au laboratoire et maintenues en captivité lorsque cela est nécessaire, ce qui est parfois le cas pour déterminer, par exemple, si un rythme biologique est contrôlé par l’horloge circadienne. Le génome des épinoches est également séquencé et annoté (Jones et al. 2012, Peichel et al. 2017), ce qui est très utile pour étudier l’oscillateur moléculaire circadien. Que savons-nous sur les rythmes journaliers de l’épinoche ? Au niveau comportemental, nous avons certaines informations sur le rythme journalier d’activité de l’épinoche dans différents écotypes, mais on ne sait pas si ce comportement est sous contrôle circadien. En effet, à notre connaissance, les épinoches ont été étudiées une seule fois en conditions constantes. En utilisant une population anadrome, les auteurs de cette étude ont montré que la fréquence à laquelle les mâles visitaient leur nid (dans l’espoir d’y trouver des œufs déposés par une femelle) ne présentait pas de rythme circadien en lumière constante (Sevenster et al. 1995). Les autres informations que nous avons sur le rythme journalier d’activité des épinoches ne permettent pas de dire si cette espèce est diurne ou nocturne ni de mettre en évidence des tendances selon les écotypes. D’un côté, certaines évidences suggèrent que les épinoches sont diurnes. Notamment, les opsines 11
visuelles des épinoches (Rennison et al. 2012) correspondent à celles des poissons diurnes (Carleton et al. 2020). Des études ont également indiqué que les épinoches étaient majoritairement actives le jour dans un marais salant (Worgan & FitzGerald 1981), une rivière (Sjoberg 1985) et un ruisseau (Reebs et al. 1995), mais ces résultats ont été obtenus à l’aide de données de capture, c’est-à-dire en dénombrant à intervalles réguliers les poissons capturés dans des burroles pendant quelques jours. Cette méthode peut introduire plusieurs biais dans les résultats parce que tous les individus n’ont pas nécessairement la même probabilité d’entrer dans les pièges et parce que les burroles sont statiques, contrairement aux poissons qui peuvent se déplacer pendant la journée. D’un autre côté, des auteurs ont observé que certaines épinoches marines s’alimentaient la nuit en laboratoire (Mussen & Peeke 2001) et que l’activité de ventilation des mâles envers les œufs dans leur nid était légèrement plus élevée la nuit que le jour dans un marais salant (Reebs et al. 1984). De plus, les épinoches semblent présenter un rythme journalier de la migration verticale dans la colonne d’eau d’un lac : elles restent en profondeur durant le jour et sont actives à la surface durant la nuit (Quinn et al. 2012). D’un point de vue physiologique, nous savons que le niveau de mélatonine est plus élevé la nuit que le jour dans le sang, le cerveau, les yeux, la peau et les reins des épinoches (Mayer et al. 1997, Kulczykowska et al. 2017, Pomianowski et al. 2020). Cette même variation journalière est observée chez tous les autres vertébrés, qu’ils soient diurnes ou nocturnes (Challet 2007, Falcón et al. 2009). Ce résultat montre donc que les épinoches sont similaires aux autres vertébrés concernant le rythme de sécrétion de la mélatonine, mais ne nous donne pas d’information sur le contrôle potentiel de l’horloge circadienne. En effet, l’enzyme qui s’occupe de la synthèse de la mélatonine peut être contrôlée directement par la lumière sans l’intervention de l’oscillateur moléculaire (Falcón et al. 2009). Ainsi, comme le niveau de mélatonine a été mesuré alors que les épinoches étaient exposées à un cycle lumière-noirceur, on ne peut pas savoir s’il y a un contrôle endogène de cette hormone. Au niveau moléculaire, il a été démontré que l’expression des gènes per1b et clock1b variait significativement au cours d’une journée dans le foie d’épinoches exposées à un cycle lumière-noirceur en laboratoire (Prokkola et al. 2015). Ce résultat suggère qu’il y a un 12
Vous pouvez aussi lire

















































