Lutte biologique contre le tarsonème du fraisier (Acari : Tarsonemidae) à l'aide d'acariens prédateurs (Acari : Phytoseiidae) en fraisière - Corpus UL
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Lutte biologique contre le tarsonème du fraisier
(Acari : Tarsonemidae) à l’aide d’acariens
prédateurs (Acari : Phytoseiidae) en fraisière
Mémoire
Stéphanie Patenaude
Maîtrise en biologie végétale
Maître ès sciences (M. Sc.)
Québec, Canada
© Stéphanie Patenaude, 2018
Lutte biologique contre le tarsonème du fraisier
(Acari : Tarsonemidae) à l’aide d’acariens
prédateurs (Acari : Phytoseiidae) en fraisière
Mémoire
Stéphanie Patenaude
Sous la direction de :
Valérie Fournier, directrice de recherche
Stéphanie Tellier, codirectrice de recherche
Résumé
Le tarsonème du fraisier Phytonemus pallidus Banks (Acari : Tarsonemidae) est un acarien ravageur
invisible à l’œil nu et de plus en plus préoccupant en fraisière en raison des pertes de rendement
considérables qu’il occasionne. Le retrait récent de l’insecticide/acaricide endosulfan nécessite la
recherche de solutions de remplacement. La lutte biologique à l’aide d’acariens prédateurs (Acari :
Phytoseiidae) fait partie de celles-ci, mais aucune recherche n’avait encore été réalisée en champ sous
les conditions du Québec afin de valider sa faisabilité. Les objectifs de cette étude étaient : (1) évaluer
l’efficacité de trois espèces d’acariens phytoséiides, Neoseiulus cucumeris Oudemans, N. fallacis
Garman et Amblyseius andersoni Chant, à contrôler le tarsonème du fraisier en conditions de
fraisières commerciales; (2) évaluer le rendement et la qualité des fruits en lien avec cette méthode
de lutte; (3) examiner l’effet possible de ces prédateurs sur d’autres ravageurs en fraisière et (4) mieux
comprendre la dynamique de population du tarsonème du fraisier sous notre climat. Nos résultats
démontrent que N. cucumeris offre une bonne efficacité, mais qu’il est très sensible au froid et trop
dispendieux pour une utilisation en champ. En 2017, lorsqu’un plafond de coût fut fixé (1000$/ha),
aucun des trois prédateurs n’a pu réduire de façon significative le nombre de tarsonèmes. Le suivi des
populations de ce ravageur au champ a permis la confirmation de certains faits connus, tout en
permettant l’acquisition de connaissances nouvelles et essentielles sur sa phénologie. À la lumière
des résultats obtenus, les trois phytoséiides ont offert un contrôle insuffisant du tarsonème du fraisier
en champ dans les conditions expérimentées. Le tarsonème du fraisier demeure un problème très
complexe et beaucoup de recherches supplémentaires sont nécessaires.
iii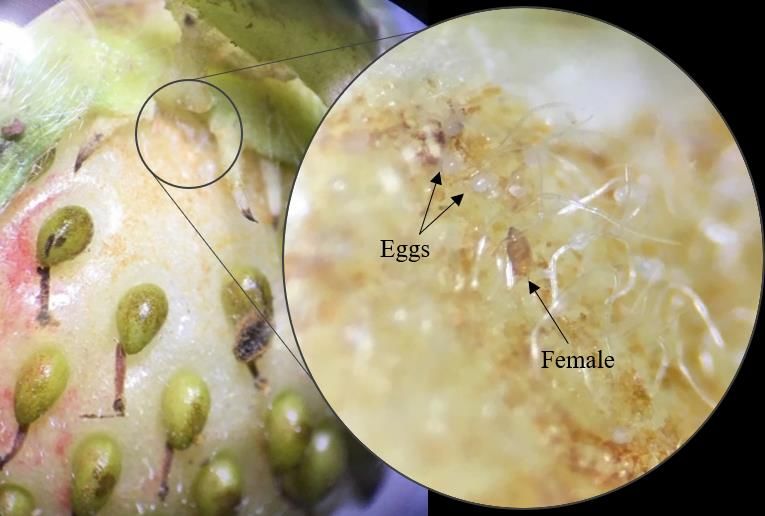
Abstract
The cyclamen mite Phytonemus pallidus Banks (Acari: Tarsonemidae) is a major pest in strawberry
production, causing serious yield losses. Due to the recent phase-out of endosulfan in Canada,
alternative treatments are required. Biological control with predatory mites (Acari: Phytoseiidae) is
promising, but its effectiveness under the conditions in Eastern Canada’s strawberry fields remains
unknown. The aim of this study was to (1) assess the potential of three phytoseiid mite species:
Neoseiulus cucumeris Oudemans, N. fallacis Garman and Amblyseius andersoni Chant to control
cyclamen mites in commercial strawberry fields in the province of Québec; (2) measure the effect of
this control method on fruit yield and quality; (3) evaluate predatory mite effects on other pests in the
field and (4) monitor field populations of cyclamen mites. In 2016, our results showed that
N. cucumeris was able to effectively suppress cyclamen mites but was too expensive for commercial
use and appeared to be too cold-sensitive to maintain adequate control throughout the season in this
region. In 2017, with an upper limit on the biocontrol cost ($1000/hectare), none of the predator
species achieved effective control. Monitoring cyclamen mite populations in the field has confirmed
some well-known facts on this pest and has led to the acquisition of new and essential knowledge on
its phenology. Based on these results, all three phytoseiid mites provided insufficient control of
cyclamen mites under the in-field conditions tested. Control of the cyclamen mite remains a very
complex issue and further research is needed.
iv
Table des matières
Résumé.................................................................................................................................. iii
Abstract ................................................................................................................................ iv
Table des matières ................................................................................................................ v
Liste des tableaux ............................................................................................................... vii
Liste des figures ................................................................................................................. viii
Remerciements ..................................................................................................................... xi
Avant-propos...................................................................................................................... xiii
Chapitre 1 : Introduction générale ..................................................................................... 1
1.1. La production de fraises ................................................................................................... 2
1.1.1. Historique .................................................................................................................... 2
1.1.2. Taxonomie et caractéristiques générales ..................................................................... 2
1.1.3. Statistiques de production ........................................................................................... 3
1.1.4. Régies de production ................................................................................................... 3
1.1.5. Ravageurs de la fraise ................................................................................................. 5
1.2. Le tarsonème du fraisier ................................................................................................... 5
1.2.1. Description .................................................................................................................. 5
1.2.2. Cycle de vie ................................................................................................................. 7
1.2.3. Dommages ................................................................................................................... 8
1.2.4. Ennemis naturels ....................................................................................................... 10
1.3. Lutte au tarsonème du fraisier ....................................................................................... 10
1.3.1. Moyens de lutte préventive ....................................................................................... 10
1.3.2. Lutte chimique........................................................................................................... 11
1.3.3. Moyens alternatifs d’assainissement des plants ........................................................ 15
1.3.4. La lutte biologique .................................................................................................... 16
1.4. Objectifs et hypothèses.................................................................................................... 30
Chapitre 2: Assessing the effectiveness of the predatory mite Neoseiulus cucumeris
(Acari: Phytoseiidae) in the control of the cyclamen mite Phytonemus pallidus (Acari:
Tarsonemidae) in Eastern Canada strawberry fields ................................................. 32
Résumé ......................................................................................................................................... 33
Abstract ........................................................................................................................................ 33
Introduction ................................................................................................................................. 34
Materials and Methods ............................................................................................................... 35
Field experiment........................................................................................................................ 35
Phenology of the cyclamen mite ............................................................................................... 38
Results .......................................................................................................................................... 38
Field experiment........................................................................................................................ 38
Phenology of the cyclamen mite ............................................................................................... 42
Discussion ..................................................................................................................................... 43
Neoseiulus cucumeris efficacy in the field ................................................................................ 44
Phenology of P. pallidus in the field ......................................................................................... 46
v
Acknowledgments ....................................................................................................................... 47
References .................................................................................................................................... 48
Chapitre 3: Biological control of the cyclamen mite Phytonemus pallidus (Acari:
Tarsonemidae) using three predatory mites (Acari: Phytoseiidae) in commercial
Eastern Canada strawberry fields .................................................................................... 51
Résumé ......................................................................................................................................... 52
Abstract ........................................................................................................................................ 52
Introduction ................................................................................................................................. 53
Materials and Methods ............................................................................................................... 54
Results .......................................................................................................................................... 58
Discussion ..................................................................................................................................... 62
Acknowledgments ....................................................................................................................... 65
References .................................................................................................................................... 66
Chapitre 4 : Conclusion générale ...................................................................................... 70
Bibliographie ....................................................................................................................... 75
vi
Liste des tableaux
Tableau 1. Matières actives des pesticides expérimentés et leur efficacité contre P. pallidus ........ 14
Table 2. Total number of phytoseiid mite species identified in the 2016 field experiment ............. 40
vii
Liste des figures
Figure 1. Champ de fraisiers en rangs nattés ...................................................................................... 4
Figure 2. Tarsonème du fraisier A- Femelle; B- Femelle, larves et œufs; C- Mâle ........................... 6
Figure 3. Dommages sévères du tarsonème du fraisier sur les feuilles de fraisier (gauche) et sur les
fruits (droite). .............................................................................................................................. 9
Figure 4. Étapes de la dispersion aérienne de N. fallacis. (A) Recherche aléatoire sur le substrat (B)
Orientation dans le sens du courant d’air dominant (vue dorsale); (C) Position redressée sur les
pattes postérieures, dos au courant d’air (af); (D) Décollage .................................................... 19
Figure 5. Cycle de vie d'un acarien phytoséiide typique à 27°C et 70 % HR.. ................................ 21
Figure 6. Neoseiulus cucumeris adulte. ............................................................................................ 24
Figure 7. Neoseiulus fallacis adulte ................................................................................................. 27
Figure 8. Amblyseius andersoni adulte............................................................................................. 29
Fig 9. Average numbers (±SE) of P. pallidus motile forms (Fig. a) and phytoseiid mite motile forms
(Fig. b) per young leaf for the acaricide, N. cucumeris and control treatments on each sampling
date for the 2016 field experiment in Sainte Foy, Quebec ........................................................ 39
Fig. 10. Average numbers of P. pallidus motile forms per young leaf (±SE in shaded area) for the
acaricide, N. cucumeris and control treatments on each sampling date for the 2016 field
experiment and follow-up in 2017 in Sainte Foy, Quebec ........................................................ 41
Fig. 11. Strawberry yield for the three treatments (Acaricide, N. cucumeris, and Control) at the
second-year follow-up (2017) of the 2016 field experiment in Sainte Foy, Quebec.. .............. 42
Fig. 12. Evolution in average number of P. pallidus females, larvae, males and eggs per young leaf
for 2016 and 2017 sampling dates in untreated plots in Sainte-Foy, Quebec. .......................... 43
Fig. 13. Experimental units (cages). ................................................................................................. 55
viii
Fig 14. Average number (±SE) of P. pallidus motile forms (Fig. a & b) and phytoseiid mite motile
forms (Fig c & d) per young leaf for the six treatments on each sampling date of the 2017 field
experiment 2017 on Île d’Orléans (left) and in Trois-Rivières (right), Québec. ....................... 59
Fig. 15. Picture of P. pallidus female and eggs on fruit.................................................................... 60
Fig. 16. Identification and total number of phytoseiid mites per treatment found on fruits, young
leaves and old leaves for each sampling date of the 2017 field experiment in Trois-Rivières. 61
Fig 17. Strawberry yield for the six treatments (Acaricide, A. andersoni, N. cucumeris, N. fallacis,
Control under cage and Control without cage) on Île d’Orléans, Québec.. .............................. 62
ix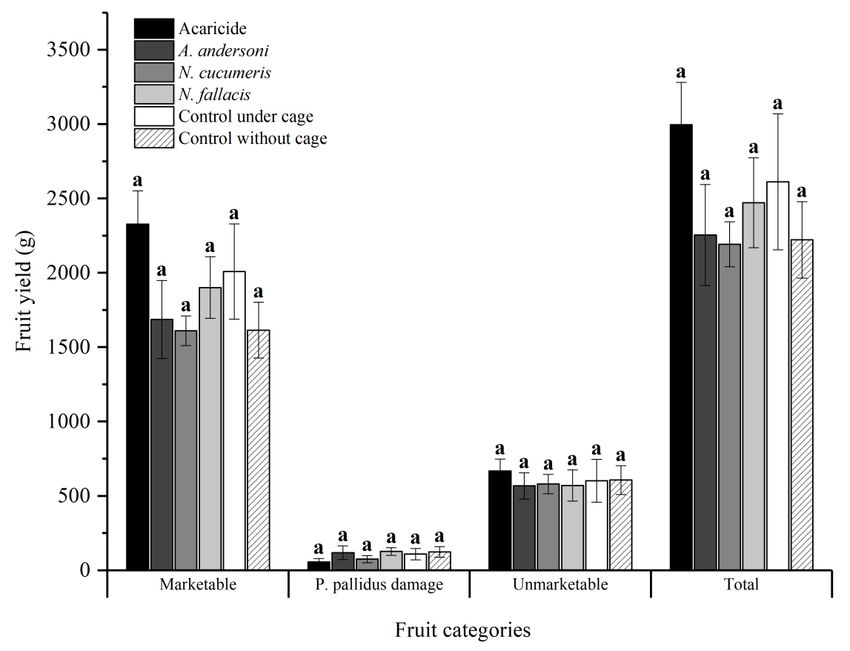
À ma famille x
Remerciements
« Oh mon dieu, dans quoi me suis-je embarquée… », voici le sentiment éprouvé lors de mes premiers
contacts avec le très problématique tarsonème du fraisier au début de cette maîtrise. Heureusement,
j’ai eu l’aide indispensable de nombreuses personnes dans cette aventure et ce projet est le fruit d’un
réel travail d’équipe. Je voudrais donc prendre le temps de les remercier.
Tout d’abord, je voudrais remercier ma directrice et ma codirectrice, Valérie Fournier et Stéphanie
Tellier, pour votre soutien, vos connaissances et vos conseils. Je suis également reconnaissante pour
cette confiance témoignée tout au long de cette maîtrise.
Ce projet aurait été impossible sans la participation, l’expertise et la générosité de François Demers,
présent depuis le tout début du projet, Thierry Chouffot, Diane Gagnon, Liette Lambert,
Guy-Anne Landry, Pierrette Lavoie, Jean-Philippe Légaré, Émilie Lemaire, Caroline
Martineau, Patrick Martineau, Joseph Moisan-De Serres, Annie-Pier Paradis, Alexandre
Tanguay, Patrice Thibault, Jacinthe Tremblay, Édith Tousignant et le personnel du CRIV, de
même qu’Awa Diop et Gaétan Daigle pour le soutien statistique. Un merci tout spécial pour
l’expertise taxonomique (et la grande patience) de Mario Fréchette du Laboratoire de diagnostic en
phytoprotection du Ministère de l’agriculture, des pêcheries et de l’alimentation (MAPAQ) et de
Frédéric Beaulieu d’Agriculture et agroalimentaire Canada (AAC), des atouts rares et indispensables
dans les contrées nébuleuses de l’acarologie.
Je voudrais remercier chaleureusement les producteurs Laurent Fiset, Jean-Julien Plante, David
Lemire et toute leur équipe pour votre grande disponibilité, votre collaboration et votre patience
(notamment dans la gestion des fameuses bâches de plastique). Ce fut un réel un plaisir de vous
côtoyer au cours de ces deux années.
Je tiens également à remercier les évaluateurs de ce mémoire pour leur temps et leurs commentaires
constructifs. Merci également à Daniel Dostaler, Raymond-Marie Duchesne, André Gosselin et
Véronique Martel pour l’évaluation des différents séminaires au cours de cette maîtrise.
Un grand merci à mes collègues de labo : Amélie Gervais (peintre en bourdons professionnelle),
Marianne Lamontagne-Drolet (Team agr., aux passions insolites), Frédéric McCune (l’inventeur
du McSquash), Sabrina Rondeau (l’être supérieur) et Mouna Kahia pour votre aide sur le terrain
(entre deux bouffées de moisissure grise), mais surtout pour votre présence, votre humour et les
anecdotes savoureuses en quantité industrielle à se radoter jusqu’à la fin de nos jours. Vous êtes tous
xide réels coups de cœur et cette maîtrise sans vous aurait été bien différente. Clin d’œil également aux
autres membres à distance des Charlie Angels, mais tout aussi importants : Mélanie Normandeau-
Bonneau et Phanie Bonneau. Merci aussi à Olivier Robert-Samson pour ton savoir infini et tes
apparitions de superhéros, brèves et intermittentes, mais toujours très appréciées.
Il ne faudrait pas oublier les indispensables aides de terrain qui ont dû apprendre, bien malgré eux, à
voir l’invisible dans ce projet. Tout d’abord, un grand merci à Audrey Boivin pour l’ensemble de
l’œuvre. Merci pour ta compétence, ta proactivité et ta capacité à lire dans mes pensées durant la
première année de ce projet. Merci également à Clémence Landreau et Aurélie Boilard d’avoir
brillamment pris le relais pour la 2e année, ainsi qu’à Thaïs Andro, Guillaume Guengard et Lucie
Alexandre pour leur aide supplémentaire sur le terrain.
Un grand merci à ma famille, ce que j’ai de plus précieux. À ma mère Isabelle pour ton soutien, ton
amour, ta résilience et ta grande force. À mon père Daniel pour ta confiance et ta présence rassurante,
malgré le fait que tu nous quittes maintenant peu à peu, beaucoup trop tôt. À mes sœurs et meilleures
amies Caroline et Lysane, de même qu’à ma belle-sœur Sarah, pour partager cette douce folie. À
mes nièces qui me permettent de redécouvrir la vie à travers leurs yeux et à ma grand-mère Monique,
une femme forte et fière avec qui je partage beaucoup de similarités, dont cette tête (un peu dure…)
pleine d’idées en constante ébullition.
Merci aussi à mes très bons amis Benoit Bérubé, Mélanie Dionne, Laura Thériault et Hugo Alix
pour votre présence dans ma vie. Merci pour votre aide, pour nos activités hétéroclites et pour cette
amitié, très précieuse, qui survit à tout.
Finalement, merci à tous les gens inspirants qui ont croisé ma route et qui ont contribué à faire de moi
ce que je suis aujourd’hui.
xiiAvant-propos
Ce mémoire constitue la finalité d’un projet rendu possible grâce à une subvention de recherche du
Ministère de l’agriculture, des pêcheries et de l’alimentation du Québec (MAPAQ) via le programme
Prime-Vert volet 4 - Appui au développement et au transfert de connaissances en agroenvironnement
par l’entremise de la Stratégie Phytosanitaire québécoise en agriculture 2011-2021, d’une
contribution supplémentaire du Programme d’appui au développement de l’agriculture et de
l’agroalimentaire en région (PADAAR) Mauricie et de l’implication des organismes suivants :
Association des producteurs de fraises et de framboises du Québec, Laboratoire d'expertise et de
diagnostic en phytoprotection du MAPAQ, Ferme Jean-Pierre-Plante, Ferme Ernest Fiset et fils,
Ferme horticole Gagnon, le Conseil de recherche en sciences naturelles et génie (CRSNG) pour le
financement d’une bourse d’études, ainsi que les distributeurs d’agents de lutte biologique : Koppert
Canada, Plant Products et Anatis bioprotection.
Les chapitres 2 et 3 constituent le corps du mémoire et sont rédigés en anglais sous forme d’articles
scientifiques qui seront soumis dans les journaux scientifiques Biocontrol et The Canadian
Entomologist. Les manuscrits intitulés «Assessing the effectiveness of the predatory mite Neoseiulus
cucumeris (Acari: Phytoseiidae) in the control of the cyclamen mite Phytonemus pallidus (Acari:
Tarsonemidae) in Eastern Canada strawberry fields» et «Biological control of the cyclamen mite
Phytonemus pallidus (Acari: Tarsonemidae) in commercial Eastern Canada strawberry fields :
comparison of three predatory mites (Acari: Phytoseiidae)» ont été écrits en collaboration avec
Valérie Fournier, PhD., professeure d'entomologie à l'Université Laval et Stéphanie Tellier, agr., M.
Sc., conseillère régionale petits fruits et experte provinciale dans le secteur des petits fruits au
MAPAQ de la direction régionale de la Capitale-Nationale, respectivement directrice et codirectrice
de ce projet de recherche. Les autres sections du mémoire, soit l’introduction générale présentée au
chapitre 1 ainsi que la conclusion générale constituant le chapitre 4, sont rédigées en français. Les
références citées dans ces sections sont présentées dans une bibliographie située immédiatement après
la conclusion générale.
La récolte des données, l’analyse des données ainsi que la rédaction de l’ensemble des textes ont été
réalisées par le candidat. Valérie Fournier (directrice) et Stéphanie Tellier (codirectrice) ont collaboré
aux textes et sont coauteures du chapitre 2 et 3.
xiiixiv
Chapitre 1 : État des connaissances
1.1. La production de fraises
1.1.1. Historique
Les fraises sont connues des humains depuis des millénaires. Consommées dès le Néolithique, elles
auraient fait partie de la composition des jardins des Grecs et des Romains (Hancock et Hancock
1999). À cette époque, il s’agissait principalement de l’espèce sauvage très parfumée et à petits fruits,
Fragaria vesca, la fraise des bois (Catoire 2004). Cette espèce a dominé en Europe jusqu’au début
des années 1600, jusqu’à l’arrivée de F. virginiana en provenance de l’Amérique du Nord. Cette
fraise écarlate, plus rustique et résistante aux températures froides, s’est répandue rapidement à
travers l’Europe (Husaini et Neri 2016). Toutefois, la fraise moderne majoritairement cultivée
aujourd’hui, F. x ananassa Duch., n’apparaîtra qu’un siècle plus tard. Cette espèce dotée de gros
fruits rouges caractéristiques doit sa création à un espion français, Amédée-François Frézier, qui
rapporta du Chili en 1714 un fraisier à gros fruits de couleur pâle, F. chiloensis. C’est l’hybridation
naturelle de F. virginiana et F. chiloensis dans un jardin de France qui a donné naissance à cette
nouvelle espèce, dont le nom provient de son parfum rappelant l’ananas. Elle fut rapidement adoptée
en raison de sa rusticité, de sa productivité et du calibre de ses fruits (Husaini et Neri 2016). Dans les
siècles qui ont suivi, plusieurs variétés ont été développées un peu partout dans le monde afin
d’améliorer constamment cette espèce.
1.1.2. Taxonomie et caractéristiques générales
La fraise est une plante herbacée pérenne de la famille des Rosacées et du genre Fragaria. Elle
possède une tige compacte appelée couronne et ses feuilles sont pétiolées et trifoliolées. Les plants se
reproduisent principalement par stolons. La fraise est composée d’un réceptacle charnu qui porte les
fruits, appelés akènes. La ploïdie des fraises est variable, allant de diploïde à décaploïde. Les espèces
américaines, dont fait partie F. x ananassa, sont généralement octaploïdes (Hancock et Hancock
1999). Plusieurs étapes de développement des fraisiers, dont la floraison, sont régies par la
photopériode et la température.
Il existe environ 34 espèces de Fragaria réparties en Amérique, Asie et Europe (Husaini et Neri
2016). Fragaria x ananassa demeure la plus cultivée à l’échelle mondiale avec un nombre important
de variétés qui ne cesse de croître. À l’échelle mondiale, 900 nouvelles variétés ont été développées
entre 1982 et 2008 (Della Strada et Fideghelli 2011). Bien qu’elle soit adaptée à une vaste gamme de
températures, la fraise est cultivée principalement dans les climats tempérés ou méditerranéens, soit
entre le 28e et le 60e parallèle (Hancock et Hancock 1999).
21.1.3. Statistiques de production
La production de fraises est importante à l’échelle mondiale. En 2014, plus de 8,1 millions de tonnes
étaient produites. Avec 38 % de la production mondiale, la Chine demeure le plus important pays
producteur, suivie par les États-Unis (17 %) et le Mexique (6 %). Le Canada arrive au 30e rang avec
une production de 20 210 tonnes, soit 0,2 % de la production mondiale (FAOSTAT 2016).
Avec ses 11 612 tonnes de fraises commercialisées en 2015, le Québec représente 57 % de la
production canadienne. Ceci touche 524 exploitations sur 1558 hectares en culture (ISQ et MAPAQ
2016). Les fraises fraîches demeurent le principal marché du Québec, suivies par les fraises dédiées
à la transformation et une petite proportion (4,8 %) est vouée à l’exportation (ISQ et MAPAQ 2016).
Bien que la demande pour les fruits et légumes biologiques soit en forte croissance mondialement, la
production de fraises biologiques est encore peu répandue (Willer et Lernoud 2016). En 2013,
seulement 21,8 hectares répartis sur 45 entreprises étaient dédiés à la culture de fraises biologiques
au Québec (CARTV 2018). Ceci représente environ 1 % de la production québécoise de fraise pour
la même année (ISQ et MAPAQ 2016). Toutefois, ce type de production suscite de plus en plus
d’intérêt chez les producteurs. C’est pourquoi le soutien à la recherche en production biologique fait
partie des priorités de recherche 2017 de l’Association des producteurs de fraises et framboises du
Québec (APFFQ 2017).
1.1.4. Régies de production
Au Québec, on retrouve principalement deux types de fraisiers, soit les fraisiers à jours courts et à
jours neutres. Les variétés à jours courts initient leurs bourgeons floraux à l’automne précédent et
fructifient habituellement vers la fin du mois de juin. Elles peuvent être utilisées selon les méthodes
culturales de plasticulture ou de rangs nattés, avec des plants de type « frigo », « motte » ou « frais »
(Thireau et Lefebvre 2014). Bien que la plasticulture offre une meilleure qualité de fruits et un calibre
plus gros, la régie dite en rangs nattés (Figure 1) demeure la plus répandue au Québec à ce jour
(Novafruit 2003). Selon cette régie, les plants à racines nues, soit « frais » ou « frigo », sont plantés
en plein champ au printemps et ne produisent des fruits que l’année suivante. Deux à trois années de
production suivent l’année d’implantation. Il est rare de voir une fraisière en production au-delà de
trois ans, car le calibre des fruits tend à diminuer avec les années et les problèmes phytosanitaires
sont plus fréquents, ce qui peut compromettre la rentabilité (Novafruit 2013). À la fin de l’automne,
les plants peuvent être protégés du froid par l’application d’une couche de paille appelée paillage ou
encore par l’utilisation de couvertures flottantes (Agriculture et agroalimentaire Canada 2015). Il est
3à noter que certaines couvertures flottantes, ou bâches d’automne, peuvent également permettre une
récolte plus hâtive l’année suivante en devançant la floraison (Thireau et Lefebvre 2014). Plusieurs
cultivars à jours courts sont disponibles au Québec et ceux-ci sont classés selon le moment de la
récolte dans la saison. Ainsi, ils peuvent être hâtifs, de mi-saison ou tardifs. Le cultivar de mi-saison
« Jewel » est un des plus connus dans la province.
Figure 1. Champ de fraisiers en rangs nattés. Photo: Stéphanie Patenaude
Les variétés à jours neutres, dites fraises d’automne, fleurissent tout au long de la saison. Les boutons
floraux sont donc initiés la même année que la floraison (Agriculture et agroalimentaire Canada
2015). Ces variétés permettent de récolter des fruits de juillet à octobre, soit hors de la saison
habituelle des fraises (Novafruit 2003). Le potentiel de rendement est donc supérieur. Toutefois, elles
font l’objet d’une régie plus intensive et sont généralement cultivées sur des buttes recouvertes de
plastique avec une irrigation au goutte-à-goutte (Agriculture et agroalimentaire Canada 2015).
Quelques cultivars sont disponibles au Québec, le plus cultivé étant le « Seascape ».
Outre l’utilisation de ces deux types de fraisiers, les producteurs utilisent différentes stratégies pour
étaler la production dans la saison ou pour augmenter les rendements et la qualité des fruits. Ainsi,
l’utilisation de bâches, de plastique noir, la plantation programmée ou encore l’utilisation de tunnels
font partie des stratégies possibles (Novafruit 2004). La culture hors-sol gagne également en
popularité, notamment en raison de la présence croissante des pathogènes de sol en champ. La culture
de fraises en serre, bien que de plus en plus de producteurs s’y intéressent, demeure encore marginale
au Québec en raison du climat et des coûts élevés des investissements (Novafruit 2003). Il existe donc
de multiples combinaisons possibles de méthodes de production selon les buts recherchés par les
4producteurs. Toutefois, peu importe la régie de production adoptée, les producteurs auront à composer
avec de multiples problèmes phytosanitaires, dont ceux associées à la présence de ravageurs de la
culture.
1.1.5. Ravageurs de la fraise
Les fraisiers sont sujets à plusieurs problèmes phytosanitaires. Outre les désordres abiotiques et les
maladies, plusieurs ravageurs affectent cette culture. Au Québec, la drosophile à ailes tachetées
(Drosophila suzukii), la punaise terne (Lygus lineolaris), l’anthonome de la fleur du fraisier
(Anthonomus signatus), le charançon noir de la vigne (Otiorhynchus sulcatus), le charançon de la
racine du fraisier (O. ovatus) et le tétranyque à deux points (Tetranychus urticae) sont bien présents
et causent beaucoup de dommages (Agriculture et agroalimentaire Canada 2015). Dans une moindre
mesure, les thrips (Frankliniella occidentalis et F. tritici) et les pucerons (plusieurs espèces) peuvent
être présents en pression modérée ou de façon sporadique. Le tarsonème du fraisier (Phytonemus
pallidus) est également très important puisqu’il démontre une forte pression et une présence annuelle
généralisée (Agriculture et agroalimentaire Canada 2015). Il est également préoccupant puisqu’il est
un ravageur d’importance à l’échelle mondiale, notamment en Europe et en Amérique du Nord
(Fountain et al. 2010; Hoy 2011b; Zhang 2003). Pour bien cerner ce problème, il incombe de mieux
connaître ce ravageur.
1.2. Le tarsonème du fraisier
1.2.1. Description
Le tarsonème du fraisier (Phytonemus pallidus Banks), antérieurement Tarsonemus pallidus (Banks),
Tarsonemus fragariae (Zimmerman) et Steneotarsonemus pallidus (Banks), est un acarien
phytophage de la famille des Tarsonemidae. Il s’agit d’une grande famille mondialement distribuée
qui possède 40 genres regroupant plus de 500 espèces (Zhang 2003). Le tarsonème du fraisier fait
partie de la sous-famille des Tarsoneminae. Sa présence aurait été remarquée pour la première fois
en 1898 à New York et en 1908 au Canada (Danemark 2000). Il s’agit d’un acarien minuscule
impossible à voir à l’œil nu qui fuit la lumière directe et qui nécessite une humidité relative élevée
pour compléter son cycle de vie (Hoy 2011b; Jeppson et al. 1975). Il démontre donc une préférence
pour les jeunes feuilles non déployées et les boutons floraux (Fitzgerald et al. 2008; Zhang 2003).
Il possède une large gamme d’hôtes, avec près de 60 plantes-hôtes potentielles (CABI Plantwise
2017). Outre le fraisier, il s’attaque également à plusieurs plantes ornementales, tels les bégonias, les
5Gerberas et les cyclamens (Hoy 2011b; Zhang 2003). C’est d’ailleurs l’origine de son nom anglais :
cyclamen mite. Certaines mauvaises herbes présentes au Québec pourraient également héberger ce
ravageur, notamment la petite oseille (Rumex acetosella), le galinsoga (Galinsoga parviflora), l’ortie
dioïque (Urtia dioica) et l’amarante à racine rouge (Amaranthus retroflexus) (CABI Plantwise 2017).
Le mâle et la femelle sont faciles à différencier puisqu’un dimorphisme sexuel important est présent
au sein de cette espèce. La femelle est brunâtre, ovoïde et mesure environ 250 µm (voir Figure 2).
Contrairement aux femelles, dont les pattes postérieures sont minces et se terminent en une soie
allongée, celles des mâles sont robustes et se terminent en pince (Alford 2007). Les œufs sont
elliptiques et mesurent 125 µm x 75 µm. Ils sont blanchâtres et translucides (Alford 2007). Lors de
fortes infestations, ils peuvent être visibles à l’œil nu (Zhang 2003). Les larves sont translucides,
blanchâtres et possèdent uniquement trois paires de pattes. Elles mesurent environ 200 µm x 80 µm
(Zhang 2003).
Figure 2. Tarsonème du fraisier A- Femelle (Photo : S. Patenaude) ; B- Femelle, larves et œufs (Photo : S. Patenaude);
C- Mâle (Photo : Karin Westrum/NIBIO).
Il est possible de confondre le tarsonème du fraisier avec le tarsonème trapu
(Polyphagotarsonemus latus Banks), un ravageur important en serre, notamment en horticulture
ornementale. Il s’agit d’une espèce très semblable à P. pallidus, causant des dommages comparables.
Toutefois, il est possible de différencier les deux espèces par la présence de tubercules sur les œufs
de P. latus, absents chez P. pallidus. Deux autres espèces de Tarsonemidae, soit Tarsonemus setifer
(Erwing) et T. confusus (Erwing), ont été retrouvées dans des fraisières de la Californie et pourraient
aussi porter à confusion lors des dépistages (Zalom et al. 2009). Ces espèces cosmopolites ne
semblent cependant pas occasionner de dommages aux plants. Il est possible de les différencier de P.
pallidus par l’absence de bride sur le fémur IV des mâles, par leur distribution plus uniforme sur la
feuille et par leurs œufs, pondus individuellement et non regroupés comme P. pallidus (Zalom et al.
62009). Cette espèce fongivore et ravageuse mineure occasionnelle de cultures ornementales serait
répandue un peu partout en Amérique du Nord, dont au Canada (Zhang 2003).
1.2.2. Cycle de vie
Au cours de son cycle de vie, le tarsonème traverse trois stades distincts, soit l’œuf, la larve et l’adulte.
Au printemps, les femelles adultes ayant hiverné profondément enfouies dans la couronne des plants,
émergent et commencent à se nourrir à la base des feuilles (Alford 2007; Jeppson et al. 1975). Peu de
temps après, elles se mettent à pondre. Au Canada, cette étape s’amorcerait vers la mi-avril (Jeppson
et al. 1975). Les femelles se reproduisent principalement par parthénogenèse, donc en absence de
reproduction sexuée. Certaines populations de P. pallidus sont dites thélytoques, signifiant que les
femelles non fécondées ne donneront que des femelles, alors que d’autres pourront donner à la fois
des mâles et des femelles (Zhang 2003). La ponte se fait à un rythme moyen de 1,8 à 2,0 œufs par
jour. Au cours de sa vie (jusqu’à 45 jours), une femelle peut pondre en moyenne 24,3 œufs à 20°C et
28,5 œufs à 25°C, pour un maximum de 49 œufs (Easterbrook et al. 2003). Ces œufs sont pondus
entre les poils épidermiques de la plante, généralement à l’intérieur des bourgeons foliaires et floraux,
des jeunes feuilles ou encore entre deux folioles adjacentes non déployées (Alford 2007; Zhang 2003).
Les œufs peuvent prendre de quatre jours à 20°C à 13 jours à 15°C pour éclore, donnant place aux
larves (Zhang 2003). Celles-ci vont se nourrir d’un à quatre jours avant d’entrer dans le stade
quiescent (Zhang 2003). C’est à ce moment que s’amorcera la transformation au stade adulte à
l’intérieur de leur cuticule (Alford 2007). Les mâles, lorsque présents, sont reconnus pour avoir un
comportement de « pré-copulation » qui consiste à guetter et transporter sur leur dos des larves
femelles. Ce comportement peut durer jusqu’à 24 heures et dès que la femelle émerge, la vraie
copulation a lieu (Zhang 2003). Le temps de développement du tarsonème est inversement
proportionnel à la température. Un cycle peut être complété en 28,4 jours à 12,5°C et en aussi peu
que 8,8 jours à 25°C (Easterbrook et al. 2003). Le tarsonème aime les températures fraîches.
Toutefois, les larves ne peuvent se transformer en adulte lorsque les températures sont sous 10°C
(Easterbrook et al. 2003). Aucune étude ne rapporte leur diapause et en serre, ils peuvent se reproduire
à l’année (Zhang 2003).
Peu d’études se sont attardées sur la dynamique de population de P. pallidus en champ. Une des rares
études a eu lieu dans l’État de New York, dans les années 60. Dans une fraisière en première année
de production, on dénotait une augmentation des densités de la population lors de la floraison, soit
dès la fin du mois de mai, pour atteindre un pic au début du mois de juillet. Par la suite, un déclin
rapide survenait en juillet et août. Un deuxième pic plus modeste se manifestait à la fin septembre.
7Finalement, les densités de la population atteignaient leur creux saisonnier à la fin octobre (Schaefers
1963). Les chercheurs affirment toutefois que les pics et creux seraient davantage reliés à la croissance
végétative des fraisiers. En effet, lorsque le taux de croissance est faible, notamment lors de la
floraison et de la fructification, les tarsonèmes se retrouvent concentrés sur un nombre limité de jeunes
feuilles. Ceci occasionne une plus grande détection lors des décomptes. Cette concentration peut aussi
entraîner une forte mortalité des tarsonèmes en raison de la détérioration de la feuille plus rapide que
son renouvellement. Ceci expliquerait la baisse des densités de la population qui suit invariablement
un pic, notamment à la fin juin. Les creux peuvent également être dus à l’effet de dilution lors de
fortes croissances végétatives, comme au printemps ou après la récolte. Les tarsonèmes sont alors
davantage dispersés à travers les feuilles. Finalement, cette baisse peut également être partiellement
expliquée par l’action de prédateurs indigènes, détectés en plus forte importance de juin à octobre
(Schaefers 1963). Il est donc difficile d’avoir la véritable dynamique de population du tarsonème du
fraisier, lorsqu’observé en champ. Le suivi du champ par les chercheurs l’année suivante, soit lors de
la deuxième année de production, a démontré la même tendance, mais avec des densités de
tarsonèmes plus élevées. Au Canada, le pic de population surviendrait de la mi-juin à la fin juillet,
moment pendant lequel jusqu’à 300 tarsonèmes par plant peuvent être présents. La ponte des œufs
diminuerait en août pour se terminer en octobre (Jeppson et al. 1975). En Europe, le pic surviendrait
davantage vers le mois d’août ou septembre (Alford 2007).
1.2.3. Dommages
Les tarsonèmes du fraisier vont se nourrir sur les tissus tendres à l’abri de la lumière, principalement
à la base des nouvelles folioles (Schaefers 1963). Les dommages sont causés, d’une part, par leur
alimentation qui entraîne une nécrose et une distorsion du mésophylle. D’autre part, ils injecteraient
une toxine causant une hypertrophie et une prolifération des cellules épidermiques (Schaefers 1963).
Ces changements irréversibles donnent un aspect tordu et cloqué à la feuille (figure 3). Les boutons
floraux en développement voient leur base noircir sous l’attaque de ces acariens.
Les tarsonèmes du fraisier se nourrissent à l’intérieur des jeunes feuilles jusqu’à ce qu’elles s’ouvrent
et s’endurcissent. Par la suite, ils migreront pour trouver un nouveau site d’alimentation sur le même
plant ou sur les plants adjacents. Pour se faire, ils se déplaceront par le feuillage, si les feuilles se
touchent, ou par les stolons. Ils éviteront toutefois le sol, car ils sont très sensibles à la dessiccation
(Hoy 2011b). En effet, la mort de ceux-ci surviendrait en quelques jours sur un sol sec ou en seulement
quelques heures lorsqu’exposés aux rayons du soleil (Jeppson et al. 1975). Ils peuvent également être
8dispersés de façon passive, soit via d’autres insectes, par la machinerie, les cueilleurs, l’eau
d’irrigation ou encore par les vents (Alford 2007; Hoy 2011b; Jeppson et al. 1975).
Figure 3. Dommages sévères du tarsonème du fraisier sur les feuilles de fraisier (gauche) et sur les fruits (droite).
Photos : Stéphanie Patenaude
Les manifestations et la gravité des symptômes peuvent varier selon la densité de tarsonèmes présents.
Le taux de croissance de la plante joue également un rôle dans la sévérité des dommages. En effet,
plus la croissance est lente, plus les tarsonèmes profitent longtemps d’un environnement favorable à
leur développement. Ceci entraîne donc des dommages plus importants (Schaefers 1963). La variété
de fraisiers peut également influencer la manifestation des symptômes, puisque les dommages
observés ne sont pas toujours corrélés de la même façon à la quantité de tarsonèmes présents. Ainsi,
les cultivars possédant un haut taux de croissance et une forte pression osmotique seraient plus
résistants à l’attaque de ces acariens (Schaefers 1963). Toutefois, aucun cultivar n’est actuellement
résistant à ce ravageur (Łabanowska 2004).
Très peu d’études ont quantifié les dommages de P. pallidus. La plus connue demeure celle de
Stenseth et Nordby (1976). Une population inférieure à huit tarsonèmes par foliole en post-récolte
n’affecterait pas le rendement en fruit l’année suivante. Toutefois, une population plus importante,
soit 34 et 44 acariens par foliole, entraînerait des pertes de rendement de 23 % et 33 % respectivement
(Stenseth et Nordby 1976). En termes de rendement en fruits, aucune différence significative n’a été
décelée pour le nombre d’inflorescences par plant. Toutefois, le nombre de fleurs par inflorescence
était moindre lors de fortes infestations de P. pallidus. De plus, les fruits étaient plus petits pour les
traitements où des tarsonèmes avaient été introduits par rapport au traitement sans tarsonèmes et au
traitement acaricide. La baisse de rendements constatée serait due en partie à l’alimentation de
P. pallidus lors de l’initiation des boutons floraux, soit après la récolte de l’année précédente. Quant
9à la réduction du calibre des fruits, elle serait davantage causée par une infestation avant la récolte,
soit en diminuant la vigueur des plants ou en attaquant directement les fleurs (Stenseth et Nordby
1976). Cette étude n’a pas dénoté de différence dans la mise à fruit, alors que Schaerfers (1963)
affirmait le contraire.
Aucun seuil d’intervention contre ce ravageur n’a été répertorié au Canada, comme c’est le cas pour
la quasi-totalité des pays où il est présent. Toutefois, la Pologne s’est fixé un seuil de deux à trois
formes mobiles de P. pallidus par plus jeune feuille trifoliolée en post-récolte (Łabanowska et al.
2015). Au-delà de ce seuil, un traitement est recommandé. Il est à noter que les variétés tardives (mi-
et fin de saison) ainsi que celles à jours neutres sont plus susceptibles d’être infestées par le tarsonème
du fraisier. C’est également le cas lorsque des abris ou des protections sont utilisés dans la régie de
culture (Fountain et al. 2010). Cela s’explique en partie par la création de conditions favorables à
celui-ci plus longtemps en saison.
1.2.4. Ennemis naturels
Dans la nature, le tarsonème du fraisier possède des prédateurs naturels, bien que le phénomène soit
encore peu documenté. Parmi ceux-ci se trouvent le thrips prédateur Scolothrips sexmaculatus
(Pergande) ainsi que la punaise prédatrice Orius sp. (Solomon et al. 2010). Les acariens de la famille
des phytoséiides, principalement les genres Amblyseius et Neoseiulus, sont également d’excellents
prédateurs du tarsonème du fraisier (Solomon et al. 2010). Des champignons entomopathogènes,
principalement du genre Hirsutella, peuvent infecter les acariens Tarsonemidae (Miętkiewski et al.
2000). Hirsutella nodulosa Petch a d’ailleurs été identifié en Angleterre sur des tarsonèmes du fraisier
morts (Alford 1979). En conditions naturelles, la mortalité de P. pallidus résultant de ce champignon
oscille entre 0,6 % et 13,0 %, le maximum étant atteint à l’automne (Tkaczuk et al. 2004). Cependant,
ces agents fongiques peuvent également infecter les acariens prédateurs (Miętkiewski et al. 2000).
De plus, certains pesticides utilisés dans la culture de la fraise ont un effet négatif sur la croissance
de ce champignon (Tkaczuk et al. 2004). Il est donc plausible de penser que le contrôle de P. pallidus
par celui-ci demeure marginal en conditions de fraisières commerciales.
1.3. Lutte au tarsonème du fraisier
1.3.1. Moyens de lutte préventive
La lutte au tarsonème du fraisier passe d’abord par la prévention. À cet effet, Hoy (2011b) a dressé
une liste de moyens de lutte préventive à intégrer dans une optique de lutte intégrée. Tout d’abord,
10des mesures de prophylaxie, telles que la destruction des résidus de culture infestés et l’élimination
des mauvaises herbes hôtes, doivent être adoptées. Ensuite, l’équipement et la machinerie devraient
être nettoyés régulièrement pour éviter le déplacement par inadvertance de tarsonèmes d’un champ
infesté vers un champ sain. Une attention particulière devrait être portée à la fertilisation. En effet,
celle-ci doit être adéquate, car un surplus favorise les infestations d’acariens. De plus, il est essentiel
que le choix des pesticides soit fait dans le but de préserver au maximum les ennemis naturels déjà
présents. Finalement, il est primordial de débuter la production avec des plants de fraisiers exempts
de tarsonèmes et des cultivars moins sensibles à ce ravageur, lorsque disponibles.
1.3.2. Lutte chimique
La lutte chimique a été et demeure la plus utilisée pour contrôler le tarsonème du fraisier.
Historiquement, les producteurs obtenaient un bon contrôle avec des pulvérisations d’endosulfan
(Thiodan®, Thionex®) effectuées principalement en post-récolte, après la tonte du feuillage
(rénovation de la fraisière). Cet insecticide/acaricide à large spectre de la famille des cyclodiènes
chlorés (groupe 2A) s’est montré le plus efficace pour lutter contre ce ravageur à maintes reprises
(Labanowska 1992; Schaefers 1963; Stenseth et Nordby 1976; Tanigoshi et Bergen 2005). Toutefois,
sa toxicité et ses effets nocifs pour la santé humaine ont forcé son abandon graduel. D’abord retiré de
l’Union européenne en 2007, c’est au tour de l’Agence de Protection de l’Environnement des États-
Unis (EPA) d’annoncer, en 2010, son bannissement progressif. Ceci en raison des « risques
inacceptables aux systèmes neurologique et reproducteur des agriculteurs et de la faune, de même
que pour sa persistance dans l’environnement » (EPA 2010). Au Canada, l’Agence de réglementation
de la lutte antiparasitaire (ARLA) a suivi le pas et amorcé l’abandon graduel de ce pesticide en 2011,
aboutissant à son interdiction complète le 31 décembre 2016 (Gouvernement du Canada 2011).
L’endosulfan a d’ailleurs été placé à l’Annexe A des polluants organiques persistants de la
Convention de Stockholm en 2011, obligeant ainsi son élimination à l’échelle mondiale. Dans la
foulée de ce retrait, plusieurs équipes de recherche ont tenté de trouver des acaricides de
remplacement à l’endosulfan. Le tableau 1 résume les principales matières actives expérimentées
ainsi que l’efficacité obtenue. Toutefois, ces résultats sont difficiles à généraliser en raison des
nombreuses disparités entre les études en ce qui a trait aux doses et aux conditions d’application. En
effet, comme l’efficacité du traitement dépend beaucoup de ces conditions, ceci explique la grande
variabilité des résultats.
Les acaricides agissent principalement par contact. Cependant, réussir à atteindre ce ravageur est un
défi de taille. Tout d’abord, cet acarien est protégé à l’intérieur des bourgeons ou des jeunes feuilles,
11Vous pouvez aussi lire

















































