ORIGIN AND ONTOGENY OF HUMAN LUNG MACROPHAGES - From Department of Medicine, Huddinge Karolinska Institutet, Stockholm, Sweden
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
From Department of Medicine, Huddinge
Karolinska Institutet, Stockholm, Sweden
ORIGIN AND ONTOGENY OF HUMAN
LUNG MACROPHAGES
Elza Evren
Stockholm 2022
All previously published papers were reproduced with permission from the publisher. Published by Karolinska Institutet. Printed by Universitetsservice US-AB, 2022 © Elza Evren, 2022 ISBN 978-91-8016-647-8 Cover illustration: Digital collage of MISTRG mice lungs stained with anti-human CD68 antibody. The branches of the tree represent the differentiation process from fetal and adult progenitors to lung macrophages depicted as iridescent flowers. Designed by Elza Evren.
ORIGIN AND ONTOGENY OF HUMAN LUNG
MACROPHAGES
THESIS FOR DOCTORAL DEGREE (Ph.D.)
By
Elza Evren
The thesis will be defended in public at Karolinska Institutet, room 4U Solen, Alfred Nobels
Allé 8, Floor 4 (entrance floor) on Friday 10th of June 2022 at 13:30.
Principal Supervisor: Opponent:
Associate Professor Tim Willinger Associate Professor Claudia Jakubzick
Karolinska Institutet Darthmouth College
Department of Medicine, Huddinge Department of Microbiology and Immunology
Center for Infectious Medicine
Co-supervisor(s): Examination Board:
Associate Professor Anna Smed-Sörensen Professor Martin Rottenberg
Karolinska Institutet Karolinska Institutet
Department of Medicine, Solna Department of Microbiology, Tumor and
Division of Immunology and Allergy Cell Biology (MTC)
Associate Professor Apostolos Bossios Professor William Agace
Karolinska Institutet Lund University
Department of Medicine, Huddinge Immunology Section
Hjärt och lungsjukdomar
Associate Professor Katharina Lahl
Lund University
Immunology Section

To my grandmother Emine,


Homo sum, humani nihil a me alienum puto.
Terence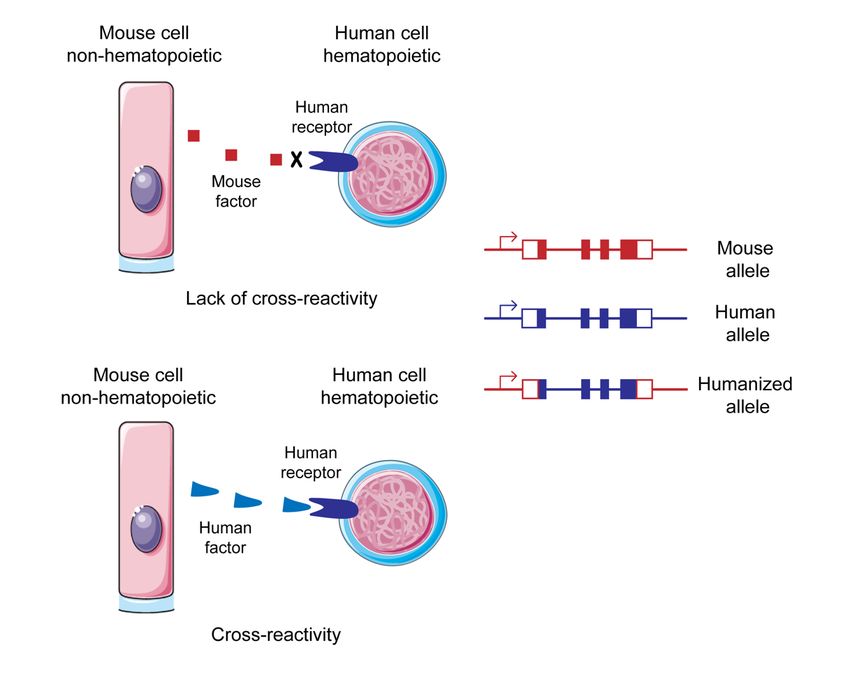
POPULAR SCIENCE SUMMARY OF THE THESIS What do starfish and this thesis have in common? They both contributed to macrophage biology! Macrophages are cells that are part of the immune system whose role is to defend our body against microbes. They have been discovered in starfish larvae and named after their most defining function: phagocytosis. Phagocytosis is the cellular process by which macrophages attach to, cover and engulf virus, bacteria and fungi. Each organ harbors its own type of macrophages defined according to their localizations and functions. One could imagine macrophages as a trustworthy friend to whom you leave your keys when going travelling. They will tidy around in your apartment and keep it clean, stop any unwanted visitors from entering and only if they cannot, they will call the police for help. From the first breath we inhale, our lungs are constantly exposed to airborne pathogens and particles. It is very important to keep the lung environment clear of microbes but at the same time avoid excessive inflammation which might damage the tissues. Alveolar macrophages (AMs) are the predominant cell type in the airways, where their main duty is to recycle lung surfactant, a thin layer of mucus that serves as lubricant to the alveolar surface. In addition, they protect the lungs by phagocytosing potential intruders. AMs are involved in many lung diseases such as asthma, tuberculosis, chronic obstructive pulmonary diseases and the now infamous coronavirus disease 2019. The origin of different macrophages residing in the lungs is unknown as it is very challenging to study these cells in humans. The common belief in the field was that all macrophages derive from circulating blood monocytes. Monocytes are immune cells which develop in the bone marrow from stem cells. They egress from the bone marrow into the bloodstream and can migrate to the organs according to the signals they receive. Studies conducted in mice triggered a paradigm shift by showing that mouse AMs derive from embryonic progenitors which seed the lungs and give rise to AMs right after birth, and not from blood monocytes as previously thought. In this thesis, we defined the progenitors of human lung macrophages and we showed that they can derive both from embryonic precursors and adult blood monocytes. Even though macrophages from different origins had similar gene signatures, we still noted some potential differences in their function. For example, monocyte-derived macrophages seemed to be involved in inflammatory responses to viruses and they might contribute to disease severity and prolongation. Our results were particularly relevant in a viral disease context such as SARS-CoV-2 infection, in which monocyte derived-macrophages have a harmful role.
POPULÄRVETENSKAPLIG SAMMANFATTNING AV AVHANDLING Vad har sjöstjärnor och denna avhandling gemensamt? De bidrog båda till makrofagbiologin! Makrofager är celler som är en del av immunsystemet, vars roll är att försvara vår kropp mot mikrober. De har upptäckts i sjöstjärnans larver och fått namn efter deras mest definierande funktion: fagocytos. Fagocytos är den cellulärprocess genom vilken makrofager fäster vid, täcker och uppslukar virus och bakterier. Varje organ har sin egen typ av makrofager definierade enligt deras lokaliseringar och funktioner. Man skulle kunna föreställa sig makrofager som en pålitlig vän som du lämnar dina nycklar till när du reser bort. De kommer att städa i din lägenhet, stoppa eventuella oönskade besökare från att komma in och, om de inte kan hindra inbrottet så kallar de på polisen för hjälp. Från vårt första andetag utsätts vårar lungor ständigt för luftburna patogena och partiklar. Det är mycket viktigt att hålla lungmiljön fri från mikrober men också att samtidigt undvika överdriven inflammation som kan skada lungans vävnader. Alveolära makrofager (AM) är den dominerande celltypen i luftvägarna, där deras huvudsakliga uppgift är att återvinna lungsurfaktant, ett tunt lager av slem som fungerar som smörjmedel till alveolytan. Dessutom skyddar de lungorna genom att fagocytera potentiella inkräktare. AM är involverade i många lungsjukdomar som astma, tuberkulos, kronisk obstruktiv lungsjukdom och den nu ökända COVID- 19. Ursprunget till olika makrofager som finns i lungorna är okänt eftersom det är mycket utmanande att studera dessa celler hos människor. Den vanliga uppfattningen inom området var att alla makrofager härrör från cirkulerande blodmonocyter. Monocyter är immunceller som utvecklas i benmärgen från stamceller. De kommer ut från benmärgen till blodomloppet och kan migrera till organen enligt de signaler de får. Studier utförda på möss utlöste ett paradigmskifte genom att visa att mus-AMs härrör från embryonala stamfäder som sår lungorna och ger upphov till AMs direkt efter födseln, och inte från blodmonocyter som man tidigare trott. I denna avhandling definierade vi stamfäder till mänskliga lungmakrofager och vi visade att de kan härröra både från embryonala prekurser och vuxna blodmonocyter. Även om makrofager från olika ursprung hade liknande gensignaturer, noterade vi fortfarande några skillnader i deras funktion. Till exempel verkade monocythärledda makrofager vara involverade i inflammatoriska svar på virus och de kan bidra till sjukdomens svårighetsgrad och förlängning. Våra resultat var särskilt relevanta i ett virussjukdomssammanhang som SARS-CoV-2- infektion, där monocythärledda makrofager har en skadlig roll.
RÉSUMÉ DE RECHERCHE VULGARISÉ Quel est le point commun entre une étoile de mer et cette thèse? Ils ont tous les deux contribué à la recherche sur les macrophages! Les macrophages sont des cellules du système immunitaire dont le rôle est de défendre notre corps contre les microbes. Ils ont été observés pour la première fois dans des larves d’étoiles de mer et nommés d’après leur fonction la plus caractéristique: la phagocytose. La phagocytose est le processus biologique par lequel les macrophages engloutissent les virus et les bactéries. On retrouve des macrophages spécifiques à chaque organe dans le corps et ils sont définis selon leurs localisations et leurs fonctions. On pourrait imaginer les macrophages comme des amis de confiance à qui l’on laisse les clés de sa maison quand on part en vacances. Ils rangeront votre appartement tout en le gardant bien propre, le protègeront contre des visiteurs indésirables et dans le cas où ils n’y arriveraient pas tous seuls, ils appelleront la police à la rescousse. De notre premier souffle jusqu’au dernier, nos poumons sont constamment exposés aux agents pathogènes présents dans l’air. Il est très important d’empêcher l’entrée des microbes dans le corps à travers les poumons tout en évitant de déclencher une réaction inflammatoire excessive qui pourrait endommager le tissu pulmonaire. Les macrophages alvéolaires sont les cellules les plus nombreuses dans les voies respiratoires où ils protègent les poumons en ingérant les potentiels intrus par phagocytose. Ces macrophages sont impliqués dans de nombreuses maladies pulmonaires telles que l’asthme, la tuberculose, la maladie pulmonaire obstructive chronique et rajoutée récemment à la liste, la maladie à coronavirus 2019. L’origine des macrophages résidant dans les poumons est inconnue car il est très difficile d’étudier ces cellules chez l’homme. Pendant longtemps, les scientifiques ont pensé que tous les macrophages dérivaient des monocytes circulant dans le sang. Ces monocytes sont des cellules immunitaires qui se développent dans la moelle osseuse à partir de cellules souches. Des études menées chez les souris ont montrés que les macrophages alvéolaires de souris dérivaient de précurseurs embryonnaires qui migrent vers les poumons avant la naissance, et non de monocytes du sang comme on l’assumait auparavant. Cette thèse a pour but de définir l’origine des macrophages pulmonaires chez l’homme. Nos résultats montrent que ces cellules peuvent dériver à la fois de précurseurs embryonnaires mais aussi de monocytes du sang. Bien que les macrophages d'origines différentes aient des signatures génétiques similaires, nous avons tout de même noté certaines différences dans leur fonction. Par exemple, les macrophages dérivés de monocytes semblent être spécialisés dans la réponse immunitaire aux virus et ils pourraient contribuer à la gravité et à la prolongation de la maladie. Nos résultats sont particulièrement importants dans un contexte de maladie virale telle que l'infection par le SRAS-CoV-2 dans laquelle les macrophages dérivés de monocytes ont un rôle néfaste.
TEZİN POPÜLER BİLİM OZETİ Bu tezin bir deniz yıldızıyla ortak noktası nedir? İkisi de makrofaj bilimine katkıda bulundu! Makrofaj adı verilen hücreler, görevi vücudumuzu mikroplara karşı savunmak olan bağışıklık sisteminin bir parçasıdır. İlk olarak deniz yıldızı larvalarında keşfedilmişlerdir ve adlarını en önemli işlevleri olan fagositozdan alırlar. Fagositoz, makrofajların virüs ve bakterileri yakalayıp yutmasını sağlayan biyolojik işlemdir. Vücudumuzdaki her organda değişik bir makrofaj tipi bulunur. Makrofajları tatile giderken evinizin anahtarlarını emanet ettiğiniz güvenilir bir arkadaşınız olarak düşünebilirsiniz. Siz yokken evinizi toplarlar ve temizlerler, istenmeyen misafirlerin evinize girmesini engellemekte üstlerine yoktur, ve diyelim ki engelleyemediler, o zaman da hemen polisi ararlar. Aldığımız ilk nefesten son nefesimize kadar, akciğerlerimiz sürekli havada bulunan mikropların saldırısı altındadır. Bir yandan akciğerleri mikroplardan korumak, diğer yandan ise akciğer dokusuna zarar verebilecek aşırı iltihaplanmayı önlemek gerekmektedir. Akciğer makrofajları solunum yollarında en sıklıkla bulunan hücre tipidir. Solunum yollarındaki mikropları fagositoz yoluyla yok ederler, ancak işlevleri fagositoz ile sınırlı değildir. Bu hücreler astım, tüberküloz ve kronik obstrüktif akciğer hastalığında önemli rol oynarlar. Ne yazık ki bu listeye son yıllarda koronavirüs hastalığı da eklenmiştir. Bu hücreleri insanlarda incelemek çok zor olduğundan, akciğerlerde bulunan farklı makrofajların kökeni bilinmemektedir. Yakın zamana kadar tüm makrofajların kan dolaşımında bulunan monositlerden türediği düşünülmekteydi. Monositler, kemik iliğinde kök hücrelerden gelişen bağışıklık sistemi hücreleridir. Kemik iliğinden kan dolaşımına akarlar ve aldıkları sinyallere göre organlara göç ederler. Son yıllarda farelerde yapılan araştırmalar akciğer makrofajlarının daha önce sanıldığı gibi kan monositlerin değil, embriyonik kök hücrelerden ürediğini kanıtladı. Bu tezde insan akciğer makrofajlarının kökenini araştırdık ve bu hücrelerin hem embriyonik kök hücrelerden hem de yetişkin kan monositlerinden ürediğini gösterdik. Farklı kökenlerden gelen makrofajlar arasında büyük benzerlikler görmüş olmamıza rağmen, işlevlerinde bazı farklılıklar kaydettik. Örneğin, monosit kökenli makrofajlar vücudumuzun virüslere karşı verdiği tepkilerde rol oynuyor ve hastalığın şiddetine ve uzamasına katkıda bulunuyorlar. Bulgularımız, özellikle monosit kökenli makrofajların zararlı bir role sahip olduğu koronavirüs hastalığı kapsamında ciddi önem taşımaktadır.
ABSTRACT The lung is a vital organ occupied by a large diversity of immune cells due to its close proximity with the external environment. This thesis focuses on a particular type of immune cell: the macrophage. The lungs are composed of three separate anatomical compartments: the airways that start from the trachea and end at the alveoli, the blood capillaries and the connective tissue supporting the lung structure. Correspondingly, three macrophage subsets have been described in the lungs: alveolar, intravascular and interstitial macrophages. Contrary to the reigning dogma that macrophages were generated from circulating monocytes, studies in mice demonstrated that fetal monocytes seeding the lungs during embryonic stages gave rise to alveolar macrophages after birth. The origin of lung macrophages in humans remains unknown as invasive experiment to track cellular origin in vivo are ethically impossible to conduct. To face this challenge, we used the MISTRG humanized mouse model. MISTRG mice support the development of human myeloid cells (especially human monocytes and macrophages). MISTRG mice were transplanted intrahepatically, intranasally and intravenously with different adult and embryonic precursors to assess their macrophage progenitor capacity as well as migration potential. In addition, we performed intravascular cell labelling, bead-based fate-mapping, microarray gene expression analysis, bulk and single-cell RNA sequencing to investigate the precise localization of macrophage progenitors and the heterogeneity of the human lung macrophage compartment. We demonstrated in Paper I that two distinct developmental pathways from circulating blood monocytes exist: CD14+CD16- classical monocytes give rise to interstitial and alveolar macrophages while their CD14lowCD16+ non-classical counterparts generate pulmonary intravascular macrophages. In Paper II, we defined the CD116+CD64- human fetal progenitor of alveolar macrophages, resembling Myb-expressing erythro-myeloid progenitors (EMPs) and CD64- myeloid progenitors derived from EMPs in mice. These fetal progenitors had a highly proliferative gene expression profile and were consequently better at occupying the alveolar niche than the adult monocytes. Gene expression analysis of alveolar macrophages of adult and embryonic origin revealed that the environment had a strong impact on macrophage function as very few genes were differentially expressed between alveolar macrophages of different origin. However, adult precursors preferentially generated interferon-responsive macrophages, similar to a subset expanding in inflammatory disease in humans such as COVID-19. This thesis provides new insights into the origin and development of human lung macrophages, thereby contributing to the development of macrophage-based therapies for lung diseases in humans.
LIST OF SCIENTIFIC PAPERS
I. Elza Evren, Emma Ringqvist, Kumar Parijat Tripathi, Natalie Sleiers, Ines Có
Rives, Arlisa Alisjahbana, Yu Gao, Dhifaf Sarhan, Tor Halle T, Chiara Sorini,
Rico Lepzien, Nicole Marquart, Jakob Michaëlsson, Anna Smed-Sörensen,
Johan Botling, Mikael C.I. Karlsson, Eduardo J. Villablanca, and Tim
Willinger. Distinct developmental pathways from blood monocytes generate
human lung macrophage diversity. Immunity. 2021;54, 1–17.
II. Elza Evren, Emma Ringqvist, Jean-Marc Doisne, Anna Thaller, Natalie
Sleiers, Richard A. Flavell, James P. Di Santo, and Tim Willinger. CD116+
fetal precursors migrate to the perinatal lung and give rise to human alveolar
macrophages. Journal of Experimental Medicine. 2022; 219 (2).SCIENTIFIC PAPERS NOT INCLUDED IN THE THESIS
I. Johanna Emgård, Hana Kammoun, Bethania Garcia-Cassani, Julie Chesne,
Sara M. Parigi, Jean-Marie Jacob, Hung-Wei Cheng, Elza Evren, Srustidhar
Das, Paulo Czarnewski, Natalie Sleiers, Felipe Melo-Gonzalez, Egle
Kvedaraite, Mattias Svensson, Elke Scandella, Matthew R. Hepworth, Samuel
Huber, Burkhard Ludewig, Lucie Peduto, Eduardo J. Villablanca, Henrique
Veiga-Fernandes, João P. Pereira, Richard A. Flavell, and Tim Willinger.
Oxysterol Sensing through the Receptor GPR183 Promotes the Lymphoid-
Tissue-Inducing Function of Innate Lymphoid Cells and Colonic
Inflammation. Immunity. 2018;48(1):120-32 e8.
II. Felipe Melo-Gonzalez, Hana Kammoun, Elza Evren, Emma E. Dutton,
Markella Papadopoulou, Barry M. Bradford, Ceylan Tanes, Fahmina Fardus-
Reid, Jonathan R. Swann, Kyle Bittinger, Neil A. Mabbott, Bruce A. Vallance,
Tim Willinger, David R. Withers and Matthew R. Hepworth. Antigen-
presenting ILC3 regulate T cell-dependent IgA responses to colonic mucosal
bacteria. Journal of Experimental Medicine. 2019;216(4):728-42.
III. Arlisa Alisjahbana, Yu Gao, Natalie Sleiers, Elza Evren, Demi Brownlie,
Andreas von Kries, Carl Jorns, Nicole Marquart, Jakob Michaëlsson, and Tim
Willinger. CD5 surface expression marks intravascular human innate lymphoid
cells that have a distinct ontogeny and migrate to the lung. Frontiers in
Immunology. 2021; 12:752104.CONTENTS
1 INTRODUCTION.................................................................................................. 1
1.1 MACROPHAGES ........................................................................................ 1
1.1.1 Alveolar macrophages ........................................................................ 1
1.1.2 Interstitial macrophages...................................................................... 3
1.1.3 Intravascular macrophages ................................................................. 5
1.2 MONOCYTES.............................................................................................. 7
1.2.1 Adult monocytes ................................................................................ 7
1.2.2 Fetal monocytes ................................................................................. 8
1.3 ORIGIN AND DEVELOPMENT OF LUNG MACROPHAGES ..................10
1.3.1 Steady state .......................................................................................10
1.3.2 Injury................................................................................................11
1.4 LUNG MACROPHAGE FUNCTIONS ........................................................13
1.4.1 Tissue homeostasis and wound repair ................................................13
1.4.2 Phagocytosis and innate cytokine responses to pathogens ...................13
1.4.3 Immune regulation and crosstalk .......................................................14
1.4.4 Trained immunity .............................................................................15
2 RESEARCH AIMS ...............................................................................................17
3 MATERIAL AND METHODS .............................................................................19
3.1 MISTRG HUMANIZED MICE ...................................................................19
3.2 HUMAN SAMPLES ....................................................................................22
3.3 FLOW CYTOMETRY AND CELL SORTING ............................................22
3.3.1 Principle ...........................................................................................22
3.3.2 Analysis of human myeloid cells by flow cytometry ..........................22
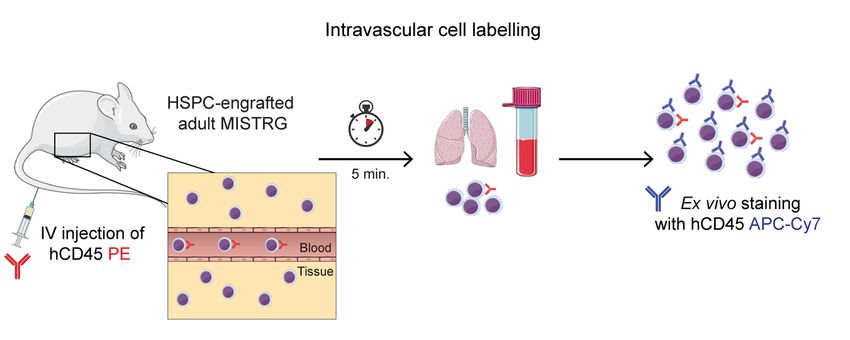
3.4 INTRAVASCULAR CELL LABELLING ...................................................23
3.5 BEAD-BASED FATE-MAPPING OF CIRCULATING PHAGOCYTES .....25
3.6 GENE EXPRESSION ANALYSIS ..............................................................26
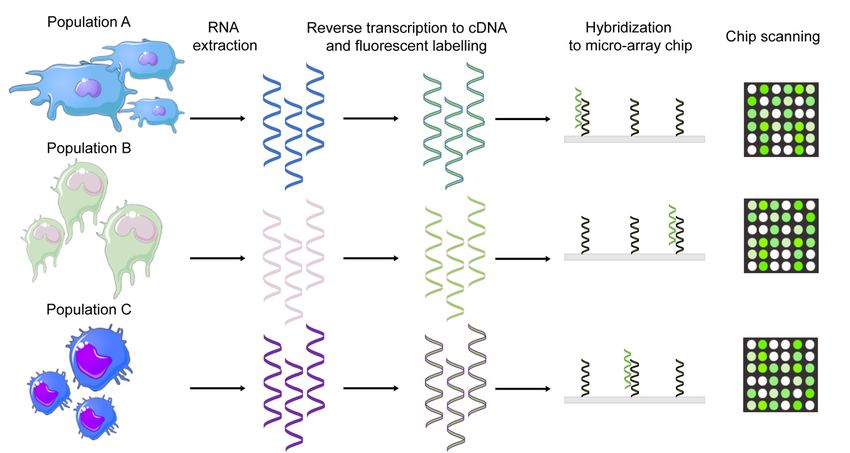
3.6.1 Microarray ........................................................................................26
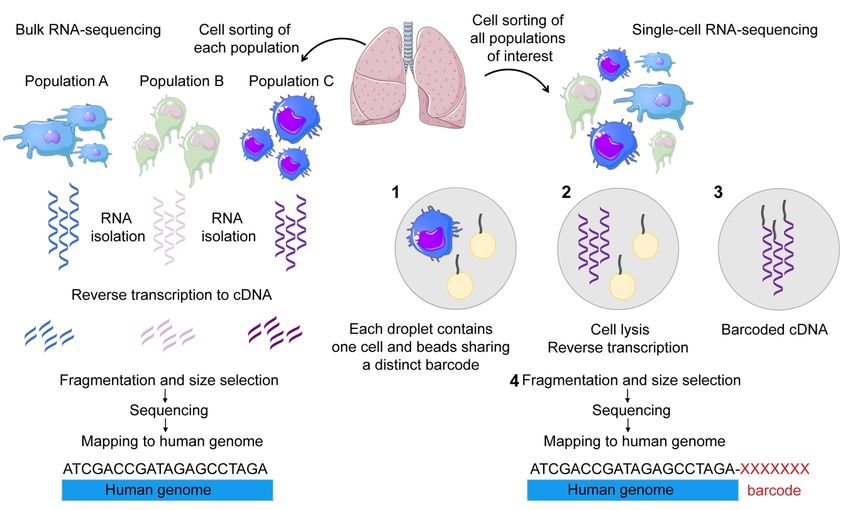
3.6.2 Bulk RNA sequencing.......................................................................26
3.6.3 Single-cell RNA sequencing..............................................................27
3.7 STATISTICAL ANALYSIS ........................................................................28
3.8 ETHICAL CONSIDERATIONS ..................................................................28
3.8.1 Mouse experiments ...........................................................................28
3.8.2 Human tissues...................................................................................29
4 RESULTS AND DISCUSSION ............................................................................31
4.1 ADULT PROGENITORS OF HUMAN LUNG MACROPHAGES ..............31
4.1.1 CD14+CD16- classical monocytes......................................................31
4.1.2 CD14lowCD16+ non-classical monocytes ............................................33
4.2 FETAL PROGENITORS OF HUMAN LUNG MACROPHAGES ...............37
4.2.1 CD116+CD64- fetal precursor-like cells .............................................37
4.2.2 CD116+CD64+ fetal monocytes .........................................................38
4.2.3 Assessing the in-vivo macrophage potential of fetal progenitors..........384.3 IMPACT OF FETAL VERSUS ADULT ORIGIN ON LUNG
MACROPHAGE IDENTITY AND FUNCTION ......................................... 40
4.3.1 Macrophage identity ......................................................................... 40
4.3.2 Function ........................................................................................... 41
5 CONCLUSIONS .................................................................................................. 45
6 POINTS OF PERSPECTIVE ................................................................................ 47
6.1 HUMAN INTERSTITIAL MACROPHAGE HETEROGENEITY AND
ORIGIN ...................................................................................................... 47
6.2 PULMONARY INTRAVASCULAR MACROPHAGE FUNCTIONS IN
HUMANS ................................................................................................... 47
6.3 FETAL PROGENITOR MIGRATION FROM THE LIVER TO THE
LUNGS ....................................................................................................... 48
6.4 ORIGIN OF LUNG MACROPHAGES IN DISEASE CONTEXT ............... 48
7 ACKNOWLEDGEMENTS .................................................................................. 51
8 REFERENCES ..................................................................................................... 55LIST OF ABBREVIATIONS AM Alveolar macrophage AT2 Alveolar type 2 BAL Broncho-alveolar lavage BCG Bacillus Calmette-Guérin BrdU Bromodeoxyuridine cMoP Common monocyte progenitor COPD Chronic obstructive pulmonary disease DEG Differentially expressed gene DNA Deoxyribonucleic acid EGR2 Early growth response 2 EMP Erythromyeloid progenitor FACS Fluorescence activated cell sorting GM-CSF Granulocyte-macrophage colony-stimulating factor GMP Granulocyte-monocyte progenitor HSPC Hematopoietic stem and progenitor cell IHC Immunohistochemistry IL Interleukin IM Interstitial macrophage IV Intravenous LFA-1 Lymphocyte function-associated antigen 1 LPS Lipopolysaccharide LYVE-1 Lymphatic vessel endothelial hyaluronan receptor 1 MAF Musculoaponeurotic fibrosarcoma oncogene homolog MARCO Macrophage receptor with collagenous structure M-CSF Macrophage colony-stimulating factor MDP Monocyte-dendritic cell progenitor MHC Major histocompatibility complex PAP Pulmonary alveolar proteinosis PIM Pulmonary intravascular macrophage PLVAP Plasmalemma vesicle associated protein
PolyI:C Polyinosinic:polycytidylic acid PPARg Peroxisome proliferator-activated receptor gamma RBC Red blood cell RNA Ribonucleic acid SIRPα Signal-regulatory protein alpha TGFb Transforming growth factor beta TPO Thrombopoietin
1 INTRODUCTION
1.1 MACROPHAGES
The Nobel Prize in Medicine of 1908 was awarded to two scientists who defended two distinct
(and yet complementary) concepts of the immune system. On one side stood Paul Ehrlich,
discoverer and adamant supporter of humoral (antibody-dependent) immunity. Opposite to him
was Elie Metchnikoff, professor of zoology from Odessa and vigorous proponent of cellular
(innate) immunity.
Inspired by Darwin’s travels, Metchnikoff was studying invertebrate marine organisms and
their development in the Mediterranean and the Baltic seas [1]. He noticed in particular some
motile amoeboid cells that were present in many tissues. According to his own memoirs, he
was travelling in Messina, Sicily when he discovered a new phenomenon: He introduced a rose
thorn under the skin of a starfish larva and observed the recruitment of the amoeboid cells to
the site of injury. These cells would later be called macrophages (from Greek: “large eaters”)
and the process they form to defend the host from injury named “phagocytosis”.
Macrophages are mononuclear leukocytes found in the tissues where their main duty is to
engulf and digest – phagocytose – pathogens. Furthermore, they are involved in tissue repair
and homeostasis. They come in different flavors: they perform unique tissue specific functions
according to the organ they occupy. Some sites where their involvement is crucial are the heart
where cardiac macrophages have been shown to facilitate electrical conduction [2] and the
brain populated by microglia that ensures its normal development by synaptic pruning [3]. We
chose to study yet another vital organ, the lungs.
The lungs can be divided in three main anatomical compartments: the airways, the lung tissue
and vasculature. Each compartment harbors specific types of macrophages.
1.1.1 Alveolar macrophages
We all start our lives with a first cry. Our first breath triggers the inflation of our lungs and is
instantly followed by exposure to airborne particles and pathogens. Our lungs will keep
ventilating on average 9,000 liters of air every day for the rest of our lives. One can then
imagine how many microbial intruders attempting to find their way into our body are stopped
at the gates.
As their name suggests, alveolar macrophages (AMs) reside in the alveoli where the gas
exchange takes place. AMs have two main roles in the airways. They ensure barrier immunity
and maintain the vital uptake of oxygen by phagocytosing microbes, dead cells, and debris. In
addition, they have an organ-specific function that is to catabolize the surfactant, a thin layer
of mucus produced by the pulmonary epithelium in order to lubricate the surface of the organ
and avoid the friction due to the expansion and contraction of the lungs [4, 5]. AMs have
historically been easier to study as they can be harvested by broncho-alveolar lavage (BAL) on
patients (or mice).
1However, it remains unclear whether this technique allows the analysis of “sessile” macrophages as they are harder to detach from the epithelium which they are in close contact with. The epithelium harbors alveolar type 2 cells (AT2), which on one hand produce surfactant while on the other hand are a crucial player in the maturation and self-renewal of AMs. In mice, epithelial AT2s are the critical source of the cytokine granulocyte-macrophage colony- stimulating factor (GM-CSF), encoded by the Csf2 gene, for AM development [6] .The binding of GM-CSF to its receptor on the surface of AMs elicits the induction of the master transcription factor peroxisome proliferator-activated receptor gamma (PPARg) [7]. Figure 1. Alveolar macrophages (AMs) reside in the airways where they catabolize the surfactant produced by alveolar type 2 (AT2) cells. An important role of AT2 cells is to secrete GM-CSF which is necessary for AM development and survival. AM recognize GM-CSF via the receptor CD116. TGF-b is produced by AMs and act in an autocrine manner. Figure designed with Servier Medical Art. PPARg regulates lipid metabolism and lack of PPARg (due to loss of GM-CSF signaling) leads to the accumulation of lipids and proteins in the airways. This condition is called pulmonary alveolar proteinosis (PAP) and can be observed in Csf2-/- mice as well as patients with mutations in CSF2RA and CSF2RB genes encoding for the a (CD116) and the b (CD131) subunits of GM-CSF receptor, respectively [8]. Moreover, PPARg triggers the activity of the transcription factor early growth response 2 (EGR2) which is required for AM identity and function. Interestingly, EGR2 is dispensable for the lipid handling metabolism of AMs but necessary for host defense functions and generation of monocyte-derived AMs [9]. 2
An important characteristic of AMs is their longevity. Macrophage colony-stimulating factor
(M-CSF) signals through its receptor CD115 at the surface of macrophages and activates
transcription factors such as c-MYC, KLF2 and KLF4 involved in self-renewal. In contrast,
musculoaponeurotic fibrosarcoma oncogene homolog (MAF) transcription factors (c-MAF,
MAFB) suppress macrophage self-renewal by repressing c-MYC [10-12]. Recent studies in
mice have identified the transcription factors Bhlhe40 and Bhlhe41 as indispensable regulators
of self-renewal and proliferation in AMs by potentially inhibiting Maf and Mafb [13].
AMs have first been described as large (larger than 20 µm in humans) and highly auto-
fluorescent macrophages. Flow cytometry revealed that human AMs are CD14lowCD16+ cells
that express common myeloid lineage markers such as HLA-DR, CD11b, CD11c and CD64
and several other specific markers such as the mannose receptor CD206, the sialoadhesin
CD169 [14, 15] and the scavenger macrophage receptor with collagenous structure MARCO
[16]. AMs are sentinels of lung homeostasis: they secrete transforming growth factor beta
(TGFb), a cytokine that induces the regeneration of the injured lung by promoting immune
tolerance [17]. Moreover, this cytokine acts in an autocrine manner and is necessary for the
development and homeostasis of AMs [18].
As some of the characteristics mentioned above indicate that AMs display an anti-
inflammatory phenotype, it is tempting to classify them as “M2 macrophages”. However, the
binary M1/M2 classification is outdated as it does not capture the complexity of macrophage
heterogeneity in response to cues from the tissue environment in vivo.
Technological advances, such as the advent of single-cell RNA sequencing, have allowed to
investigate cellular heterogeneity with a much greater resolution. Recent single-cell RNA
sequencing of BAL samples from healthy donors and cystic fibrosis patients revealed the
presence of four AM superclusters that could be defined by the expression of IFI27 and APOC2
[19]. The authors also highlighted a cluster of chemokine-expressing AMs that was composed
of four separate clusters according to the family of upregulated chemokines. Each cluster of
AMs seems to be specialized in attracting a certain type of cell, which further confirms the
abundance of different AM phenotypes in the airways.
1.1.2 Interstitial macrophages
For decades, interstitial lung macrophages (IMs) have been living in the shadow of their
alveolar counterparts. The first reason behind this neglect is their anatomical localization: They
reside in the lung tissue and thus cannot be washed out by BAL, which makes them harder to
study than AMs. A second probable explanation is the prevailing confusion between tissue
infiltrating lung monocytes and bona fide IMs.
Human IMs are less granular and smaller than AMs and they have been described by flow
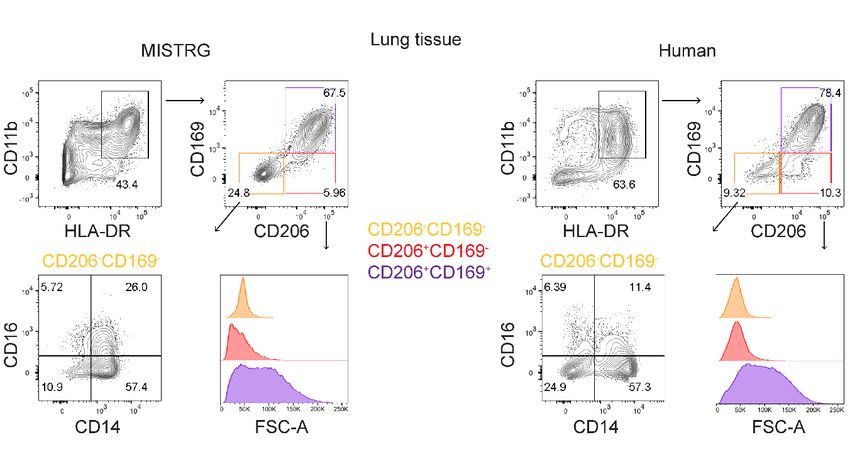
cytometry as CD206+CD169lowCD14+CD16+ [14, 15] or CD64+CD14+CD16-CD206+HLA-
DRhiCD169+ cells [20]. They have been previously observed in several different lung
3compartments such as the alveolar interstitium, the submucosa or the perivascular adventitia [21]. While their precise localization remains controversial, we can assume that they interact with other immune cells such as lymphocytes or dendritic cells in the lung interstitium. In mice, IMs have been observed in the bronchial interstitium, but they were absent from the alveolar interstitium [22]. Mouse IMs can be divided in three subtypes according to their expression of MHC (Major histocompatibility complex) Class II and the integrin CD11c. Interestingly, two of the three reported mouse IM populations expressed the mannose receptor CD206 and the sialoadhesin CD169 while the third one had a CD206lowCD169- phenotype. This third IM population had a transcriptome that was similar to human CD1c+ IMs, with high expression of inflammatory monocyte genes such as IL1B, C1QB, C1QC and F13A1 [23]. Conversely, CD1c+ IMs were mostly similar to mouse splenic CD4+ dendritic cells, hinting towards a potential dendritic cell programming in this subset. A second study in mice revealed that lung IMs occupied distinct niches and functions [20]. LYVE-1lowMHC Class IIhigh IMs were preferentially associated with nerve endings and specialized in antigen presentation. LYVE-1high MHC Class IIlow IMs were strategically located around the lung vasculature where they carried out tissue remodeling and wound healing activities. Consistent with its perivascular localization, this second IM subset has been shown to restrain lung inflammation and fibrosis by controlling the permeability of the blood vessels and thereby the immune cell infiltration into the tissue. Figure 2. Interstitial macrophages (IMs) are a heteregenous population residing in the lung tissue. Cellular markers such as CD11c, MHC Class II and LYVE-1 have been used in previous studies in order to define different IM subsets [20, 22]. LYVE-1high MHC Class IIlow IMs had a perivascular localization while LYVE- 1low MHC Class IIhigh IMs with a higher antigen presenting function preferentially associated with nerve endings [20]. Figure designed with Servier Medical Art. 4
Furthermore, IMs are the main interleukin 10 (IL-10) producer among mouse lung myeloid
cells [24]. IL-10 is an immunosuppressive cytokine expressed highly by IMs in response to the
bacterial endotoxin lipopolysaccharide (LPS) as well as Deoxyribonucleic acid containing
unmethylated CpG motifs (CpG-DNA) both in mice and humans. This feature of IMs suggests
that they are involved in a immunoregulatory loop in the immune response to bacterial
infection.
IMs are multifaceted: They can either display anti or pro-inflammatory properties according to
the physiological condition. In chronic obstructive pulmonary disease (COPD) patients, IMs
express CD36, involved in the efferocytosis of neutrophils, and CD38, associated with
increased macrophage functions such chemotaxis and cytokine production [25]. In addition,
COPD-specific IMs had higher expression of pro-inflammatory genes such as CCL20 and
CXCL1. Interestingly, RNA sequencing of mouse lung IMs following intraperitoneal LPS
administration revealed an upregulation of genes encoding both pro-inflammatory (Il1b, Ccl5,
Cxcl10) and anti-inflammatory (Il10) cytokines, confirming the complex responses of IMs to
inflammation [26].
1.1.3 Intravascular macrophages
A third type of macrophage has been observed in the lung vasculature of several mammals
such as sheep, goats, horses, and cats [27, 28]. These pulmonary intravascular macrophages
(PIMs) have a similar size to their alveolar counterparts (between 20-80 µm depending on the
species) and are tightly attached to the capillary endothelium. Their role, comparable to Kupffer
cells in the liver or red pulp macrophages in the spleen, is to phagocytose bloodborne particles
and damaged or aged red blood cells (RBCs). They are coated by a thick glycocalyx composed
of lipids and lipoprotein globules and contain iron droplets (siderosomes) resulting from the
phagocytosis of erythrocytic debris. In sheep, 20% of the capillary endothelium is covered by
PIMs and their phagocytosis of bloodborne particles occurs faster than the uptake of airborne
particles by AMs [29]. PIMs seem to be adult monocyte-derived in pigs as they were not
observed in newborns but appeared one week or later in older animals [30].
Intravascular macrophages express Toll-like receptors such as TLR4 and TLR9 which when
stimulated, will trigger the production of TNF-a and IL1-b [29].
The presence of PIMs in humans has been controversial. As the alveolar barrier is very thin, it
is worth considering that what has been described as an IM or AM could be as well localized
in the pulmonary capillaries. Another interesting aspect is that PIMs can be induced in bile-
duct ligated rats and humans suffering from liver dysfunction (hepato-pulmonary syndrome)
and sepsis. This observation is consistent with a model in which Kupffer cells are the main
actors in removing bloodborne debris from the circulation, but in case of disease and systemic
physiological stress PIMs can be induced as reinforcements and ensure the same functions.
5Figure 3. Three different type of lung macrophages occupy three respective compartments. Alveolar macrophages (AMs) are found in the airways, interstitial macrophages (IMs) in the interstitium and pulmonary intravascular macrophages (PIMs) in the lung capillaries. Figure designed with Servier Medical Art. 6
1.2 MONOCYTES
1.2.1 Adult monocytes
Van Furth and Cohn described monocytes as “free” mononuclear phagocytes in opposition to
their “fixed” counterparts found in tissues, the macrophages [31]. Considered for decades as
the sole circulating progenitor of tissue macrophages, it has only recently been shown that adult
monocytes can reside in extravascular reservoirs without differentiating into macrophages [32].
In addition, extravascular monocytes can also ensure antigen-presenting functions in the lymph
nodes.
Human and mouse blood monocytes both derive from common monocyte precursors (cMoP)
in the bone-marrow [33]. Human cMoPs are defined as a subset of conventional granulocyte-
monocyte progenitors (GMPs) expressing the C-type lectin CLEC12A and the Fcg-receptor I
CD64 [34]. In mice, GMPs differentiate into bipotent monocyte-macrophage/dendritic cell
progenitors (MDPs) which either give rise to classical dendritic cells or cMoPs [35, 36]. Two
main subsets of monocytes exist during adult life in mice and humans. They are related to each
other by a progenitor-product relationship and have distinct localizations and functions.
In mice, blood monocytes are divided into classical Ly6high and non-classical Ly6low subsets,
the first one giving rise to the second in the circulation [37]. Their egress from the bone marrow
into the blood circulation is dependent on the chemokine receptor CCR2 [38]. Ccr2-/- mice
display reduced numbers of Ly6high monocytes in the blood, tissues and lymph nodes as these
cells stay trapped in the bone marrow [39]. Interestingly, the lifespan of non-classical Ly6low
monocytes is significantly increased in these mice due to the absence of Ly6high monocytes
[37]. Furthermore, Nr4a1-/- mice are characterized by a drastic reduction of Ly6low monocytes,
highlighting the major role of the transcription factor NRA41 in the survival of this patrolling
monocyte subset [40].
In humans, the expression of the LPS-receptor CD14 and the Fcg-receptor III CD16 allows us
to define three monocyte subsets: CD14+CD16- classical, CD14lowCD16+ non-classical and
CD14+CD16+ intermediate monocytes [41, 42]. The latter has been previously described as a
transitional population between classical and non-classical monocytes. However, single-cell
RNA sequencing studies have revealed that this subset is highly heterogenous and composed
of more than one particular type of cell [43].
CD14+CD16- classical monocytes are considered as the human equivalent of mouse Ly6Chigh
monocytes [44]. They can migrate to the organs and show effector inflammatory functions,
whereas CD14lowCD16+ non-classical monocytes are vascular housekeepers that patrol the
blood vessels and support endothelial function. After egressing from the bone marrow, classical
monocytes have a short lifespan and will either give rise to the intermediate subset or migrate
into tissues [45]. The intermediate monocytes will in turn downmodulate CD14 expression and
convert into CD14lowCD16+ non-classical monocytes.
7The effective patrolling of non-classical monocytes is mediated through their expression of the integrin Lymphocyte function-associated antigen 1 (LFA-1) [46]. In addition, they have a higher expression of the fractalkine receptor CX3CR1 [47] compared to the classical monocytes, which fits with their function as fractalkine is a chemokine secreted by the vascular endothelium. Similar to PIMs, non-classical monocytes have been shown to produce TNF-a and IL1-b. However, these cytokines are secreted in response to stimulation of TLR7/8 receptors on non-classical monocytes by nucleic acids and viruses [48]. This cytokine production induces the recruitment of neutrophils to the site of infection and the subsequent necrosis of infected endothelial cells, with the remaining cellular debris being phagocytosed by non-classical monocytes [49]. 1.2.2 Fetal monocytes Primitive macrophage progenitors appear in the yolk-sac, an extra-embryonic structure that provides the embryo with nutrients, around embryonic day E7.0 in mice and week 3 of gestation in humans [50, 51]. This primitive differentiation program consists of early erythro- myeloid progenitors (EMP) that directly differentiate into yolk-sac macrophages without transiting through a monocyte intermediate. A second wave of progenitors called late EMPs appears at the site of the hemogenic endothelium, in the vasculature of the yolk-sac, at E.8.25. Upon the development of the blood circulation from E8.5 onward, late yolk-sac EMPs will seed the fetal liver and give rise to fetal monocytes after E12.5 which are released into blood at E13.5 [52, 53]. First described by Naito et al. in 1990, fetal monocytes are now known to colonize all embryonic tissues after E14.5 and generate tissue-resident macrophages. In contrast to adult hematopoiesis in the bone marrow, the Ly6Clow subset in the mouse fetal liver corresponds to a transient myeloid progenitor for Ly6Chigh fetal monocytes [53]. Similarly, human CD14+ fetal monocyte cells can derive from CD14lowCD16+ progenitors [54]. Fetal and adult monocytes display a comparable responsiveness to GM-CSF [55]. Furthermore, they both express CCR2, even though the egress of fetal monocytes from the fetal liver and migration to the tissues are independent from this expression [53]. Instead, mouse fetal monocytes rely on plasmalemma vesicle associated protein (PLVAP) present on the sinusoidal endothelial cells to exit the fetal liver [56]. Additional differences with adult monocytes include the lack of dependence on M-CSF for the differentiation of fetal monocytes [57, 58], their reduced capacity at recognizing and presenting antigens, as well their highly proliferative phenotype [55]. The transcription factor c-Myb is a crucial player in the differentiation of late EMPs into fetal monocytes. Moreover, late EMPs seem to be dependent on c-Myb expression for their survival and expansion [53]. In contrast, yolk-sac macrophages develop from early EMPs in an Myb- 8
independent manner, as illustrated by embryos from Myb-/- mice having less late EMPs and
fetal monocytes while the yolk-sac macrophage numbers remain unaltered [59].
Figure 4. Adult CD14+CD16- classical monocytes (the equivalent of mouse Ly6Chigh monocytes) develop in
the bone-marrow from common monocyte precursors (cMoP) and egress to the blood circulation in a CCR2-
dependent manner. Once in the blood, they either extravasate into tissues or differentiate into CD14+CD16+
intermediate and CD14lowCD16+ non-classical monocytes. Fetal monocytes emerge from c-Myb expressing
EMPs and their exit from the fetal liver is depending on PLVAP in mice. Figure designed with Servier Medical
Art.
91.3 ORIGIN AND DEVELOPMENT OF LUNG MACROPHAGES The debate around the origin of macrophages has been rekindled by seminal discoveries in the past decade. In 1968, Van Furth and Cohn proposed the mononuclear phagocyte system concept with monocytes being the circulating progenitors of resident tissue macrophages. By contrast, pioneering work by Ginhoux and colleagues have demonstrated that most tissue macrophages in mice derive from fetal monocytes that seed the organs before birth. Elie Metchnikoff and Paul Ehrlich were both right in their vision of the immune system, but they were wrong to consider that their concept was the only right one. Similarly, there is not only one but many complementary origins of lung macrophages that vary according to developmental age, environmental exposure, and location in health and disease. 1.3.1 Steady state Macrophage progenitors seed organs in three consecutive waves. First, primitive macrophages originating from early yolk-sac EMPs colonize every embryonic tissue [53]. In fact, adult microglia in the brain has been shown to derive from yolk-sac macrophages [57]. Primitive macrophages are sequentially replaced by fetal monocytes, which are the predominant macrophage progenitors around the time of birth [53]. For example, Langerhans cells in the epidermis originate from fetal monocytes with a negligible blood monocyte recruitment in steady state [58, 60, 61]. Finally, in some organs such as the gut, dermis and the heart, adult blood monocytes are recruited after birth and differentiate into macrophages that replace their fetal monocyte-derived counterparts [62-64]. In mice, AMs have been shown to arise from fetal monocytes [65]. The development of AMs from fetal monocytes is highly dependent on the expression of the master transcription factor PPARg that is induced by GM-CSF in the fetal lung [7]. Many conclusions regarding the origin of AMs result from experiments and observations in mice as it is extremely challenging to perform cell fate-mapping in humans. Lung transplantation studies in humans have indicated that blood monocytes contribute substantially to the human AM compartment already in steady state [66-70]. Humans are constantly exposed to airborne microbes which could explain that fetal monocyte-derived AMs are slowly depleted and replaced by adult blood monocyte-derived counterparts. It is likely that human AMs are from both embryonic and adult origins. It is hard to mimic the effects of environmental exposure over the lifetime in mice as mouse studies are normally conducted in specific pathogen-free animal facilities over a short time period. Interestingly, experiments using Flt3Cre mice which tags hematopoietic stem cells and their progenitors (including circulating blood monocytes), have shown that the labelling of AMs increased from 16% in 12 weeks-old mice up to 40% at one year of age, indicating a higher contribution of recruited monocyte-derived cells to the lung macrophage pool [71]. Similarly, mouse fate-mapping models based on the expression of Ms4a3 established that blood monocytes increasingly supplied the AM compartment in older mice [72]. In mice, AMs 10
from embryonic and adult origins had a very similar transcriptome, demonstrating the impact
of the lung environment on AM development [73]. However, some genes were found to be
associated with AM origin: Marco was differentially expressed in embryonic-derived AMs
while C1qb and Plbd1 were upregulated in bone marrow-derived AMs.
On the other hand, the absence of a consensus on different IM populations makes it harder to
precisely define their origin. Both LYVE-1lowMHC Class IIhigh and LYVE-1highMHC Class
IIlow IMs have been described to be monocyte-derived but from two separate lineages
originating from Ly6Chigh monocytes [20]. Conversely, CD169+ nerve and airway associated
macrophages – similar to LYVE-1lowMHC Class IIhigh nerve-adjacent macrophages – originate
from yolk-sac progenitors [74].
Gibbings et al. have demonstrated that while all three IMs populations could be reconstituted
by circulating progenitors, CD11c+ MHC Class II+ IMs could be replaced more easily than
their CD11c- MHC Class II+ and CD11c- MHC Class IIlow counterparts [22]. Furthermore,
Ly6Clow patrolling monocytes have been proposed to extravasate to the lung tissue and give
rise to CD206- IMs that specialize in antigen presentation [75]. Likewise, “definitive” mouse
IMs that replace their primitive yolk-sac derived F4/80+ counterparts have been shown to
originate from adult bone-marrow hematopoiesis [76]. Overall, studies in mice converge on a
greater contribution of adult blood monocytes to IMs compared to AMs.
Interestingly, the dichotomy in macrophage origin can be reflected in metabolism: while AMs
use fatty acid oxidation to survive, IMs are dependent on glycolysis similarly to monocytes [7,
77].
1.3.2 Injury
Lung injury is defined by the disruption of the lung tissue and the capillary-alveolar interface.
It can occur due to harmful micro-organisms penetrating the lungs or sterile inflammation
induced by necrotic cells and environmental chemicals such as silica or asbestos particles [78].
Severe lung injury can result in the death and depletion of tissue-resident macrophages, which
triggers the recruitment of circulating blood monocytes into the lung.
An actual and relevant example is SARS-CoV-2 infection which leads to the accumulation of
pro-inflammatory monocyte-derived macrophages in the BAL fluid. These monocyte-derived
macrophages express genes typical for circulating classical monocyte, such as S100A8, FCN1
and CD14, pointing at their monocytic origin [79]. A complementary study supported the
finding that BAL macrophages in individuals with COVID-19 originated from blood
monocytes [80].
Another viral infection resulting in the recruitment of blood monocytes to the airways is
influenza [81]. Bone-marrow derived progenitors replenish the alveolar macrophage pool in
influenza-infected mice and produce IL-6 that ensures protection against a secondary infection
11by Streptococcus pneumoniae. The authors highlight an important notion in this study: time. Indeed, the protective effect of monocyte-derived AMs slowly decreases over time and their transcriptome resembles the transcriptome of the resident AMs initially present in the lungs. A similar imprinting by the lung environment is observed in fibrosis. The fibrotic phase is characterized by the influx of blood monocytes which differentiate into AMs and drive the disease. The difference observed in the transcriptome of tissue-resident versus recruited AMs is however dramatically reduced one year after the resolution of fibrosis [82]. The monocytic progenitor of lung macrophages is not always a circulating one. In fact, stimulation by the TLR9 agonist CpG induces not only the differentiation of resident lung monocytes to IMs but also the mobilization of splenic monocytes to the lung to give rise to IMs [83]. An important question in the field arises from the relationship between macrophage origin and function, however the distinct adult and fetal progenitors of human lung macrophages remain unknown. Figure 5. Alveolar macrophages (AMs) have different origins according to the stage of life. The embryonic lungs are first colonized by yolk-sac macrophages that are replaced by fetal monocyte-derived AMs around the time of birth in mice. The precise identity of the human fetal AM progenitor remains unknown. Embryonic derived AMs are depleted with ageing, environmental stress and lung injury and adult blood monocytes are recruited from the blood circulation to give rise to monocyte-derived AMs. Thus, AMs found in adults have a dual origin. Figure designed with Servier Medical Art. 12
1.4 LUNG MACROPHAGE FUNCTIONS
Lung macrophages can accomplish various functions according to cues from the environment
and interactions with potential invaders. Their high plasticity supports their role as gatekeepers
in the lungs. They participate in the homeostasis of the lung tissue, eliminate intruders and, if
needed, induce an inflammatory reaction to call for reinforcements.
1.4.1 Tissue homeostasis and wound repair
In addition to the main homeostatic duty of metabolizing pulmonary surfactant, AMs
potentially contribute to the alveolar integrity by releasing TGF-b which orchestrates the ion
and fluid transport at epithelial sites [84]. Upon tissue injury, AMs also contribute indirectly to
TGF-b production by releasing amphiregulin which triggers a cascade of TGF-b activation that
induces the differentiation of pericytes into collagen-secreting myofibroblasts [17].
Furthermore, AMs produce fibronectin [85], which has been shown to promote epithelial cell
proliferation [86].
On the other hand, IMs secrete platelet-derived growth factor (PDGF) which likely stimulates
epithelial and fibroblast growth [87]. Similarly, the depletion of perivascular LYVE-1highMHC
Class IIlow IMs causes increased infiltration of immune cells into the lung tissue, suggesting
that these IMs control the permeability of lung capillaries [20].
1.4.2 Phagocytosis and innate cytokine responses to pathogens
Phagocytosis is a biological process by which cells can ingest particles (bacteria, dead cells)
with a diameter greater than 0.5µm [88]. It is the primary cellular function that defines
macrophages across tissues and species. The particle to engulf is either directly recognized by
the phagocytic cell via surface receptors such as the mannose receptor CD206 [89], the LPS
receptor CD14 [90], the scavenger receptor MARCO [91] or it is coated with opsonins which
bind to the phagocytic cell and induce phagocytosis [88].
The main types of opsonins are IgG antibodies and complement proteins. Antibodies attach to
the Fcg receptors on the phagocytic cell which results in the ingestion of the antibody-particle
complex. This internalization is mediated by a fast remodeling of the actin cytoskeleton and
lipids forming the extracellular membrane and leads to the extension of the membrane to engulf
the particle or antibody-particle complex. The folding of the extracellular membrane over the
particle forms the phagosome that progresses inside the cytoplasm to fuse with parts of the
lysosome and create the phagolysosome where the particles are destroyed.
AMs and IMs eliminate airborne pathogens by phagocytosis and form the first and second lines
of immune defense in the lungs, respectively. AMs phagocytose bacteria and consequently die
by apoptosis to clear the airways [92, 93]. Antibodies elicited following influenza infection or
vaccination trigger phagocytosis of opsonized pathogens and inflammatory cytokine release
13Vous pouvez aussi lire

















































