Passer du génotype au phénotype - Expliquer le polymorphisme des gènes qui contrôlent les caractères quantitatifs Prédire l'évolution : des ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Passer du génotype au phénotype • Expliquer le polymorphisme des gènes qui contrôlent les caractères quantitatifs • Prédire l’évolution : - des caractères quantitatifs - des fréquences des allèles

Génétique et évolution des flux
métaboliques
Christine Dillmann, Julie Fievet, Bruno Bost1, Frédéric Gabriel,
Sébastien Lion, Luc Négroni, Gilles Curien2, Jean Labarre3,
Delphine Sicard, Dominique de Vienne
1 IGM, Orsay; 2 CEA, Grenoble; 3 CEA, Saclay
UMR de Génétique Végétale, INRA/UPS/CNRS/INA PG
Ferme du Moulon, 91190 Gif-sur-Yvette, France
dillmann@moulon.inra.fr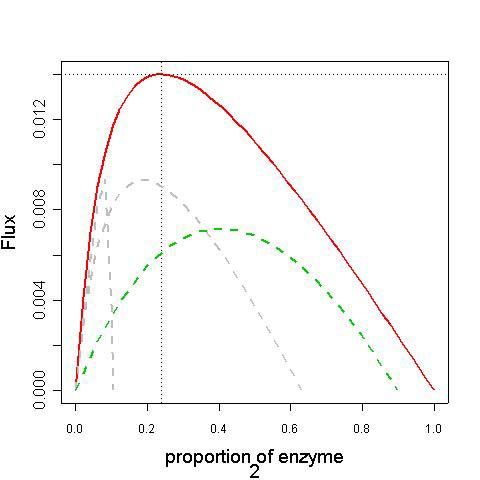
La cinétique enzymatique : un peu d’histoire
• 1903 : Victor Henri : la formation du complexe
enzyme-substrat est l’étape essentielle du
mécanisme catalytique
• 1913 : Michaelis & Menten proposent les
équations de vitesse
• 1973-1974 : Kacser & Burns et Heinrich et
Rapoport proposent la théorie du contrôle
métabolique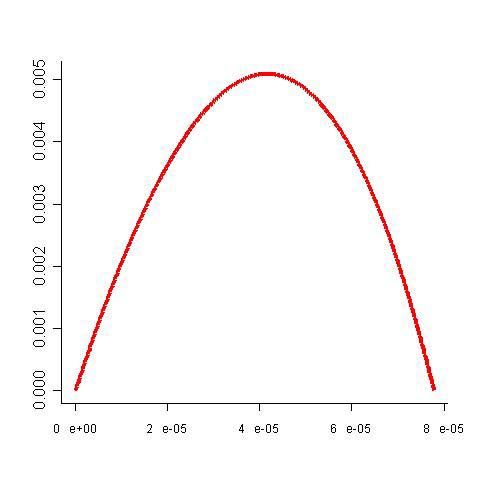
Equation de vitesse de Michaelis-Menten
k+1 kcat
S + E ES P
k–1
Vmax S v vitesse de la réaction
v = Vmax vitesse maximum
S concentration en substrat
Km + S Km = (k–1 + kcat) / k+1,
constante de Michaelis-Menten
Equation de vitesse : cas général
E
Si Sj
Vmax/Km Keq = [Sj]/[Si]
« efficacité constante d’équilibre
physiologique »
(V maxi / Km i) (Si − S j / Keq)
v=
1+ Si / Kmi + S j / Km j
La genèse de la théorie du contrôle
métabolique Kacser 1957
« The problem is therefore the investigation of
systems, i.e. components related or
organised in a specific way. The properties
of a system are in fact « more » than (or
different from) the sum of the properties of
its components »
Travail à la chaîne…
Formalisation de la théorie du contrôle métabolique
E1 E2 Ej Ej+1 En-1 En
S0 S1 … Sj … Sn-1 Sn
Chaîne linéaire d’enzymes michaéliennes indépendantes
à l’état stationnaire : v1 = v2 = … = vj = … = vn = J
Les enzymes fonctionnent loin de la saturation
(V max i / Km i ) ( Si − Si +1 / K eq )
vi = ≈ (V max i / Km i ) ( Si − Si +1 / K eq )
1 + Si / Km i + Si +1 / Kmi +1
Kacser and Burns, 1973
Heinrich and Rappoport, 1974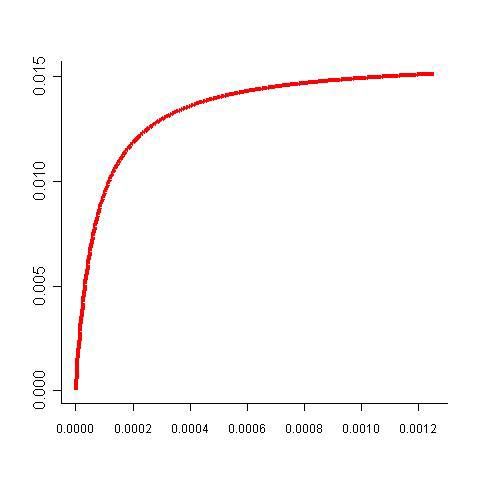
Formalisation de la théorie du contrôle métabolique
E1 E2 Ej Ej+1 En-1 En
S0 S1 … Sj … Sn-1 Sn
à l’état stationnaire : v1 = v2 = … = vj = … = vn = J
S
S0 − n
K
J= 0,n
1
∑ j =1 E
n
j
Kacser and Burns, 1973
Heinrich and Rappoport, 1974
Formalisation de la théorie du contrôle métabolique
Efficacité de l’enzyme :
Vmax j
Ej = K 0, j +1 = Aj Qj
KM j
Paramètres cinétiques de la réaction catalysée :
kcat j
Aj = K 0, j +1
KM j
Concentration cellulaire de l’enzyme
QjLa théorie du contrôle métabolique :
Relation entre enzymes et flux : enzymes indépendantes
Jmax
mes
nz y
Flux J ne
e
z ym
1 en
Qj ou Aj ou Ej = Qj x Aj
• Le flux varie de façon non linéaire avec la concentration d’une enzyme de la
chaîne
• Il tend asymptotiquement vers un maximum qui dépend des paramètres de
toutes les enzymesLa théorie du contrôle métabolique :
Le contrôle du flux : enzymes indépendantes
Jmax
∂J Ei
C =J
Ei
∂Ei J
Flux J
1
Ei
= n
∑
j =1
1
Ej
Qj ou Aj ou Ej = Qj x Aj
• Le coefficient de contrôle mesure la variation relative du flux en réponse à
une variation relative d’efficatité enzymatique.
• Le contrôle qu’une enzyme exerce sur la chaîne dépend de l’ensemble des
enzymes de la chaîne.La théorie du contrôle métabolique :
Le contrôle du flux : propriété de sommation
0≤C J
Ej ≤1
∑
n
j =1
C J
Ej =1
• Le contrôle est partagé entre toutes les enzymes de la chaîne.
• La propriété de sommation est vraie également pour des systèmes
branchés.Validation expérimentale des prédictions de la
théorie du contrôle métabolique : in vivo
Kacser and Burns, 1981. Genetics 97:639Validation expérimentale des prédictions de la TCM :
reconstruction d’une chaîne métabolique in vitro
Ces résultats ne sont pas encore publiés. Nous les diffuseront lorsque le manuscript sera so
Julie Fievet et Gilles CurienLa théorie du contrôle métabolique en génétique et
évolution
Gènes Enzymes Flux Phénotype
Caractère : flux J
Gènes : enzymes allèle caractérisé par
Aj paramètres cinétiques de la réaction cataclysée
Qj concentration cellulaire de l’enzyme
pj fréquence dans une population
Paramètres non génétiques :
S0 concentration en substrat initial
Sn concentration en substrat final
Kj,j+1 constantes d’équilibre de la réactionLa théorie du contrôle métabolique en génétique et
évolution
• Les flux sont reliés au phénotype et à la
valeur sélective de l’individu
• La théorie du contrôle métabolique fournit
une relation simple entre la variation du
phénotype (le flux) et la variation des
paramètres génétiques des enzymes,
activité et concentration.La théorie du contrôle métabolique en génétique et
évolution
E1 E2 Ej Ej+1 En-1 En
S0 S1 … Sj … Sn-1 Sn
à l’état stationnaire : v1 = v2 = … = vj = … = vn = J
Paramètres non génétiques
(environnement) S
S0 − n
K
J= 0,n
1
∑ j =1 A Q
n
j j
Constantes cinétiques de Ej
Concentration cellulaire de EjFlux métaboliques et valeur sélective
Dean et al, 1989
Taux croissance
J
Il existe une relation linéaire entre le flux de lactose et la
vitesse de croissance en chémostat d’Escherichia coliVariabilité génétique des paramètres cinétiques
- Peu de données in vivo
- Modérément variable
Wang & Dykhuisen, 2001. Pathway of gluconate
metabolism in E. coli. Evolution, 55:897.Variabilité génétique des concentrations des enzymes
IPG
SDS
Concentrations des enzymes de la
glycolyse dans deux souches de
levure
fba1
40
qi CENPK
35
qi S288C
30
25
Julie Fiévet 20
15
10
5
0
I
I
H1
A1
PM
K
K1
O
K1
DH
K
PG
TP
PG
PY
EN
FB
HX
PF
AD
AP
G
GVariations génétiques des quantités de protéines chez le maïs
F1254 FxW W117
Additivité : ~ 80 %
Dominance positive ~ 16 %
Dominance négative ~ 4 %La TCM : implications génétiques
Pourquoi les mutations défavorables sont-elles récessives?
Flux
JAA
JAa
Kacser and Burns, 1981.
The molecular basis of dominance
Genetics 97:639
Jaa
Génotype aa Aa AA Qi“ Sélection naturelle de la neutralité sélective ”
Hartl et al., 1985. Limits of adaptation: the evolution of selective neutrality. Genetics 111:655.
J or w
Asymétrie de Sélection Neutralité
l’effet des
mutations
naturelle sélective
Ei= Qi . Ai
−δ +δ
Pb : grandes valeurs de Qi ?
Koehn, 1991. BJLS, 44:231Constraints on enzyme concentration
I – Total enzyme content
1- Space constraints Limited cell/organelle volume ⇒ crowding
E. coli
Goodsell,1992
Warner, 1999, TIBS, 24:437.
- As much as 30–40% of the cytoplasmic volume is occupied by ribosomes.
- The proteins represent 20% to 30% of the cell volume (120 to 300 mg.mL–1).
Beyond: limitation of solubility and diffusion → aggregation or crystallization of proteins,
lower diffusion of important metabolites (e.g. ATP, O2).Constraints on enzyme concentration I – Total enzyme content 2- Energy constraints (ATP/GTP) The cost of maintaining the protein pool depends on Mr, turnover and concentration of the proteins. Proteins are expensive: 12 to 74 ATP/GTP per amino acid, ~ 5 per peptide bond (+ cost of tRNA and mRNA syntheses) → Maintaining protein pool would consume up to 45% of the ATP/GTP!
Natural selection to save ATP/GTP Cost Akashi and Gojobori, 2002. Metabolic efficiency and amino acid composition in the proteomes of Escherichia coli and Bacillus subtilis. PNAS, 99:3695
Constraints on enzyme concentration
I – Total enzyme content
3- Resource constraints
- Internal resources: elements of the transcription and
translation machinery. Ex: ribosomes are limiting for
gene expression in E. coli (Vind et al., 1993, 231:678.)
- External resources (nitrogen, phosphate, etc.) are
never unlimited.Constraints on enzyme concentration
I – Total enzyme content
The cell must limit enzyme content allocated to a given pathway
Proportion of the other enzymes
n
∑Q
i =1
i = Q Tot = Constant
Redistribution
coefficient (slope):
αij = δQj /δQi =- Qj /(QTot- Qi)
Proportion of enzyme k
« Competition » between enzymes: concentrations
are globally negatively correlatedConstraints on enzyme concentration
II – Co-regulated enzymes
PCA on 500 protein volumes along a maize leaf
2
1
Axe 1 : 32,6%
Axe 2 : 24,9%
Delphine Vincent & Michel Zivy, in prep.
Physiological, positive or negative correlationsMetabolic control theory under constraint
Two constraints on enzyme variation are
considered,
Co-regulation and/or competition
Qj
Q1 Independence
Positive
Q2 co-regulation
Negative
Q3 co-regulation,
and/or competition
QiMetabolic control theory under constraint
Proportion of the other enzymes Co-regulation and competition
Redistribution
coefficient (slope):
αij = δQj /δQi
Can be positive or negative
Proportion of enzyme kLa théorie du contrôle métabolique en présence de contraintes
Quelle est la répartition optimale des concentrations des enzymes,
cad la répartition qui maximise le flux ?
⇒ Lagrange’s multiplier
∂ J q*j
Flux
∀ i, =0 ∀j, k, * = Ak
∂ Qi qk Aj
Maximal flux
Heinrich et al., 1991.
Eur. J. Bioch. 201:1.
ot
Q T
Optimal
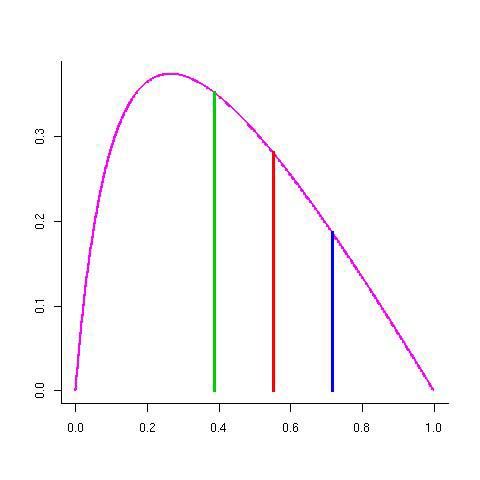
concentration QiLa théorie du contrôle métabolique en présence de contraintes
JOPT
Flux J
Concentration de l’enzyme j
• Il existe une répartition des quantités d’enzymes qui maximise le flux
• L’ensemble des points expérimentaux doivent se trouver sous la courbe rougeCo-régulation et competition
compétition
Proportion des autres enzymes
Proportion enzyme 2
Corégulation + compétition
Proportion des autres enzymes
Proportion enzyme 2
Proportion enzyme 2
•L’ensemble des points expérimentaux doivent se trouver sous la courbe rouge
• A moyens (Qtot) constants, le flux maximal ne peut être atteint en présence de
co-régulationsCo-régulation et competition : coefficient de réponse
combiné
∂J/J J α ij
qi ∑ j =1
n
R =
J
=
∂ Qi / Qi XQTot
Qi
A j q 2j
Coefficient de réponse combinée
Enzyme trop peu abondante
Enzyme en excès
− ∞ ≤ R ≤ +∞
J
Qj
Proportion de l’enzyme iGabriel, Dillmann,
Implications sélectives de la contrainte :
Fievet, Bost, de
assymétrie des coefficients de sélection Vienne, in prep
Flux
Jm(+)
Jw
Le coefficient de sélection s
mesure l’avantage relatif des
Jm(-) individus ayant la plus forte valeur
sélective
δ -δ Proportion de l’enzymeGabriel, Dillmann,
Implications sélectives de la contrainte :
Fievet, Bost, de
assymétrie des coefficients de sélection Vienne, in prep
Flux δ R qw
J
Jm(+) s+ =
1 + δ R qw
J
Jw
s − = − δ R qw
J
Jm(-)
La sélection est plus efficace
pour éliminer une mutation
défavorable que pour fixer une
mutation favorable
δ -δ Proportion de l’enzymeImplications sélectives de la contrainte
contre-sélection des concentrations extrêmes
Flux
Jm(-x)
Jm(+x)
La sélection est plus efficace pour diminuer
la quantité d’une enzyme en excès que pour
Jw augmenter la quantité d’une enzyme en
déficit
+ 10% - 10%
Proportion de l’enzymeImplications sélectives de la contrainte
Scénarios évolutifs
Suppression des corégulations
corégulation
Proportion de l’enzymeConclusions • La TCM, avec ou sans contraintes, est un modèle robuste pour prédire les variations de flux (phénotype) à partir des variations des concentrations des enzymes (génotype) • Elle permet d’expliquer des phénomènes génétique connus (dominance, hétérosis) • Elle permet de proposer des scénarios évolutifs pour les enzymes et les flux • Les prédictions sont fortement dépendantes de la nature des contraintes sur les concentrations des enzymes
Questions o Quelle est l’étendue de la variation des quantités des enzymes d’une même chaîne métabolique dans les populations naturelles ? o Les enzymes d’une même chaîne métabolique sont-elles co-régulées ?
Christine Dillmann Julie Fiévet Sébastien Lion Frédéric Gabriel Gilles Curien Bruno Bost CNRS, Grenoble
Vous pouvez aussi lire

















































