Développement de dispositifs et de méthodologies pour mesurer des paramètres biophysiques reliés au transport d'eau cellulaire avec la microscopie ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous

Développement de dispositifs et de méthodologies pour
mesurer des paramètres biophysiques reliés au
transport d’eau cellulaire avec la microscopie
holographique numérique
Mémoire
Emile Rioux-Pellerin
Maîtrise en biophotonique - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Emile Rioux-Pellerin, 2021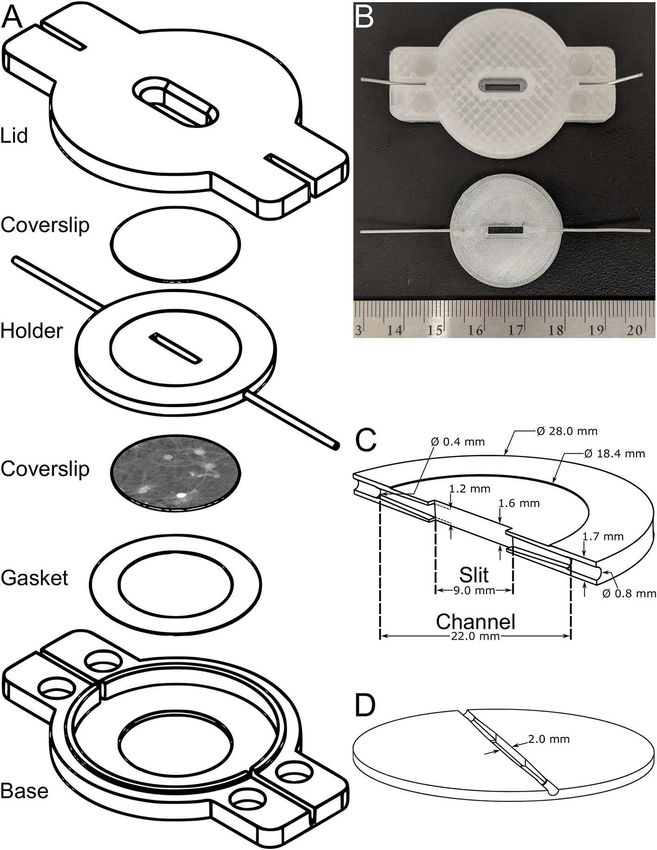
Résumé
La capacité à mesurer le volume cellulaire est essentielle pour l’étude de la régulation du volume
cellulaire et ses nombreux processus. La perméabilité de la membrane des cellules de mammifères
implique que les intrants ou les extrants de biomolécules causent des transports d’eau qui éprouvent
l’homéostasie cellulaire. Dès lors, le volume d’une cellule, ainsi que d’autres paramètres biophysiques
y étant reliés, sont modifiés aussi bien par des changements du milieu extracellulaire que par sa propre
activité. Les cellules ont développé toute une série de mécanismes pour réguler leur volume afin de
préserver leur homéostasie. Ainsi, la régulation de volume d’une cellule est un indicateur potentiel
de son état. Dans le but de produire des mesures fiables de volume lors de l’étude de ces processus,
il est à priori avantageux de développer des outils et des méthodes efficaces. À cet effet, l’imagerie
quantitative de phase avec l’holographie numérique de phase peut donner d’utiles informations sur
la morphologie et la composition de la cellule. Le signal quantitatif de phase d’un objet contient
deux paramètres critiques pour l’étude de la régulation de volume et du transport d’eau : l’indice
de réfraction et l’épaisseur cellulaire. L’ambiguïté de leur contribution dans le signal total peut être
résolue par le découplage à deux liquides, mesurant deux fois le même échantillon dans deux milieux
extracellulaires d’indices de réfraction différents et connus. La procédure sépare les paramètres en un
système facilement soluble de deux équations et de deux inconnus. Avec ces deux paramètres, il est
possible de déduire le volume cellulaire absolu ainsi que d’autres paramètres biophysiques, comme la
masse sèche, la concentration en biomolécule et la fraction d’eau. Pour mieux utiliser l’information
obtenue par le changement de fluide, une chambre d’écoulement imprimable en 3d a été conçue et
utilisée conjointement avec des lamelles rainurées permettant d’obtenir l’indice de réfraction du milieu
d’immersion en temps réel. Cet ajout, avec une compréhension de la mécanique des fluides, ouvre de
nouvelles possibilités pour résoudre des changements de volume dans le temps. Le découplage de
l’indice de réfraction et de l’épaisseur cellulaire n’est désormais plus limité à l’état initial et final
des changements de fluide, mais peut être effectué dans la période transitoire. Ces nouveaux essais
fluidiques sont utilisés pour mesurer les changements de volume des cultures primaires d’astrocytes
de cortex de rat lors de chocs au glutamate.
ii
Abstract
The ability to measure the absolute cell volume is essential to the study of cellular volume regulation
and its many different pathways. The permeability of the cell membrane implies that biomolecule
inputs and outputs cause transmembrane water transport challenging cell homeostasis. Thenceforth, a
cell’s volume and a number of other biophysical parameters are modified by changes to the extracel-
lular medium as well as its own activity. Thus, cells have developed a whole series of mechanisms to
regulate their volume and maintain their homeostasis. Therefore, cell volume regulation is indicative
of the cell’s state. In the interest of producing reliable volume measurements, it is first advantageous
to develop effective tools and methods. For this purpose, quantitative-phase imaging with digital holo-
graphic microscopy can provide useful information on the morphology and composition of the cell.
The quantitative-phase signal of any object contains two critical parameters for the study of volume
regulation, the refractive index and the thickness. The ambiguity of their contribution in the total
quantitative phase signal can be resolved by the two-liquid decoupling approach, measuring twice the
same sample in two extracellular mediums with different but known refractive indices. The procedure
results in two easily solvable phase shift equations with two unknown variables. With the two param-
eters, the whole-cell volume can be deduced in addition to the dry mass, intracellular concentration
and water content fraction. In order to make better use of the information acquired with the change of
fluid, a custom 3d-printed imaging chamber was designed and optimized for stable laminar flow. For
further accuracy, grooves were etched on the sample coverslips to monitor in real-time the extracel-
lular refractive index. The combination of both tools expands the utility of the decoupling procedure.
It is no longer limited to the initial and final state of the liquid exchange, but can now be used in
the transition period. These novel fluidic essays are used to measure glutamate-induced swelling of
primary rat cortical astrocytes.
iii
Table des matières
Résumé ii
Abstract iii
Table des matières iv
Liste des tableaux vi
Liste des figures vii
Remerciements x
Avant-propos xi
Introduction 1
0.1 Importance du volume cellulaire et des mouvements d’eau transmembranaires . . 1
0.2 Microscopie par holographie numérique . . . . . . . . . . . . . . . . . . . . . . 2
0.3 Calculs de paramètres biophysiques . . . . . . . . . . . . . . . . . . . . . . . . 6
0.4 Mécanique des fluides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
0.5 Astrocytes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
1 Mesures de volume de cellules entières avec découplage à deux liquides 10
1.1 Matériels et méthodes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
1.2 Résultats et discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2 Développement des outils et techniques connexes d’imagerie 30
2.1 Chambre d’imagerie à grand champ de vue . . . . . . . . . . . . . . . . . . . . 30
2.2 Conception de masques de photolithographie et production de lamelles rainurées 38
2.3 Calcul de l’indice de réfraction lors de changements de liquides . . . . . . . . . 46
3 Émergence de nouvelles techniques 52
3.1 Utilisation d’un gradient d’indice lors d’un changement de liquide . . . . . . . . 52
3.2 Détermination d’indice de réfraction cellulaire par égalisation d’indice de réfraction 56
4 Réponse astrocytaire aux stimulations glutamatergiques 59
4.1 Réponse de phase des astrocytes lors de stimulations glutamatergiques . . . . . . 59
4.2 Réponse en volume des astrocytes aux stimulations glutamatergiques . . . . . . . 62
Conclusion 69
iv
A Caractérisation supplémentaire avec perfusion d’encre 70
B Détermination de l’indice de réfraction et l’épaisseur cellulaire avec un lissage 72
Bibliographie 75
v
Liste des tableaux
1.1 Paramètres biophysiques mesurés sur des populations de neurones primaires, cellules
HEK293T et NIH3T3 à l’état basal . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
1.2 Variations des paramètres biophysiques induites par des chocs osmotiques . . . . . . 26
2.1 Procédure d’écoulement pour générer un gradient d’indice de réfraction du milieu
avec l’iodixanol . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
vi
Liste des figures
0.1 Géométrie considérée pour la reconstruction d’un hologramme . . . . . . . . . . . . 3
0.2 Schématisation du processus d’imagerie avec la microscopie holographique numérique 4
0.3 Schéma d’une vue de côté d’une cellule dans une chambre d’imagerie et définition de
variables. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
0.4 Schéma du profil de vitesse pour un écoulement laminaire. . . . . . . . . . . . . . . 9
1.1 Séparation de l’écoulement dans une fente d’imagerie rectangulaire à un débit de 2,0
mL/min . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
1.2 Présentation de la chambre d’imagerie imprimée en 3D . . . . . . . . . . . . . . . . 16
1.3 Comparaison des lignes de courant obtenues avec simulation et par µPIV . . . . . . . 18
1.4 Graphique de la longueur laminaire simulée et expérimentale en fonction du débit de
perfusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
1.5 Estimation de la survie cellulaire dans la chambre imprimée en 3D . . . . . . . . . . 20
1.6 Graphique du QPS cellulaire moyen d’une cellule HEK293T durant une procédure de
découplage à deux liquides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
1.7 Incertitudes absolues et relatives des mesures de paramètres biophysiques cellulaires 23
1.8 Distribution des paramètres biophysiques cellulaires . . . . . . . . . . . . . . . . . . 24
1.9 Graphique du QPS moyen d’une cellule HEK293T durant un choc hypo-osmotique . 25
1.10 Découplage pixel par pixel d’une cellule HEK293T . . . . . . . . . . . . . . . . . . 28
2.1 Présentation de la chambre d’imagerie à grand champ de vue . . . . . . . . . . . . . 34
2.2 Comparaison des lignes de courant expérimentales et simulées de la chambre à grand
champ de vue . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
2.3 Graphique de l’aire laminaire normalisée en fonction du débit . . . . . . . . . . . . 37
2.4 Simulations en régime transitoire du transport de solutés dans la chambre à grand
champ de vue . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
2.5 Viabilité de neurones dans la chambre d’imagerie à grand champ de vue et dans une
alternative commerciale. Comparaison avec la viabilité d’une culture cellulaire témoin. 39
2.6 Dessins de CAO des masques par dépôt métallique . . . . . . . . . . . . . . . . . . 41
2.7 Image de phase quantitative d’une lamelle rainurée obtenue avec un masque sur acétate 43
2.8 Image de phase quantitative d’une lamelle rainurée obtenue avec le masque 18mm-
20X par dépôt métallique . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
2.9 Images de phase quantitative de lamelles rainurées obtenues avec les masques conçus
pour trois grossissements (10X, 20X et 40X) . . . . . . . . . . . . . . . . . . . . . . 45
2.10 Réfractométrie en temps réel lors des transitions de liquides . . . . . . . . . . . . . . 49
2.11 Réfractométrie en temps réel lors d’ajouts graduels d’iodixanol . . . . . . . . . . . . 50
vii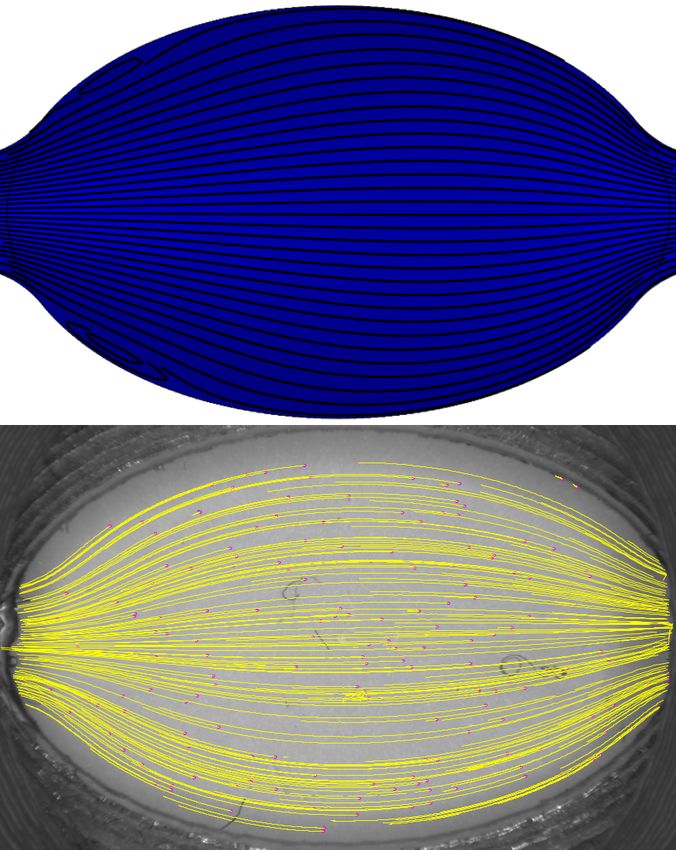
3.1 Réfractométrie en temps réel et QPS lors d’une transition entre le mannitol et le Ny-
condenz® . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
3.2 Paramètres biophysiques issus de multiples découplages lors d’une transition de liquide 54
3.3 Annulation du signal de phase quantitative de régions sous-cellulaires . . . . . . . . 57
4.1 QPS et masse sèche d’un astrocyte lors d’une perfusion de glutamate . . . . . . . . . 61
4.2 Essai fluidique de stimulation glutamatergique avec gradient d’indice de réfraction
extracellulaire . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
4.3 Volume en fonction du temps des astrocytes suite à la perfusion de glutamate . . . . 65
4.4 Indice de réfraction mesuré à différents endroits dans la rainure . . . . . . . . . . . . 67
A.1 Intensité moyenne de l’hologramme lors de l’écoulement d’une solution d’encre de
chine dans la chambre d’imagerie à grand champ de vue et utilisant le système de
perfusion à gravité . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71
B.1 Lissage de l’indice de réfraction du milieu en temps réel sur le QPS moyen cellulaire 73
viii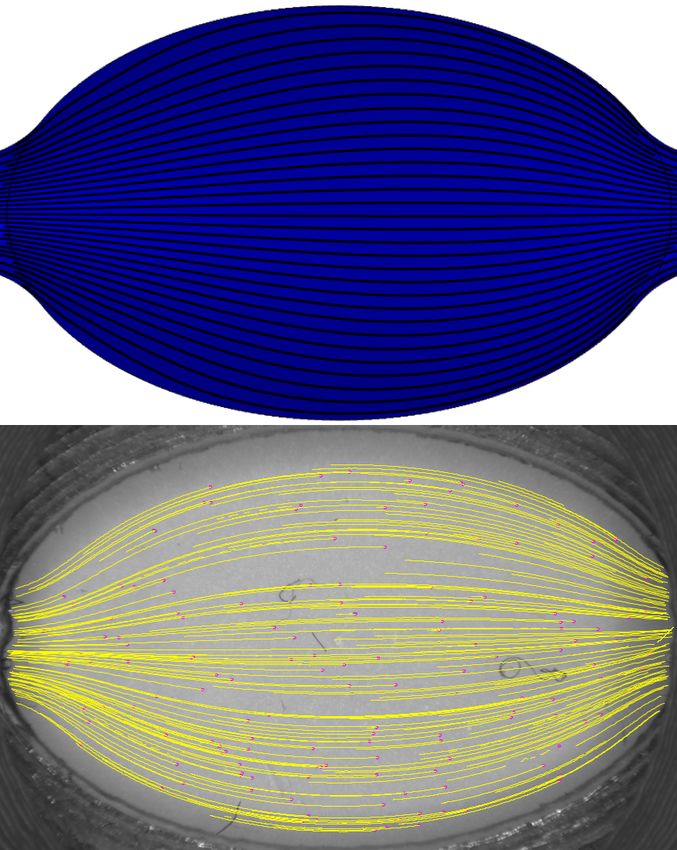
Liste des abbréviations
CAO Conception Assistée par Ordinateur
CCD Charged-Coupled Device
CFD Dynamique des fluides numérique (Computational fluid dynamics)
CMOS Complementary Metal-Oxide-Semiconductor
COPL Centre d’Optique, Photonique et Laser
DHM Microscopie holographique numérique (Digital Holographic Microscopy)
DMEM Dulbecco’s Modified Eagle Medium
FFT Tranformée de Fourrier Rapide (Fast-Fourrier Transform)
GLAST Glutamate Aspartate Transporter
NA Ouverture numérique (Numerical Aperture)
PLO Poly-L-Ornithine
QPI Imagerie de phase quantitative (Quantitative Phase Imaging)
QPS Signal de phase quantitative (Quantitative Phase Signal)
RI Indice de réfraction (Refractive Index)
RIE Gravure par ions réactifs (Reactive Ion Etching)
µPIV Vélocimétrie par imagerie de micro-particules (Micro-Particle Image Velocimetry)
ix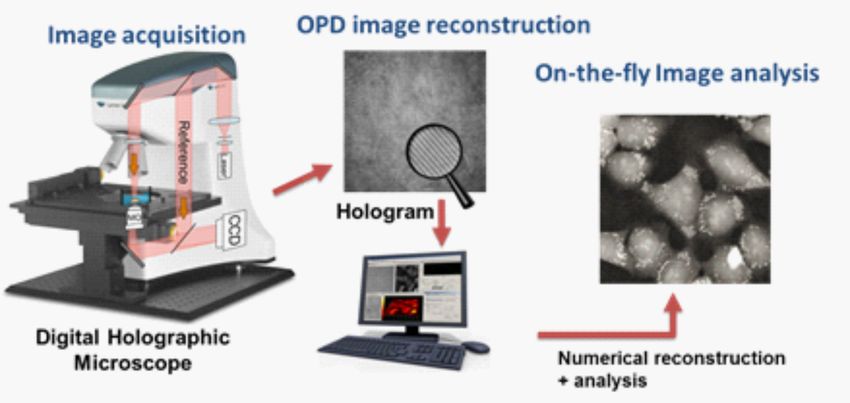
Remerciements
De nombreuses personnes m’ont aidé à accomplir le travail présenté dans ce manuscrit. J’aimerais
commencer par remercier mon directeur de recherche Pierre Marquet pour cette incroyable opportu-
nité qui m’a tant appris. Je dois aussi une grande partie de mon travail à mon co-directeur Erik Bélan-
ger, qui m’a constamment conseillé et guidé dans mes travaux. Je n’aurais pas pu avoir un meilleur
superviseur.
J’aimerais maintenant remercier l’ensemble de mes collègues, présents et passés, en particulier Sébas-
tien Lévesque que je considère comme un deuxième superviseur. Je veux aussi mentionner Marie-Ève
Crochetière et Chloé Martel pour m’avoir directement assisté à de maintes reprises pour la réalisa-
tion de mes travaux. J’ai aussi reçu de l’aide de plusieurs techniciens du COPL. Merci à Souleymane
Toubou Bah et Stéphane Gagnon, ainsi qu’à Hugues Auger pour m’avoir formé et appuyé lors de mes
passages au COPL.
Je veux remercier mes parents qui m’ont toujours soutenu et écouté. Finalement, je dois remercier ma
copine Jade. Merci de m’avoir constamment encouragé tout au long de ma maitrise, surtout dans les
moments les plus incertains.
xAvant-propos
Ce mémoire est divisé en quatre chapitres. Le premier chapitre, ainsi qu’une partie de mon introduc-
tion sont très fortement inspirés des travaux d’un article publié dans Frontiers in Physics dans lequel
est présenté plusieurs de mes travaux de maîtrise :
Bélanger E, Lévesque SA, Rioux-Pellerin É, Lavergne P and Marquet P (2019) "Mea-
suring Absolute Cell Volume Using Quantitative-Phase Digital Holographic Microscopy
and a Low-Cost, Open-Source, and 3D-Printed Flow Chamber," Front. Phys. 7 :172. doi :
10.3389/fphy.2019.00172
Cet article est essentiellement le point de départ de plusieurs résultats présentés dans les chapitres
subséquents et est nécessaire pour comprendre ces avancements. Il n’est toutefois pas inséré dans
son intégralité. Erik Bélanger et Pierre Marquet ont supervisé l’ensemble des travaux et ont écrit
l’article avec l’aide de Sébastien Lévesque, Pauline Lavergne et moi-même. Sébastien Lévesque et
Pauline Lavergne ont réalisé les expériences d’imagerie cellulaire avec le DHM, incluant le traitement
d’images. J’ai réalisé la fabrication et la caractérisation de la chambre imprimée en 3D. J’ai aussi
analysé l’ensemble des données. Ainsi, j’ai traduit librement les sections résultant directement de mon
travail et j’ai résumé les sections résultant des travaux d’analyses. J’ai omis délibérément quelques
sections auxquelles je n’ai pas participé et qui sont sans importance pour la suite de ce mémoire.
À l’inverse, j’ai pris la liberté d’ajouter quelques sections détaillant mon travail dans le cadre de
cette étude qui n’avaient pas été mentionnées dans la publication. Finalement, car les enjeux décrits
dans l’article sont essentiellement les mêmes que ceux de ce mémoire, les introductions des deux
manuscrits ont certaines ressemblances.
Au cours de ma maîtrise, j’ai aussi contribué à un second article :
Bertrand de Dorlodot, Erik Bélanger, Émile Rioux-Pellerin, and Pierre Marquet, "Si-
multaneous measurements of a specimen quantitative-phase signal and its surrounding
medium refractive index using quantitative-phase imaging," Opt. Lett. 45, 5587-5590
(2020)
Bien que certains aspects présentés dans ce mémoire (aux chapitres 2 et 3) ont mené à ce qui est
présenté dans cette seconde publication, aucun texte ni figure en est tiré directement.
xiIntroduction
0.1 Importance du volume cellulaire et des mouvements d’eau
transmembranaires
Les cellules eucaryotes sont caractérisées par leur noyau et sont délimitées par une membrane bi-
couche lipidique. Elles nécessitent d’échanger avec leur environnement via leur membrane pour leur
bon fonctionnement. À cause de ce besoin intrinsèque, les cellules sont constamment dans le pro-
cessus de transporter leur contenu vers l’espace extracellulaire et d’intégrer des biomolécules ou des
ions [1; 2]. Une majorité d’activités cellulaires ont un impact sur les concentrations intracellulaires
d’ions ou de biomolécules et donc aussi sur leur osmolarité. La membrane des cellules animales est
perméable à l’eau par l’intermédiaire de nombreux canaux transmembranaires nommés aquaporines.
La perméabilité passive de cette membrane implique que les changements d’osmolarité intracellulaire
seront nécessairement suivit par des mouvements d’eau transmembranaires qui auront un impact sur
le volume cellulaire. Or, plusieurs fonctions cellulaires dépendent de l’homéostasie ionique intracel-
lulaire essentielle, incluant les mécanismes d’expression de gènes, les cascades de seconds messagers
et la production d’hormones [3; 4]. Pour assurer ces processus cellulaires, le volume est sévèrement
régulé principalement par l’influx d’ions Na+ , K+ et Cl− [5]. Ainsi, le volume cellulaire est un para-
mètre biophysique qui reflète fidèlement la capacité homéostatique d’une cellule et en particulier les
mouvements d’eau transmembranaires. Dans le cadre des objectifs du LRNP, ayant pour but d’iden-
tifier des biomarqueurs pour les troubles psychiatriques majeurs (i.e la schizophrénie, la bipolarité et
la dépression majeure), il devient particulièrement intéressant de se pencher sur la résistance des cel-
lules aux changements de volume par la régulation de ce dernier, directement reliée à l’homéostasie
et fragilisée dans des cas pathologiques [6; 5].
Toutefois, une bonne mesure de volume en contexte d’imagerie cellulaire non-invasive est encore
difficilement réalisable [7]. Pour l’étude d’une grande quantité de cellules en suspension, la cytomé-
trie en flux est la méthode la plus efficace [8]. Toutefois, en ce qui concerne les cellules en culture,
représentant un milieu où les cellules peuvent croître et communiquer, il n’y a pas de consensus mé-
thodologique. Utilisant les outils les plus conventionnels de biophotonique, le volume peut être estimé
avec l’absorption lors de déplacements de colorants générés par les cellules [9] et aussi par l’exclusion
de fluorescence [10; 11]. Ces deux techniques nécessitent cependant un grand travail de calibration.
1L’imagerie de phase quantitative (QPI) est bien positionnée pour explorer ce domaine d’étude, par son
habilité à faire des mesures à haute résolution non-invasives d’échantillons transparents, tels que des
cellules en cultures, sans marquage et en temps réel [12; 13]. La QPI est un ensemble de technique
d’imagerie, incluant parmi d’autres, l’équation du transport d’intensité, la microscopie par phase de
fourrier ainsi que d’autres techniques basées sur l’holographie [14]. Dans le but de pouvoir mesurer la
dynamique du volume cellulaire de manière non-invasive, j’ai réalisé un ensemble de développements
technologiques à partir d’une approche d’imagerie de QPI, la microscopie holographique numérique
(DHM)
0.2 Microscopie par holographie numérique
La microscopie holographique numérique est basée sur un principe d’holographie, c’est-à-dire le pa-
tron d’interférence entre la lumière diffractée par un objet et la lumière d’une source de référence
cohérente. Tout délai de phase induit dans le front d’onde par un objet produira un changement dans
le patron d’interférence à la recombinaison des deux faisceaux. Bien que l’holographie n’est pas for-
cément numérique, l’enregistrement des hologrammes sur des caméras CCD présente l’avantage de
reconstruire numériquement l’objet générant l’hologramme. En holographie standard, deux images de
l’objet (une réelle et une virtuelle) sont obtenues avec la diffraction d’une source lumineuse traversant
l’hologramme avec un angle précis. La reconstruction numérique de l’hologramme utilise un concept
similaire multipliant l’hologramme à une onde de référence virtuelle et propageant le front d’onde
avec les équations de diffraction de Fresnel (entre autres)[15; 16; 17]. Schnars et Jüptner [18] furent
les premiers à utiliser un microscope holographique avec une caméra CCD permettant de reconstruire
les hologrammes numériquement et utilisant une configuration hors-axe. La reconstruction du front
d’onde est le produit de diffractions, elle sera constituée de plusieurs ordres en quatre contributions
[19] :
Ψ = RIh = R|R|2 + R|O|2 + |R|2 O + R2 O∗ . (1)
Le dernier terme est l’image réelle et est donc la contribution qui nous intéresse. Dans une confi-
guration alignée sur l’axe, c’est-à-dire que les deux faisceaux sont recombinés parallèlement un à
l’autre, il y aura un chevauchement des ordres dans le plan de Fourrier, rendant impossible l’isolation
de celui désiré. Une façon d’outrepasser cette limitation est d’incliner un des faisceaux pour qu’il se
recombine avec un léger angle, séparant dans le plan de Fourrier les deux ordres. La région d’intérêt
correspondant à l’image réelle peut être conservée.
Les équations de diffraction de Fresnel, sous la forme de transformée de Fourrier, sont utilisées pour
propager une onde sphérique partant de l’hologramme filtré jusqu’au plan de l’objet (voir la Figure
0.1). Sous sa forme numérique (transformée de Fourrier rapide (FFT)), le front d’onde reconstruit à la
2F IGURE 0.1 – Géométrie considérée pour la reconstruction d’un hologramme. Image tirée de Cuche
et al. [19].
forme de l’équation suivante :
iπ 2 2
Ψ(m,n) =A exp (m ∆ξ + n2 ∆η 2 )
λd
(2)
iπ 2 2 2 2
× FFT IH (k,l) exp (k ∆x + l ∆y ) ,
λd m,n
où k,l,m et n sont des entiers, λ est la longueur d’onde du laser, d la distance de reconstruction et IH est
l’hologramme numérique enregistré sur une caméra CCD N × N. En prenant en compte la courbure
de l’onde crée par un objectif (ou n’importe quel élément optique grossissant) et la multiplication
de l’hologramme par une onde de référence numérique, l’équation de propagation du front d’onde
devient :
iπ 2 2 2 2
Ψ(m,n) =AΦ(m,n) exp (m ∆ξ + n ∆η )
λd
(3)
iπ 2 2 2 2
× FFT RD (k,l)IH (k,l) × exp (k ∆x + l ∆y ) .
λd m,n
La matrice Φ(m,n) est un masque de phase permettant de corriger des aberrations numériquement et
RD (k,l) est la matrice de l’onde de référence numérique. Suite à cette rétro-propagation, une matrice
complexe est obtenue, où la partie réelle est équivalente à l’amplitude du front d’onde à la distance de
propagation et la partie imaginaire est la phase :
I(m,n) = Re[Ψ(m,n)]2 + Im[Ψ(m,n)]2
!
Im[Ψ(m,n)] . (4)
φ (m,n) = arctan
Re[Ψ(m,n)]
Avec l’argument de Ψ, il est donc possible d’obtenir une image de phase quantitative φ (m,n). En pra-
tique, l’utilisation de la DHM passe donc par deux étapes clés. L’acquisition de l’hologramme et sa
3reconstruction numérique en image de phase quantitative, un processus schématisé par la Figure 0.2,
ouvrent la porte à plusieurs avantages. La reconstruction peut être faite en temps réel ou subséquem-
ment à l’acquisition lors d’enregistrement rapide. La distance de reconstruction peut être déplacer lors
des reconstructions, et même être différente d’image en image.
F IGURE 0.2 – Schématisation du processus d’imagerie avec la microscopie holographique numérique.
Image adaptée de Rappaz et al. [20]
Au même titre que plusieurs autres méthodes d’imagerie de phase quantitative, la DHM ce distingue
de la microscopie à contraste de phase conventionnelle. La DHM permet de mesurer un délai de phase
précisément (allant jusqu’à une fraction de degré). Dans sa forme intégrale, le signal d’un pixel de
l’image de phase quantitative est exprimé ainsi :
Z T
2π
φ (x,y) = n(x,y,z)dz, (5)
λ 0
où n(x,y,z) est l’expression de l’indice de réfraction dans l’axe vertical (z) et (x,y) est un pixel donné.
Il est toutefois plus utile de parler de différence de phase ∆φ lorsqu’on mesure un objet précis pour
obtenir sa contribution exacte. La différence de phase est obtenue en soustrayant le délai de phase de
l’objet d’étude à la valeur de référence de l’image :
∆φ (x,y) = φ (x,y) − φre f . (6)
Dans un contexte d’étude de cellules en culture imagées dans une chambre de perfusion (voir la
Figure 0.3) ces équations peuvent être détaillées en fonction des objets dans le trajet de la lumière.
Le laser traverse la première lamelle de la chambre, le volume au-dessus de l’échantillon, la cellule et
la dernière lamelle. La contribution des lamelles peut être ignorée car elle sera la même partout dans
l’image. La contribution au délai de phase de la cellule est séparée :
4φref φc
nc nm
hc
F IGURE 0.3 – Schéma d’une vue de côté d’une cellule dans une chambre d’imagerie et définition de
variables.
2π T
Z
φ (x,y) = nc (x,y,z)dz
λ 0
Z h Z T
2π c
= nc (x,y,z) + nm (z) dz (7)
λ 0 h
2π
= hc (x,y)nc (x,y) + T nm − hc (x,y)nm ,
λ
où n̄c (x,y) est l’indice de réfraction intracelluaire intégré, hc (x,y) est la hauteur de la cellule, nm est
l’indice de réfraction extracellulaire et T est l’épaisseur du canal de la chambre de perfusion. Géné-
ralement, le QPS est moyenné sur l’ensemble de l’échantillon pour mitiger les effets du bruit, donc le
couple (x,y) précisant un pixel donné sera omis pour la suite. Utilisant la définition de la différence de
phase, le signal de phase quantitative (QPS) cellulaire est :
∆φ̄c = φ̄c − φre f
2π
= h̄c n̄c + T nm − h̄c nm − T nm (8)
λ
2π
= (n̄c − nm )h̄c .
λ
Nous pouvons donc observer que dans un contexte cellulaire, le QPS dépend de trois variables, l’épais-
seur moyenne de la cellule (h̄c ), l’indice de réfraction moyen cellulaire (nc (x,y)) et l’indice de réfrac-
5tion du milieu (nm ). Les deux premières étant des paramètres biophysiques cellulaires et alors que la
troisième est en relation directe avec les conditions expérimentales.
0.3 Calculs de paramètres biophysiques
La richesse du signal de phase est aussi bien un avantage qu’un inconvénient. D’une part, le signal
dépend de paramètres intrinsèques aux échantillons biologiques et un changement dans ces para-
mètres sera ultimement détectable. Toutefois, même en connaissant précisément l’indice de réfraction
extracellulaire, il reste encore une ambiguïté dans le signal de phase : il peut varier soit avec un
changement d’indice de réfraction cellulaire ou de son épaisseur. En effet, il est difficile d’extraire la
contribution de chaque paramètre. Les deux étant plus riches en information sur la cellule que la phase
seule, il est désirable d’être en état de pouvoir les déterminer.
Avec l’imagerie quantitative de phase, quelques méthodes peuvent être utilisées pour outrepasser le
couplage de l’épaisseur et de l’indice de réfraction. Une des plus simples est de fixer un indice de
réfraction moyen et d’assumer l’homogénéité de la cellule. La contrepartie de cette méthode serait de
fixer, à l’aide de modèles, la géométrie de la cellule pour en déduire l’indice de réfraction cellulaire
[21]. Ces deux techniques seraient particulièrement pertinentes pour les types cellulaires qui ont peu
de variabilité dans leur contenu intracellulaire ou encore possèdent une forme prédictible, i.e des
globules rouges.
Toutefois, plusieurs types cellulaires ne sont pas aptes à l’utilisation de ce type d’approximation. Des
techniques alternatives doivent être employées pour découpler les paramètres biophysiques du signal.
Car le QPS dépend aussi d’autres variables (jusqu’à présent considérées fixes), il est possible de
prendre deux images du même échantillon dans des conditions différentes en changeant, par exemple,
la longueur d’onde du laser, ou encore l’indice de réfraction du milieu d’immersion créant un système
de deux équations avec deux inconnus. La DHM à double longueurs d’onde, quoique présentant un
grand potentiel pour la rapidité de la mesure [22] reste à ce jour difficile à implémenter dû à son
montage qui nécessite l’acquisition simultanée de deux hologrammes orthogonaux et l’utilisation de
colorants de dispersion incompatible avec plusieurs types cellulaires. La seconde solution, bien que
moins rapide, est relativement plus facile à implémenter. En prenant, deux images dans deux milieux
d’indice de réfraction différents, deux équations pour le QPS sont postulées [23; 24; 25] :
∆φ̄c1 = 2π/λ (n̄c − nm1 )h̄c n̄c (9)
∆φ̄c2 = 2π/λ (n̄c − nm2 )h̄c n̄c . (10)
Dans les Équations 9 et 10, l’indice de réfraction cellulaire n̄c et l’épaisseur cellulaire h̄c sont les
mêmes. Assumant aucun changement entre les deux images, il est possible d’isoler les deux variables :
6∆φ̄c2 nm1 − ∆φ̄c1 nm2
n̄c = (11)
∆φ̄c2 − ∆φ̄c1
λ0 (∆φ̄c2 − ∆φ̄c1 )
h̄c = . (12)
2π nm1 − nm2
Même si ces deux équations sont théoriquement valides pour chacun des pixels comprenant la cellule.
Pour limiter les erreurs potentielles dues au bruit et au micro-changements cellulaires, il est utile de
moyenner la phase sur tout le corps cellulaire avant de calculer l’indice de réfraction. Conséquemment,
les valeurs d’indices de réfraction et de hauteur seront des moyennes, respectivement pondérées et
géométriques.
Bien que ces deux paramètres biophysiques soient intéressants en soi, lorsqu’ils sont conjugués à
une segmentation de l’image, d’autres paramètres clés dans l’étude des cellules vivantes peuvent être
obtenus. On peut voir facilement que la hauteur cellulaire moyenne (h̄c ), multipliée à la surface de la
cellule (Sc ) permet d’obtenir un volume cellulaire. Car h̄c est une véritable moyenne géométrique, le
volume mesuré par h̄c · Sc est bien le volume absolu de l’échantillon.
L’indice de réfraction permet lui aussi de déterminer d’autres paramètres biophysiques, principalement
par sa relation avec la concentration en molécules et en ions dans le cytoplasme. En effet, ce qui
différencie l’indice de réfraction cellulaire de celui de l’eau est la quantité d’autres molécules en
solution [26; 27]. Cette relation est linéaire et peut être exprimée comme une addition :
n̄c = neau + αC (13)
La constante d’incrément réfractif spécifique α dépend de la substance en solution, mais demeure
entre 0,017 à 0,019 100g/mL pour les échantillons biologiques. Cette relation peut être utilisée aussi
pour déterminer la fraction d’eau massique et volumique (à l’aide de quelques approximations) [28] :
W = 1 − 0,7 ·C (14)
W
Wm = . (15)
1,4 − 0,4W
L’influence qu’a la concentration des constituants intracellulaires sur l’indice de réfraction cellulaire a
aussi un effet indirect sur la mesure de la phase. Il est alors possible de relier la phase d’une cellule à la
concentration de son contenu intracellulaire sans faire de découplage. Il faut noter que le changement
de l’indice de réfraction est causé par le contenu intracellulaire, différentiant le cytoplasme de l’eau.
Toute variation entre l’indice de réfraction de l’eau et celui de la cellule est due aux protéines, ions et
acides nucléiques. Ainsi, si la cellule est observée dans l’eau avec l’imagerie de phase quantitative, la
7différence de phase entre la référence et la cellule sera reliée à la concentration réelle de son contenu
intracellulaire. Il est possible de calculer la masse que représente cette concentration :
∆φ̄c = α ·C · h̄c
Sc
∆φ̄c = α h̄c . (16)
h̄c
∆φ̄c Sc
DM =
α
On peut donc obtenir la masse sèche en multipliant la phase par la surface et en divisant par la
constante α. Bien qu’il n’est pas nécessaire de changer de liquide pour avoir une mesure dynamique
de la masse sèche, surtout utilisée en contexte de croissance cellulaire [29; 30], la segmentation de Sc
doit être la plus inclusive possible pour obtenir la masse totale de la cellule.
0.4 Mécanique des fluides
Il a été répété plusieurs fois que la procédure de découplage à deux liquides implique l’imagerie d’un
même échantillon dans deux liquides d’indices de réfraction différents. Cette technique assume aussi
aucun changement de l’objet entre les deux images utilisées. Pour minimiser les chances de produire
des erreurs, il est avantageux de faire le changement de liquide le plus efficacement possible. À cet
effet, pour la conception du dispositif fluidique, il est important d’avoir une certaine compréhension
des mécanismes de mécanique des fluides.
Les champs d’écoulements se comportent tels que définis par les équations de Navier Stokes [31].
Bien qu’il ne soit pas nécessaire de les connaître parfaitement, il faut se souvenir que les solutions
numériques des simulations en sont dérivées. L’écoulement d’un fluide se fait généralement dans l’un
de deux régimes : laminaire ou turbulent. Le premier est caractérisé par un profil de vitesse parabolique
avec aucun mélange entre des strates de liquide, alors que le second est caractérisé par la création de
vortex, à toutes fins pratiquement aléatoires, se déplaçant aussi dans la direction du flux. Le régime de
l’écoulement peut être estimé en calculant le nombre de Reynolds :
ρvD
RE = (17)
µ
où ρ est la densité du fluide, v est sa vitesse,µ est sa viscosité et D est le diamètre du conduit. La valeur
de RE=1400 est généralement considérée comme la transition entre les deux régimes ; laminaire en
dessous et turbulent au-dessus. La Figure 0.4 présente la forme de l’écoulement laminaire.
La création de vortex est tout de même possible lorsqu’un changement de diamètre se produit dans
le conduit. Le phénomène est connu comme les pertes mineures [31]. La taille de ces vortex est
relativement prédictible par l’angle du diffuseur entre les deux sections du conduit, un changement
8Profil de vitesse
Vitesse = 0
F IGURE 0.4 – Schéma du profil de vitesse pour un écoulement laminaire.
sans diffuseur aurait un angle de 90°. Il existe un angle optimal de diffuseur, soit d’un demi-angle
d’environ 4° pour un conduit circulaire. Cet angle change légèrement en fonction de la géométrie
de la section du tuyaux [32]. Ces principes théoriques de mécaniques de fluides ont été utilisés pour
le développement des dispositifs fluidiques décrits dans les chapitres subséquents du mémoire, mais
aussi lors de l’élaboration des protocoles de perfusion.
0.5 Astrocytes
L’étude détaillée des astrocytes, un type de cellules gliales, est relativement récente [33]. Si les as-
trocytes ont longtemps été considérés comme des cellules de remplissage du système nerveux central
[34], il est maintenant bien établi qu’ils accomplissent un très grand nombre de fonctions, qui vont
d’assurer le soutien énergétique pour les neurones jusqu’à la modulation de la transmission synap-
tique [35; 36] en passant par la régulation du volume extracellulaire [37; 38]. Leurs nombreux rôles
les impliquent aussi dans plusieurs pathologies neuronales, un exemple notable étant l’épilepsie ; de
plus, ils sont en partie responsables de l’immunité du cerveau [38].
Dans le cas spécifique des travaux présentés dans ce mémoire, la capture de neurotransmetteurs, plus
précisément la capture du glutamate par les astrocytes sera utilisée comme paradigme biologique
pour mesurer la dynamique du volume cellulaire. La capture de glutamate est multifonctionnelle ; non
seulement cet apport dans les astrocytes sert à réguler l’effet du glutamate dans la fente synaptique
mais permet d’en éviter des concentrations cytotoxiques [39]. Cette capture de glutamate stimule
aussi les fonctions métaboliques des astrocytes, provoquant le transport de glucose ainsi que la gluco-
lyse [40; 39]. L’entrée de glutamate par les co-transporteurs glutamate-sodium (GLAST) produit un
gonflement des astrocytes même après la stimulation, et ce pour deux raisons : le glutamate ainsi inter-
nalisé induit des influx d’eau transmembranaires pour des raisons osmotiques, mais aussi puisque le
GLAST transporte lui-même de l’eau lorsqu’il est activé [41]. Ce gonflement induit par glutamate sera
utilisé comme modèle, tentant d’ajouter une technique supplémentaire à l’étude de ce type cellulaire
complexe.
9Chapitre 1
Mesures de volume de cellules entières
avec découplage à deux liquides
Ce chapitre sera dédié à la description et discussion du travail effectué dans le cadre de l’article publié
dans Frontiers in Physics, Bélanger et al. [25]. Cette publication présente et détaille la production,
caractérisation et utilisation d’une chambre d’imagerie sur mesure imprimée en 3D conçue pour la
procédure de découplage à deux liquides. Plusieurs aspects de cette publication sont les fondements
des travaux qui seront présentés dans les chapitres subséquents de ce mémoire. Ainsi un retour sur
certains des concepts et des travaux effectués dans l’article, avec l’addition de quelques précisions sur
les sections résultant directement de mes travaux est nécessaire pour cadrer le reste des travaux de la
maîtrise. Je ne suis pas le seul contributeur des résultats présentés dans ce chapitre, les Sections 1.1.4,
1.1.5 et 1.1.6 ont été incluse uniquement car elles sont nécessaires pour comprendre les résultats de ce
chapitre et des chapitres suivants. Aussi, j’ai uniquement traduit les figures que j’ai personnellement
réalisées.
1.1 Matériels et méthodes
1.1.1 Fabrication de la chambre d’écoulement
Les dessins de la conception assistée par ordinateur (CAO) de la chambre d’écoulement ont été dessi-
nés avec SolidWorks (2016, Dassault Systèmes). La chambre de perfusion a été imprimée en 3D avec
une Ultimaker Extented 2+ et en filament de plastique PETG (Guidel !ne, Taulman3D). La base et
le couvercle sont imprimés usant une buse de 0,4 mm (Micro Swiss), précisant un remplissage léger
dans le logiciel de tranchage (Cura 3.6.0, Ultimaker). Les deux parties de la monture ont chacune
quatre trous circulaires pour y coller (5551T72, McMaster-Carr) des aimants de terre rare (5862K961,
McMaster-Carr). Le joint contenu dans la base a été découpé d’une feuille de parafilm (M, BEMIS)
avec des ciseaux. L’élément central a été imprimé avec une buse de 0,25 mm (Micro Swiss) avec
un remplissage plein et un chevauchement de remplissage de 30%. Ce chevauchement prévient les
10fuites ; cependant, l’élément central doit être traité à l’acétone pour assurer l’étanchéité considérant
les niveaux de pression interne lors d’écoulement de fluides [42]. Seule la surface la plus rugueuse
(celle du dessus) est exposée aux vapeurs d’acétone (270725, Sigma-Aldrich) pour la durée recom-
mandée de 15 min. Suite à ce traitement, une lamelle de 18 mm est pressée dans le socle prévu à
cet effet la fixant en place et scellant l’élément central. La pression est appliquée et maintenue sur
la lamelle environ 1 min, les premières 10 s étant décisives. Des micro-tuyaux en acier inoxydable
(304F10125X006SL, MicroGroup) sont partiellement insérés et collés chaque côté de l’élément cen-
tral. Les tuyaux sont coupés en employant une technique appropriée pour éviter l’écroulement du
canal interne. Une lame aiguisée (LB, OLFA) est roulée sur le micro-tuyaux appliquant seulement
une faible force. Après quelques va-et-vient les tuyaux sont coupés sans altérer leur cavité.
1.1.2 Caractérisation de la chambre de perfusion
Simulations de dynamique des fluides
Des simulations en régime permanent ont été calculées avec le module de dynamique des fluides nu-
mérique (CFD) de COMSOL Multphysics (5.3a, COMSOL). Une section transversale vue de haut
a été utilisée pour définir la géométrie des simulations, assumant qu’une représentation 2D est une
approximation suffisante. Les micro-tuyaux ont été inclus dans la géométrie pour assurer une res-
semblance supérieure avec le montage expérimental, utilisant leur diamètre interne comme dimension
d’entrée. Pour les spécifications de l’influx laminaire au début de la chambre d’écoulement, la lon-
gueur d’entrée a été calculée pour chaque débit avec l’expression du nombre de Reynolds dans un
tuyaux circulaire, donnant des valeurs de 1 à 2 mm. Les débits laminaires ont été variés de 0,2 à 5,1
mL/min à l’entrée. Une condition frontière de non-glissement a été posée à tous les rebords de la
chambre, ainsi qu’une condition de pression nulle à l’issue du conduit. Pour les simulations, le fluide
coule de droite à gauche.
Vélocimétrie par imagerie de micro-particules (µPIV)
L’écoulement empirique dans la fente d’imagerie a été évalué avec un montage de vélocimétrie par
imagerie de micro-particules étant composé d’une caméra (Grasshopper3, FLIR) et d’un système
zoom à grossissement variable (MVL-7000, NAVITAR). La caméra a été équipée avec un senseur
CMOS monochrome 2,3-megapixel Sony IMX174 avec une taille de pixel de 5,86 µm et un taux
d’acquisition maximal de 163 images par seconde. Le zoom est une lentille macro possédant une
longueur focale variable de 18 à 108 mm. Le grossissement maximal est d’environ 1,1X à la distance
de travail minimale de 130 mm. Une solution de micro-particules a été réalisée en pipetant un volume
de 200 µL d’une solution 1% m/v de base de micro-sphères en polystyrène avec un diamètre moyen
de 104 µm (PPB-1000-5, Spherotech). Ensuite, les micro-sphères ont été suspendues dans 10 mL
d’une solution de PBS 1X contenant 1 mg/mL de BSA. Cette solution a été perfusée avec une pompe
péristaltique (PPS-1100, Scientifica) à des débits d’écoulement continus allant de 0,2 à 5,1 mL/min.
Au total, approximativement trois vidéos par débit, durant 3 s, ont été enregistrés pour chacun des
11quatre éléments centraux testés. L’extension TrackMate d’ImageJ [43] a été utilisée pour le tracé
des trajectoires des micro-particules dans les données de µPIV pour obtenir les lignes de courant
expérimentales.
1.1.3 Essai de viabilité de neurones
Un total de dix cultures de neurones primaires de rat ont été préparées et ensuite séparées en deux
groupes, cinq comme témoins et cinq utilisant notre chambre imprimée en 3D. Les cultures du groupe
témoin ont été laissées 30 min dans un pétri (08-772B, Fisher Scientific) baignant dans une solution
physiologique tampon avec HEPES contanant (en mM) 75 NaCl, 5,4 KCl, 1,8 CaCl2, 0,8 MgCl2,
0,9 NaH2PO4, 10 HEPES, 5 dextrose (D-glucose) et 50 mannitol. Le deuxième groupe de lamelles
a été monté dans la chambre d’écoulement et perfusé à 2,0 mL/min pendant 30 min avec la même
solution. Toutes les cultures ont ensuite été colorées pendant 3 min avec une solution de 0,04% de
Bleu de Trypan (MT25900CI, Fisher Scientific) et imagées avec un microscope optique (EVOS FL,
Invitrogen). Quatorze images à un grossissement de 20X ont été prises arbitrairement le long de l’axe
central de la fente d’imagerie, ainsi que deux à un grossissement de 4X et centrées dans la fente
d’imagerie. Les cellules mortes et vivantes ont été recensées manuellement dans ImageJ (v1.52p,
NIH) et identifiées avec le marquage au Bleu de Trypan. La viabilité générale des neurones a été
calculée avec les images 20X positionnées aléatoirement alors que les images au 4X avaient pour but
de quantifier des dépendances spatiales de survie cellulaire. Dans ce cas, les images ont été séparées
en quatre segments parallèles à la longueur de la fente d’imagerie. Les cellules vivantes et mortes ont
été comptées séparément dans chacun des segments.
1.1.4 Microscopie de phase quantitative par holographie numérique
Le microscope holographique numérique utilisé (T-1003, Lyncée Tec) est basé sur un interféromètre
Mach-Zender. À l’intérieur du microscope, un faisceau lumineux à une longueur d’onde de 666 nm
produit par une diode laser est séparé en deux, formant une onde objet et une onde de référence.
L’onde objet diffractée par l’échantillon interfère avec l’onde de référence produisant un hologramme
hors-axe enregistré par une caméra numérique. L’objectif du microscope a un grossissement de 20X
avec une distance de travail de 590 µm et une ouverture numérique de 0,70 (HC PL APO, Leica). La
caméra est équipée d’un senseur monochrome CMOS 2,3-megapixel Somy IMX174 avec une taille
de pixel de 5,86 µm et pouvant enregistrer à un taux de 164 images par seconde (acA1920-155um,
Basler). Les hologrammes numériques sont intégrés sur une période approximative de 100 µs et acquis
à une fréquence de 4 Hz. Le temps d’intégration a été ajusté attentivement avant chaque session
d’imagerie pour éviter de saturer les hologrammes. La configuration hors-axe permet la reconstruction
quantitative des images de phase quantitative à partir d’un seul hologramme. Cette reconstruction
numérique a été effectuée avec le logiciel associé au microscope (Koala, Lyncée Tec), résultant en un
champ de vue carré de 215 µm. L’algorithme de reconstruction est une simulation de l’illumination
de l’hologramme enregistré par une onde de référence numérique [44]. Il s’ensuit une correction
numérique des aberrations du front d’onde induites par les composants optiques, de l’objectif et de
12la configuration hors-axe du microscope à l’aide d’un hologramme de référence [45]. L’aplatissement
du fond de l’image est réalisé à l’aide de polynomiales [46].
1.1.5 Découplage à deux liquides
La procédure de découplage à deux liquides a été décrite au chapitre précédent. Concrètement, une
faible quantité de NaCl (25mM) d’un milieu iso-osmotique et isotonique d’une osmolarité appropriée
pour le type cellulaire à l’étude (220 mOsm/kg pour les neurones ou 340 mOsm/kg pour les HEK293T
et le NIH3T3) est remplacée par 50 mM de mannitol ou de Nycodenz® pour obtenir les milieux de
perfusion [24]. En pratique, l’indice de réfraction et l’osmolarité de chaque solution physiologique
ont été respectivement mesurés avec un réfractomètre d’Abbé (Abbemat MW, Anton Paar) à 666 nm
et un osmomètre à point de congélation (Micro-Sample model 3320, Advanced Instruments) avant la
session d’imagerie. L’indice de réfraction de la solution de mannitol est environ 1,3335, tandis que
celui pour le Nycodenz® est 1,3380. Finalement, la perfusion consécutive de ces solutions physiolo-
giques, réalisée avec un système de perfusion à gravité relié à une valve à distribution manuelle à huit
ports [47], a été effectuée successivement trois fois ou encore avant et après un choc osmotique. Pour
toutes les solutions physiologiques, le débit d’écoulement a été fixé à 2 mL/min.
1.1.6 Choc osmotique
Des cultures de trois types cellulaires (neurones primaires de l’hippocampe, cellules NIH3T3 et
HEK293T) ont été montées dans au moins quatre éléments centraux différents et ont été soumises
à des chocs hypo et hyper-osmotiques. Dans ces expériences, les neurones primaires ont été perfu-
sés avec une solution physiologique avec tampon HEPES et une osmolarité similaire au Neurobasal
+ B27® contenant (en mM) 75 NaCl, 5,4 KCl, 1,8 CaCl2 , 0,8 MgCl2 , 0,9 NaH2 PO4 , 10 HEPES,
5 dextrose (D-glucose), 50 manniol ou Nycodenz® (pH 7,4 ; 225-230 mOsm). Les autres cultures
cellulaires (HEK293T et NIH3T3) ont été maintenues dans le DMEM ayant un plus grande osmo-
larité (330 mOsm) et ont donc été perfusées avec une solution iso-osmotique appropriée (en mM)
135 NaCl, 3 KCl, 5 dextrose, 10 HEPES, 3 CaCl2 , 1,8 MgCl2 et 50 mannitol ou Nycodenz® (pH
7.4 ; 330–335 mOsm). Le milieu hypo-osmotique a été préparé en retirant 25 mM de NaCl de la so-
lution iso-osmotique et au contraire, 25 mM de NaCl a été ajouté pour la solution hyper-osmotique,
résultant en des variations d’indice de réfraction respective de -0,0003 et 0,0003 vis-à-vis leur contre-
partie iso-osmotique. Toutes les cellules ont été perfusées avec leur solution respective préalablement
à l’enregistrement, commençant une fois que le signal quantitatif de phase (QPS) est stabilisé (nor-
malement environ après 2 min). Le niveau de référence (baseline) a été enregistré pendant environ 1
min pour ensuite être remplacé pendant approximativement 30 s par la solution iso-osmotique avec
Nycodenz® requise pour découpler les cellules au repos. Les cellules ont ensuite été soumises au choc
osmotique en perfusant pendant 5 min soit la solution hypo-omsotique ou hyper-osmotique contenant
le Nycodenz®. Finalement une fois le choc osmotique complété, une deuxième séquence de décou-
plage a été réalisée, alternant le milieu pour la solution au mannitol d’égale osmolarité pendant environ
1 min.
13Vous pouvez aussi lire

















































