La faune de l'Oppidum de Constantine. IIe - Ier s. av. J.-C. : Première approche.
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
La faune de l'Oppidum de Constantine.
IIe - Ier s. av. J.-C. : Première approche.
____________________________
Gaëlle Delpy (1)
(1)
Doctorante en Archéozoologie et Archéo-ichtyologie (2e année), Centre Camille Jullian – (Aix-
Marseille Université CNRS), UMR 6573 Archéologie Méditerranéenne et Africaine, Maison
Méditerranéenne des Sciences de l'Homme (MMSH), 5 rue du Château de l'Horloge, BP 647, 13094
Aix-en-Provence. maddy22d@aol.com. Directrice : S. Agusta-Boularot (2), Tutrice : M. Sternberg (3)
(2)
Professeur d'Archéologie et d'Histoire de l'art des mondes romains, Université Paul Valéry
Montpellier 3, UMR 5140, Route de Mende, 34 199 Montpellier Cedex 5.
(3)
CNRS, Centre Camille Jullian – (Aix-Marseille Université CNRS), UMR 6573 Archéologie
Méditerranéenne et Africaine, Maison Méditerranéenne des Sciences de l'Homme (MMSH), 5 rue du
Château de l'Horloge, BP 647, 13094 Aix-en-Provence.
____________________________
Mots-clefs : Constantine, oppidum, sanctuaire, religion, IIe-Ier s. av. J.-C., faune, élevage, chasse.
Résumé :
L'oppidum de Constantine est un site fortifié pourvu d'un sanctuaire Chthonien, occupé au
cours de deux périodes : au IIe-Ier s. av. J.-C. et au Ve-VIe s. ap. J.-C..
Durant sa première période d'occupation, l'élevage fut la principale source d'alimentation
carnée, avec une majorité d'Ovicaprinés et de Suidés, mais avec une présence non négligeable de
Bovidés. La priorité accordée au petit bétail (Ovicaprinés et Suidés) s'inscrit dans un schéma qu'on
retrouve sur des sites contemporains du pourtour de l'Etang de Berre. L'importance des Bovidés
semble cependant être propre au site, et diminuer dans la seconde phase d'occupation de l'oppidum
(Columeau, 2001). Ils passent alors de 29,2% à 20% des restes déterminés, tandis que les
Ovicaprinés deviennent les animaux nettement majoritaires avec 51,5% contre 35,5% dans la période
précédente. La proportion de Suidés reste stable, de même que celle des Lagomorphes et celle des
animaux chassés en général. La chasse n'apportait alors qu'un complément composé principalement
de Lagomorphes.
L'utilisation des animaux est visible sur les ossements, à travers des traces de découpe mais
aussi des marques de fragmentation osseuse. Elle pouvait avoir plusieurs buts : l'alimentation des
habitants, mais aussi le dépôt d'offrandes, la consommation rituelle ou la commercialisation.
1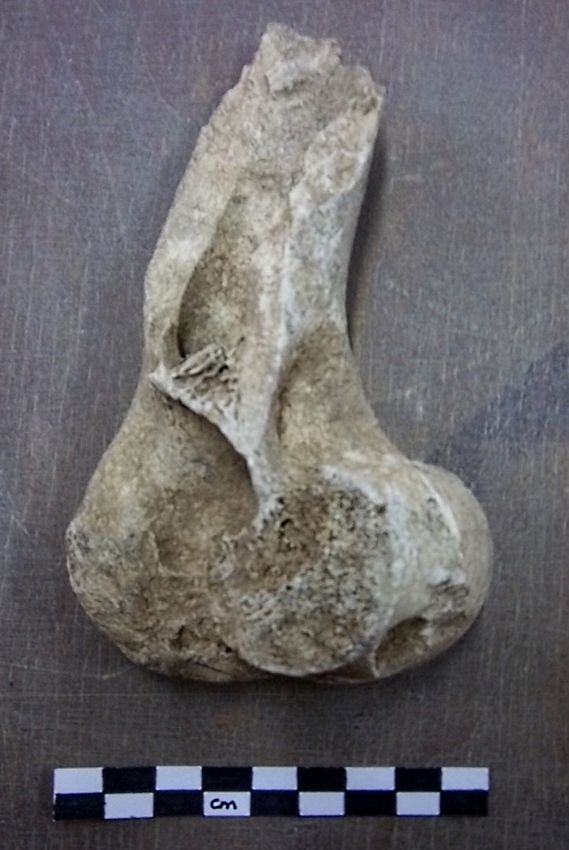
Introduction
L'oppidum de Constantine (Lançon-de-Provence, Bouches-du-Rhône) a connu une histoire
mouvementée. Il aurait été fondé durant la seconde moitié du II e s. av. J.-C., et fut abandonné dans
les années 20 – 10 av. J.-C.. Il fut réoccupé par la suite en fin du IVe – début du Ve s. ap. J.-C., pour
être délaissé à nouveau au VIe – VIIe s. ap. J.-C. (Gateau, 1996 ; Verdin, 2008, 2009).
Dès le XVIIe s., ce site a attisé la convoitise de bien des aventuriers et autres chercheurs de
trésor (Verdin, 1993, 2001). Loin de toute considération archéologique, l'intérêt qu'il suscitait alors
tournait autour de la légende d'un trésor enfoui dans l'aven naturel présent sur le site. Ceci au
détriment de l'oppidum qui eut à souffrir de ces nombreuses excavations.
Les fouilles sur ce site se déroulèrent de façon plus ou moins continue de 1824 jusqu'à nos
jours (Aubagnac, 1990 ; Verdin, 1993, 2001). Dès le début du XX e s., les indices d'une occupation
durant les Ve et VIe s. ap. J.-C. avaient été identifiés, et les structures bâties ainsi que le mobilier
découverts sur le site furent l'objet des travaux mentionnés précédemment. Le matériel faunique de
cette période historique, pour sa part, fut étudié par P. Columeau lors de la dernière phase de fouilles
débutée en 1990 (Columeau, 2001). Il révélait alors une prédominance des Ovicaprinés qui
représentaient 51,5% des restes déterminés, contre 25% de porcs et près de 20% de bœufs, tandis que
les restes d'animaux sauvages étaient quasi inexistants, mis à part quelques Lagomorphes (Columeau,
2001). Ces observations correspondent au schéma qu'il avait observé pour les autres sites
archéologiques du pourtour de l'Etang de Berre pour cette période. Il constatait en effet une
importance réduite des bœufs au profit des Ovicaprinés, tandis que la chasse n'avait plus qu'une place
très restreinte (Columeau, 1991, 1993). Mais qu'en est-il des périodes antérieures ?
Les fouilles menées depuis 2002 ont en effet permis de confirmer l'existence d'une première
période d'occupation du site au IIe-Ier s. av. J.-C. dont les restes fauniques n'avaient pas encore pu être
étudiés (Gourvest, 1956 ; Aubagnac, 1990 ; Verdin et al., 1992 ; Verdin, 2005). L'étude préliminaire
de ces ossements, qui fait l'objet de cet article, permettra de donner un aperçu du spectre faunique,
ainsi que des méthodes d'élevage et d'approvisionnement en produits d'origine animale de cette
époque. Elle apportera également des informations complémentaires aux recherches entreprises
précédemment, que ce soit sur l'oppidum de Constantine mais aussi sur les autres sites du pourtour
de l'Etang de Berre.
1/ Constantine, oppidum et sanctuaire
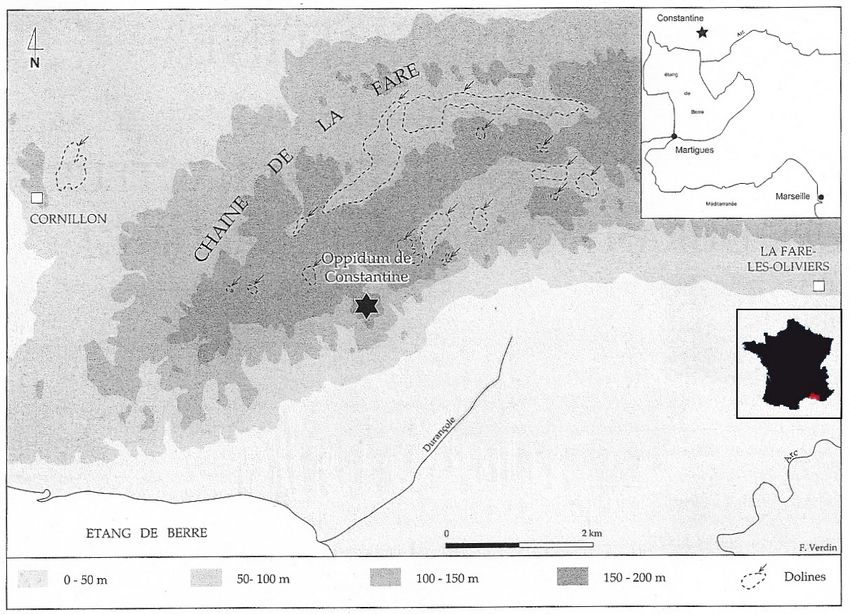
L'oppidum de Constantine est un site de hauteur (figure 1), situé sur un plateau d'une
superficie d'environ 6 ha au sud de la Chaîne de la Fare et à moins de 3 km au nord de l'Etang de
Berre. Il se trouve sur le territoire de la commune de Lançon-de-Provence, dans les Bouches-du-
2
Fig. 1 : Carte de localisation du site de Constantine (d'après Verdin, 2001).
Rhône.
Des défenses naturelles, qu'il s'agisse de fortes pentes ou d'escarpements discontinus,
complétées par la construction d'ouvrages fortifiés, assurent sa protection. Il dispose en outre d'un
sanctuaire Chthonien en forme de fer à cheval qui entoure deux avens naturels dont un de 65 m de
profondeur (figure 2).
La zone fouillée correspond au quartier situé au sud du sanctuaire et au réseau de rues
correspondant. Des pièces d'artisanat métallurgique, du mobilier céramique et en verre, des monnaies
ont été découverts sur place. L'étude de cette partie du site a révélé également de nombreux
remaniements des structures bâties. L'exemple le plus explicite de ces transformations est le
remplacement des bâtiments contemporains du sanctuaire daté de la fin du IIe et de la première
moitié du Ier s. av. J.-C. par un bâtiment à cour de 700 m² de superficie dans les années 50 – 30 av. J.-
C. (Verdin, 2008, 2009).
Pour ce travail, ce sont les restes fauniques découverts lors des fouilles menées en 2008 et
2009 sous la direction de F. Verdin (figure 2), et plus particulièrement ceux datés de la première
période d'occupation du site au IIe et Ier s. av. J.-C. (Verdin 2008, 2009), qui ont été étudiés.
3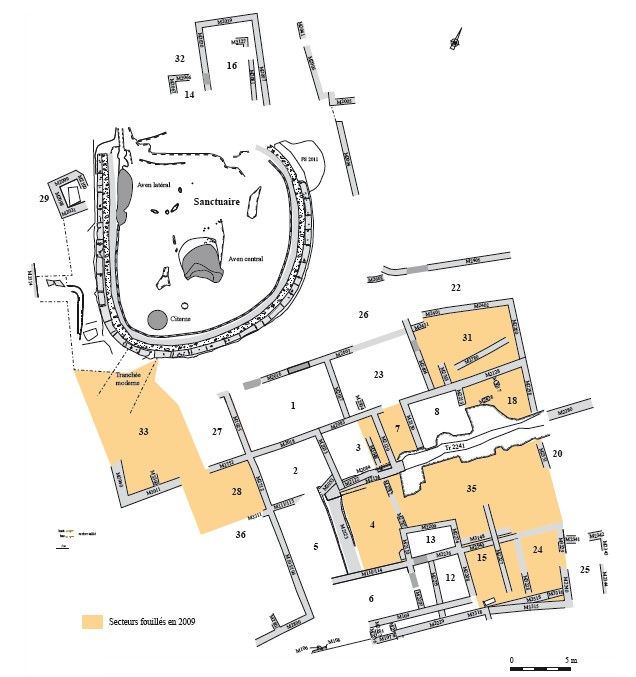
Fig. 2 : Plan général du site archéologique et plan de la zone fouillée en 2008 – 2009 (d'après Verdin,
2009).
2/ Matériel et méthodes
Les restes osseux découverts dans 79 unités stratigraphiques du site ont été étudiés de façon
classique. Ainsi, les ossements ont été examinés unité stratigraphique après unité stratigraphique.
Chaque fois que cela a été possible, les restes ont été déterminés anatomiquement et spécifiquement.
Cette détermination s'est faite par comparaison des ossements aux planches des ouvrages et fiches de
référence (Barone ,1976 ; Boessneck, 1990 ; Bressou, 1944 ; Helmer, 1987 ; Pavaux, 1976 ; Schmid,
1972), ainsi qu'à la collection de référence de J.-P. Brugal et aux conseils de M. Sternberg et M.
Leguilloux. Chaque fois que cela a été possible, les ossements ont été mesurés selon les normes
établies par A. Von den Driesch (Von den Driesch, 1976). Dans certains cas particuliers, tels que les
dents isolées par exemple, dont l'état de conservation était très variable, il a été nécessaire de
4
s'adapter avec un système de mesures particulier comportant deux mesures de la hauteur de la
couronne seule ou de la couronne avec la racine, selon que la dent en était encore pourvue ou pas. Le
nombre de ces restes déterminés est appelé NRD dans le tableau 1.
Tab. 1 : Description détaillée du nombre de restes datés du IIe – Ier s. av. J.-C..
(NRIndét. = nombre de restes indéterminés, NRDAnat. = nombre de restes déterminés uniquement anatomiquement,
NRD = nombre de restes déterminés spécifiquement et anatomiquement, NR = nombre de restes, % NR = pourcentage de
restes)
NR % NR
NRIndét. 1536 69,5
NRDAnat. 297 13,4
NRD 377 17,1
Total 2210 100
Lorsque les restes n'ont pu être déterminés qu'anatomiquement, ils ont été comptabilisés dans
la catégorie NRDAnat..
Les restes indéterminés (NRIndét.) ont été tous regroupés dans cette étude préliminaire, sans
distinction entre les esquilles d'os longs, les fragments d'os plats, etc. Une telle distinction pourrait
cependant s'avérer pertinente, et nécessitera un retour sur le matériel pour une étude plus fine. En
effet, cette différenciation pourrait permettre de mieux connaître la nature des restes majoritairement
présents sur le site, ce qui nous amènerait à mieux appréhender le type d'exploitation qui était fait des
animaux sur cet oppidum.
Le nombre élevé de restes indéterminés peut trouver son explication dans de nombreux
paramètres. Ainsi, outre le processus de conservation différentielle qui intervient de façon plus ou
moins importante sur chaque site archéologique, les nombreux remaniements du terrain évoqués
précédemment ont sans aucun doute nui à l'état des ossements. A ceci s'ajoute le fait que beaucoup de
restes proviennent de remblais et dépotoirs et s'en sont trouvés altérés. Néanmoins, ces problèmes de
conservation ne sont probablement pas les seuls à avoir participé à l'importante fragmentation des
restes osseux de Constantine. Certaines marques de découpe trouvées sur des ossements mieux
conservés permettent d'envisager l'hypothèse d'une exploitation des carcasses animales et des os à
des fins bouchères. Cet aspect sera développé plus en détail par ailleurs.
3/ Résultats
3.1/ Nombre d'individus et spectre faunique
A partir des restes déterminés anatomiquement et spécifiquement, le nombre minimum
5d'individus, ou NMI, de chaque espèce a été calculé. Pour cela, il a été nécessaire d'utiliser à la fois le
NMI de fréquence et le NMI de combinaison (Chaix et Méniel, 1996 ; Poplin, 1976, 1981), afin
d'avoir le NMI le plus fiable possible, tenant compte à la fois des os les mieux représentés mais aussi
de la latéralisation et de l'appariement possible ou non des restes. Les résultats obtenus sont présentés
dans le tableau 2 et la figure 3.
Tab. 2 : Tableau des nombres minimums d'individus par espèce.
(NR = nombre de restes, % NR = pourcentage de restes, NMI = nombre minimum d'individus, % NMI = pourcentage
d'individus)
Espèce NR % NR NMI % NMI
Bos taurus 110 29,2 28 23,1
Equidés 1 0,3 1 0,8
Ovicaprinés 134 35,5 37 30,6
Suidés 100 26,5 35 28,9
Gallus domesticus 5 1,3 3 2,5
Oiseaux de basse cour autres 2 0,5 2 1,7
Total animaux domestiques 352 93,3 106 87,6
Cervidés 2 0,5 2 1,7
Lagomorphes 13 3,5 9 7,4
Oiseaux autres (passereaux etc) 4 1,1 3 2,5
Micromammifères 6 1,6 1 0,8
Total animaux sauvages 25 6,7 15 12,4
Total déterminés 377
Total indéterminés 1536
Total 1913 100 121 100
Le trio d'espèces domestiques qui domine communément le spectre faunique des sites
archéologiques protohistoriques et antiques (Méniel, 1987) se retrouve à Constantine, à savoir les
Ovicaprinés, les Suidés et Bos taurus.
Moutons et chèvres sont les animaux les plus fréquents en nombre de têtes. A noter que les
restes osseux ne nous ont pas permis de les différencier. Les dents n'ont en effet pas permis de faire
de différence (Helmer, 2000), pas plus que certains os, comme les scapulas ou radius qui peuvent
habituellement permettre de faire la distinction entre chèvre et mouton (Boessneck, 1990). L'état de
conservation de ces restes s'est révélé insuffisant pour faire la différence.
Cette prépondérance est en accord avec la tradition pastorale fortement ancrée dans la région
rapportée notamment par Strabon (Géographie, IV, 1, 6) et Pline (Histoire Naturelle, XXI, 57) dans
leurs évocations de la plaine de la Crau, à une trentaine de kilomètres à l'ouest de l'oppidum. Ce
6pastoralisme est confirmé par les études paléoenvironnementales effectuées sur le pourtour de l'Etang
de Berre (Trément, 1999) ainsi que par les fouilles archéologiques régionales (Brun, 1996).
Autres animaux en grand nombre sur le site, les Suidés. Là encore, faire la distinction entre
porcs et sangliers n'a pas été possible, le nombre d'ossements permettant de déterminer la taille des
animaux étant trop restreint (Méniel, 1987). Deux ou trois canines incomplètes mais tout de même de
taille conséquente amènent à ne pas exclure la possibilité de la présence de quelques sangliers.
%
40
% NR (n = 377)
35 % NMI (n = 121)
30
25
20
15
10
5
0
r
s
és
B a res
s
s
Ce s
és
s
us
am ns
s
ou
dé
né
re
e
dé
ru
id
id
ph
tic
ie
t
ifè
ui
ni
ec
ri
rv
Su
au
au
ac
or
ap
Eq
es
Ca
m
sT
ss
tr
m
ux
m
vic
ba
go
o
ea
Bo
sd
O
m
La
de
is
ro
llu
O
ux
ic
Ga
M
ea
is
O
Fig. 3 : Histogramme représentant le spectre faunique selon le NR et le NMI.
(% NR : pourcentage de restes, % NMI : pourcentage d'individus)
Du IIe au Ier s. av. J.-C., sur une douzaine de sites de Narbonnaise, on retrouve une moyenne
de 44,7 à 64,8% de restes d'Ovicaprinés et de 12,4 à 35,5% de restes de Suidés (Columeau, 1991).
Par ailleurs, un équilibre entre les proportions d'Ovicaprinés et de Suidés est à noter dans les sites
répertoriés autour de l'Etang de Berre (Columeau, 1996). La faune de l'oppidum de Constantine ne
ferait donc que s'inscrire dans ce schéma régional, voire même local, avec 35,5% de restes
d'Ovicaprinés et de 26,5% de restes de Suidés. Cela reste cependant à confirmer, notamment par
l'ajout des restes fauniques découverts lors des fouilles effectuées en 2010.
La présence en contrebas de l'oppidum de terres cultivables pourrait expliquer la présence
importante de spécimens de Bos taurus qui auraient pu être utilisés autant pour le transport que pour
le travail de ces terres. En effet, ils ne représentent que de 8,9 à 12,2% des restes dans d'autres sites
7de Narbonnaise de la même période (Columeau, 1991) contre 29,2% pour ce site. Cette hypothèse
d'utilisation de la force de ces animaux se confirmerait d'après les premières estimations d'âge, le
plus jeune animal découvert ayant environ quatre ans. Elle n'est cependant pas vérifiée par de
potentielles déformations osseuses au niveau des phalanges ou des vertèbres, de telles pathologies
liées à ce type d'emploi des bœufs n'ayant pas été observées.
Le nombre d'animaux d'élevage est très supérieur à celui des animaux sauvages et
représenterait environ 90% des spécimens identifiés. Cela correspond aux rapports chasse/élevage
observés à cette période dans plusieurs sites provençaux et pour lesquels la proportion d'animaux
élevés se situe entre 87 et 92,8% (Columeau, 1991). Les Lagomorphes seraient le gibier privilégié,
étant attesté que la domestication de ces animaux remonterait au plus tôt au Moyen Age et qu'il ne
s'agirait donc pas d'animaux d'élevage (Callou, 2003). Ceci concorde à la fois avec l'environnement
de garrigue du site supposé comme déjà existant d'après les études palynologiques et
paléoclimatiques menées sur la région (Gateau, 1996), et avec les études fauniques effectuées aussi
bien en Provence que sur le pourtour de l'Etang de Berre (Columeau, 1991, 1996). Les deux
spécimens de Cervidés ont pu être pris plus en contrebas.
Ces chiffres sont cependant à prendre avec précaution du fait de l'état de fragmentation très
important des restes. Mais ce spectre faunique semble bien correspondre à la réalité du terrain
(Gateau, 1996 ; Trément, 1999) et aux pratiques agro-pastorales et cynégétiques connues pour
l'époque (Brun, 1996 ; Columeau, 1991, 1993, 1996, 2002, 2004).
3.2/ Répartitions anatomiques
Les restes osseux peuvent nous fournir d'autres types d'informations, notamment en étudiant
leur localisation anatomique pour chaque espèce, ainsi que leur fréquence (Figures 5 à 7).
Ainsi, on peut constater chez Bos taurus (figure 4) une répartition assez uniforme des restes
avec une représentation équivalente de toutes les pièces anatomiques, excepté une quantité de
vertèbres et de scapulas plus importantes que les autres types de restes. Cette différence peut
s'expliquer par les effets de la conservation différentielle, quoi que les os plats comme les scapulas
auraient plutôt tendance à disparaître plus facilement du fait de leur fragilité. Il peut aussi s'agir du
résultat d'une sélection de pièces bouchères ou bien de restes de déchets de boucherie (Lignereux et
Peters, 1996). L'importance des côtes et des dents n'est due pour sa part qu'au grand nombre naturel
de ces parties dans le squelette d'un animal.
8Nom des pièces
anatomiques Bos taurus (NR) Bos taurus (%)
crâne 1 0,93
maxillaire 0 0,00
mandibule 3 2,80
dent 27 25,23
vertèbre 8 7,48
côte 36 33,64
sternum 1 0,93
scapula 6 5,61
humérus 2 1,87
radius 3 2,80
ulna 0 0,00
carpien 1 0,93
métacarpe 2 1,87
coxal 2 1,87
fémur 1 0,93
tibia 3 2,80
fibula 0 0,00
calcaneus 4 3,74
astragale 0 0,00
tarsien 0 0,00
métatarse 2 1,87
phalange 5 4,67
Total 107 100
Fig. 4 : Répartition anatomique chez Bos taurus.
Tableau et schéma anatomique représentant le nombre de restes et le pourcentage de chaque type
d'ossement.
Chez les Ovicaprinés (figure 5), on peut remarquer une plus grande quantité de phalanges. Il
peut s'agir là de déchets de boucherie ou de cuisine, avec découpe et rejet des pieds au dépotoir
(Lignereux et Peters, 1996). Mais cela peut également être dû au traitement particulier dont les os
longs ont pu faire l'objet pour la récupération de la moelle osseuse notamment, et qui aura altéré leur
conservation. Il est à noter par ailleurs que la détermination des côtes, notamment la distinction entre
côtes d'Ovicaprinés et de Suidés effectuée grâce aux atlas ostéologiques et à la collection de
référence, n'a pas pu être systématique, leur état de conservation ne le permettant pas toujours.
Pour ce qui est des Suidés, la représentation des pièces anatomiques est très différente (figure
6). Les pièces de la tête et les extrémités des membres sont les plus représentées. A noter l'absence
des os des membres postérieurs tels que le fémur, le tibia et la fibula. Là encore, il existe plusieurs
hypothèses. La difficile conservation des fémurs, et des ossements de Suidés en général, est à prendre
en considération. Cependant, l'absence d'autres os, en particulier des membres postérieurs des Suidés,
amène à envisager l'hypothèse de l'extraction de ces pièces suite à des découpes bouchères et
charcutières, ceci en vue de consommation ou commercialisation de la viande, fraîche ou en salaison.
9Ce mode de conservation attesté dès l'Age du Bronze sur le littoral de l'Etang de Berre (Benoît,
1965), ainsi que l'évocation de ce procédé dans les sources antiques (Caton, De l'agriculture,
CLXX), rendent cette hypothèse envisageable. Elle nécessite cependant l'appui d'autres éléments,
pour pouvoir être valable. Des recherches dans ce sens devront donc être menées par ailleurs.
Nom des pièces
anatomiques Ovicaprinés (NR) Ovicaprinés (%)
crâne 2 1,53
maxillaire 2 1,53
mandibule 5 3,82
dent 29 22,14
vertèbre 2 1,53
côte 37 28,24
sternum 0 0,00
scapula 3 2,29
humérus 8 6,11
radius 6 4,58
ulna 4 3,05
carpien 1 0,76
métacarpe 7 5,34
coxal 1 0,76
fémur 0 0,00
tibia 3 2,29
fibula 0 0,00
calcaneus 4 3,05
astragale 3 2,29
tarsien 0 0,00
métatarse 4 3,05
phalange 10 7,63
Total 131 100
Fig. 5 : Répartition anatomique chez les Ovicaprinés.
Tableau et schéma anatomique représentant le nombre de restes et le pourcentage de chaque type
d'ossement.
Chez les Lagomorphes, on peut voir une forte représentation des phalanges (figure 7). La
conservation différentielle peut en être une des causes, de même que la récupération de la fourrure
qui implique « une découpe et sélection des extrémités des membres » (Callou, 2003), ou le rejet de
restes culinaires, ces deux dernières possibilités n'étant pas incompatibles entre elles. Les autres
ossements et leur fréquence sont cohérents avec une consommation de la viande, les membres et les
côtes étant les parties les plus riches en viande. Cela pourrait donc appuyer l'hypothèse du double
emploi : récupération de la fourrure et consommation de la viande.
10Nom des pièces
anatomiques Suidés (NR) Suidés (%)
crâne 4 4,08
maxillaire 6 6,12
mandibule 11 11,22
dent 36 36,73
vertèbre 2 2,04
côte 3 3,06
sternum 1 1,02
scapula 2 2,04
humérus 0 0,00
radius 2 2,04
ulna 2 2,04
carpien 0 0,00
métacarpe 14 14,29
coxal 1 1,02
fémur 0 0,00
tibia 0 0,00
fibula 0 0,00
calcaneus 0 0,00
astragale 0 0,00
tarsien 0 0,00
métatarse 6 6,12
phalange 8 8,16
Total 98 100
Fig. 6 : Répartition anatomique chez les Suidés.
Tableau et schéma anatomique représentant le nombre de restes et le pourcentage de chaque type
d'ossement.
Nom des pièces
anatomiques Lagomorphes (NR) Lagomorphes (%)
crâne 0 0,00
maxillaire 0 0,00
mandibule 0 0,00
dent 0 0,00
vertèbre 0 0,00
côte 1 7,69
sternum 0 0,00
scapula 0 0,00
humérus 5 38,46
radius 0 0,00
ulna 0 0,00
carpien 0 0,00
métacarpe 0 0,00
coxal 3 23,08
fémur 1 7,69
tibia 0 0,00
fibula 0 0,00
calcaneus 0 0,00
astragale 0 0,00
tarsien 0 0,00
métatarse 1 7,69
phalange 2 15,38
Total 13 100
Fig. 7 : Répartition anatomique chez les Lagomorphes.
Tableau et schéma anatomique représentant le nombre de restes et le pourcentage de chaque type
d'ossement.
113.3/ Poids de viande
Le calcul du NMI nous a permis d'avoir une meilleure vision de la composition du cheptel
d'animaux consommés sur l'oppidum, en nombre de têtes. Mais qu'en est-il de la quantité de viande
apportée par chaque espèce ?
Pour le savoir, le poids de viande a été estimé selon les données fournies par Columeau (1991
d'après Stouff, 1970), qui impliquent la prise en compte de carcasses éviscérées, sans tête ni pattes.
L'âge d'abattage des animaux n'ayant pas pu être déterminé pour tous les individus, l'estimation du
poids de viande (tableau 3) a été faite en considérant tous les animaux comme des adultes, permettant
ainsi d'obtenir le poids de viande maximal que chaque espèce a pu représenter en fonction du nombre
d'individus déterminés. Il convient cependant de garder à l'esprit qu'il s'agit d'une estimation basée
sur l'hypothèse de carcasses complètes exploitées sur le site. Il reste possible que seules certaines
pièces bouchères aient été apportées sur le site, ce qui pourrait modifier ces résultats.
Tab. 3 : Tableau du poids de viande estimé par espèce.
(PV : Poids de viande estimé en kg, PV : pourcentage du poids de viande total)
Espèce PV % PV
Bos Taurus 5040 65,9
Equidés 180 2,4
Ovicaprinés 629 8,2
Suidés 1680 22,0
Gallus domesticus 3 0,04
Oiseaux de basse-cour 2 0,03
Total animaux domestiques 7534 98,6
Cervidés 100 1,3
Lagomorphes 9 0,1
Total animaux sauvages 109 1,4
Total 7643 100
L'importance de l'élevage sur la chasse se trouve ainsi confirmée et même renforcée, celle-ci
représentant plus de 98 % de l'approvisionnement en viande de l'oppidum. La chasse ne serait donc
qu'un apport visant à améliorer l'ordinaire. Pour ce qui est de l'élevage, le bœuf est la viande la plus
consommée. Celle des Suidés vient ensuite, tandis que les Ovicaprinés représentent moins de 10% de
l'apport en viande. Là encore, ces résultats coïncident avec ceux observés sur les sites contemporains
du pourtour de l'Etang de Berre (Columeau, 1991), quoi que les Suidés soient particulièrement bien
représentés au détriment des Ovicaprinés.
Ces résultats amènent à s'interroger : les Ovicaprinés étaient-ils élevés pour leur viande, ou
bien l'étaient-ils plutôt pour leur lait et leur laine ? Cela signifie-t-il que les habitants avaient une
préférence pour la viande de bœuf et de porc ou que ces viandes étaient privilégiées comme
12offrandes ? Autant de questions qui vont nécessiter un approfondissement des recherches, notamment
sur les pratiques cultuelles de l'époque dans la région, mais également en se penchant plus en détail
sur les quatre phases d'occupation protohistoriques du site ainsi que sur les différents contextes dans
lesquels les ossements furent découverts.
3.4/ Découpe et abattage
Un faible pourcentage du nombre de restes comporte des marques de découpe ou bien des
marques de brûlure (tableau 3). Ils nous apportent cependant des informations importantes.
Tab. 4 : Nombre de restes portant des marques de découpe ou des traces de brûlure.
(NR = nombre de restes, % NR = pourcentage de restes)
NR % NR
NR avec marques de découpe 39 1,8
NR avec traces de brûlure ou brûlés 44 2,0
Ainsi, les traces de brûlure très prononcées sur tout ou partie des fragments concernés
témoignent plus de fragments ou d'os jetés au feu que d'une cuisson, du fait de la localisation étendue
sur toute la surface osseuse des brûlures. Il peut donc être question de restes culinaires dont on se
sera débarrassé dans le foyer, ou de restes issus de rituels liés au sanctuaire voisin.
Pour ce qui est des traces de découpe, elles sont localisées de manière typique d'un travail de
démembrement et de découpage des carcasses. On trouve en effet des stries de désarticulation au
niveau des humérus, des phalanges, ou encore des mandibules dans le cas des Suidés, comme
représentés sur les planches anatomiques suivantes (figures 8, 9 et 10) ainsi que des marques de
décarnisation au niveau des côtes et de désossage (Helmer, 1987 ; Lignereux et Peters, 1996). Un
exemple de ces traces est présenté sur la photo d'humérus d'Ovicapriné (figure 11).
13Fig. 8 : Schéma anatomique représentant les traces de découpe (en rouge) et de fragmentation (en
bleu) observées sur les ossements de Bos taurus.
Fig. 9 : Schéma anatomique représentant les traces de découpe (en rouge) et de fragmentation (en
bleu) observées sur les ossements d'Ovicaprinés.
14Fig. 10 : Schéma anatomique représentant les traces de découpe (en rouge) observées sur les
ossements de Suidés.
Fig. 11 : Traces de découpe (pointées par les flèches) sur un humérus d'Ovicapriné
(photo G. Delpy).
15Ce type de traces n'est pas le seul qu'on peut rencontrer sur les ossements (figures 8, 9 et 10).
Ainsi, le nombre important de fragments osseux découverts sur le site de Constantine peut être, pour
partie, le résultat d'un débitage par fracturation (Averbouh, 2000).
La photo d'un humérus de Bos taurus (figure 12) est un exemple d'os long découpé
longitudinalement tel qu'on peut en trouver sur l'oppidum. Ce type de fracturation par bipartition
(Averbouh, 2000) a été observé sur des humérus, mais également sur des métapodes d'Ovicaprinés et
de Bos taurus. L'hypothèse d'une fracturation ayant comme objectif à la fois la récupération de la
moelle osseuse et l'utilisation des diaphyses en vue de la réalisation de produits de tabletterie pourrait
être envisagée. A noter cependant qu'aucun objet issu de l'industrie osseuse n'a été retrouvé sur le site
durant les fouilles de 2008 et 2009.
Par ailleurs, l'absence d'extrémités des os longs pourrait résulter de la dégradation plus rapide
de ces parties composées majoritairement d'os spongieux, et donc plus fragiles que les diaphyses. Il
peut également s'agir de débitage par segmentation (Averbouh, 2000), également en vue d'un travail
de l'os comme évoqué précédemment.
Fig. 12 : Humérus de Bos taurus coupé en deux (photo G. Delpy).
164/ Conclusions et perspectives
Cette étude préliminaire des restes osseux découverts durant les fouilles de l'oppidum de
Constantine de 2008 et 2009 offre une meilleure vision du spectre faunique présent sur ce site au II e
et Ier s. av. J.-C..
On constate ainsi que l'élevage est l'activité prédominante, tandis que la chasse est une
activité très secondaire qui apporte un complément. La codominance des Ovicaprinés et des Suidés
qu'on peut observer en nombre de têtes s'inscrirait dans un schéma récurrent dans les sites découverts
autour de l'Etang de Berre durant la fin de l'Age du Fer (Columeau 1991, 1996). De même, la
prédominance des Lagomorphes comme gibiers et la rareté des Cervidés sont également observables
sur d'autres sites de la région, tel que l'oppidum de Saint-Blaise (Columeau, 1991). A noter cependant
l'importance non négligeable des bœufs, qu'il s'agisse d'animaux utilisés pour le travail ou comme
offrandes, et qui distingue ce site de ses voisins. Cette prédominance pourrait être liée à la nature
urbaine de Constantine, mais l'âge d'abattage des animaux incite à la prudence. Cet aspect fera donc
partie des points à approfondir, notamment par des comparaisons avec des sites similaires.
Ces résultats sont confirmés par le calcul du poids de viande, quoi que l'importance du
cheptel d'Ovicaprinés se trouve réduite tandis que celle de Bos Taurus est renforcée. L'élevage de
moutons et de chèvres était-il vraiment destiné à la viande ? La question se pose au vu de ces
résultats.
Le traitement boucher des carcasses des animaux d'élevage ainsi qu'une exploitation de leurs
os à des fins de récupération de la moelle est très vraisemblable. La présence du sanctuaire voisin
incite cependant à s'interroger sur le but de cette exploitation animale : commercialisation de viande
destinée à servir d'offrande ? Consommation domestique ou consommation lors de banquets rituels ?
Pour pouvoir répondre à ces questions, ce travail nécessite encore de nombreux
approfondissements, notamment au niveau des pratiques d'élevages propres à chaque espèce animale
domestique présente. La quantité restreinte d'ossements déterminés, et celle plus réduite encore de
restes dont l'âge est estimable grâce à l'épiphysation ou l'usure dentaire, limite cependant une telle
étude. Pour y remédier en partie, l'ajout des restes fauniques de la fouille de 2010 sera d'une aide
précieuse.
La présence de petites fosses creusées à même le substrat calcaire et remplies exclusivement
de restes fauniques nécessitera en outre une étude plus fine qui pourra nous renseigner sur les
coutumes des habitants de l'oppidum.
Ces résultats pourront ensuite être comparés à ceux des sites contemporains de même type
comme l'oppidum de Saint Blaise, comme évoqué précédemment, mais aussi à d'autres sites de
nature très différente. Il sera alors possible de mettre en évidence les différences ou similitudes
17existantes dans leurs spectres fauniques et les méthodes d'approvisionnement en viande et autres
produits d'origine animale, et de vérifier l'hypothèse de particularités régionales voire locales.
Remerciements
Par ces quelques lignes, je tenais à témoigner ma gratitude envers les personnes qui, par leur
aide, contribuent à faciliter mon travail mais aussi de façon plus générale ma thèse actuellement en
cours et intitulée « Saisonnalité et gestion des activités agro-pastorales, cynégétiques et halieutiques
en Gaule Méditerranéenne durant la Protohistoire et l'Antiquité ».
Le Centre Camille Jullian, son directeur D. Garcia (CCJ, CNRS, UMR 6573) et S. Agusta-
Boularot (Université Paul Valéry Montpellier 3, UMR 5140) tout d'abord, pour m'avoir offert ma
chance en me permettant de me lancer dans une thèse d'Archéozoologie. M. Sternberg (CCJ, CNRS,
UMR 6573) pour son aide importante à la conception de mon projet de thèse, ses conseils et son
accueil, mais surtout pour avoir cru en mes possibilités.
F. Verdin (Ausonius, CNRS, UMR 5607) pour m'avoir fait visiter le site et permis d'étudier la
faune de ce site exceptionnel qu'est l'oppidum de Constantine. J.-P. Brugal (Laboratoire
Méditerranéen de Préhistoire Europe Afrique, CNRS, UNR 6636) pour me permettre d'utiliser sa
collection de référence. M. Leguilloux (Centre d'Archéologie du Var) pour son aide et ses conseils.
Et enfin, A. Gardesein (Archéologie des sociétés méditerranéennes, CNRS, UMR 5140) et C.
Chandezon (Université Paul Valéry Montpellier 3, UFR Sciences Humaines et Sciences de
l'Environnement) pour m'avoir permis de présenter mon analyse préliminaire de la faune de
Constantine durant ces Rencontres Archéozoologiques de Lattes 2011.
Bibliographie
18• Aubagnac G., 1990 : « L'enceinte de Constantine (Lançon, B.-du-Rh.) et sa valeur militaire
», dans RAN 23, éditions de l'Association de la RAN, Montpellier, p. 53 à 70.
• Averbouh A., 2000 : Technologie de la matière osseuse travaillée et implications
palethnologiques. L'exemple des chaînes d'exploitation du bois de Cervidé chez les
Magdaléniens des Pyrénées, Thèse de Doctorat de l'Université Préhistoire-Ethnologie-
Anthropologie, Université de Paris I Panthéon-Sorbonne.
• Barone R., 1976 : Anatomie comparée des mammifères domestiques, Tome 1, deux fascicules
: Ostéologie. Vigot Frères, Paris (deuxième édition).
• Benoît F., 1965 : Recherches sur l’Hellénisation du Midi de la Gaule, Nouvelle série n°43,
Ophrys, Aix-en-Provence.
• Boessneck J., 1990 : Exercices archéo-ostéologiques (Archäo-osteologische übungenn)
donné aux doctorants par le Pr. Boessneck, Séminaire d'initiation à l'Archéo-ostéologie
animale, 17-21 avril 1990, par le Pr. A. von den Dreisch, de l'Institut de Paléoanatomie de
Münich à l'Ecole Vétérinaire de Toulouse.
• Bressou C., 1944 : Aide-mémoire d'ostéologie comparée des animaux domestiques, Vigot,
Paris.
• Brun J.-P., 1996 : « La grande transhumance à l'époque romaine. A propos des recherches sur
la Crau d'Arles » dans Anthropozoologica n°24, Paris, p. 31 à 44.
• Callou C., 2003 : De la garenne au clapier. Etude archéozoologique du Lapin en Europe
occidentale, Publications scientifiques du Muséum, Mémoires du Muséum National
d'Histoire Naturelle, Tome 189, Paris.
• Chaix L. et Méniel P., 1996 : Eléments d'archéozoologie, Errance, Paris.
• Columeau P., 1991 : L'animal pour l'homme. Recherches sur l'alimentation carnée dans le
sud de la France du Néolithique au Moyen-Âge d'après les vestiges osseux. I. Le monde
rural, Travaux du Centre Camille Jullian, Université de Provence, Aix-en-Provence.
• Columeau P., 1993 : « Le ravitaillement en viande, la chasse et l'élevage sur les rives de
l'Etang de Berre», dans Archéologie et environnement : de la Sainte-Victoire aux Alpilles,
Publications Université de Provence, Aix-en-Provence, p.301 à 314.
• Columeau P., 1996 : « Pratiques cultuelles et spécialisation pastorale autour de l'Etang de
Berre, de l'âge du Fer à la fin de l'Antiquité » dans L'Etang de Berre, Carte Archéologique de
la Gaule 13/1, Académie des Inscriptions des Belles Lettres, Ministère de la Culture,
Ministère de l'Enseignement Supérieur et de la Recherche avec l'Association pour les Fouilles
Archéologiques Nationales, Fondation Maison des Sciences de l'Homme, Paris, p.128 à 136.
19• Columeau P., 2001 : « Nouveau regard sur la chasse et l'élevage dans le Sud et le Sud-Est de
la Gaule, aux IVe et Ve s. ap. J.-C. et l'exemple de Constantine (B.-du-Rh.) », dans RAN 34,
éditions de l'Association de la RAN, Montpellier, p. 123 à 137.
• Columeau P., 2002 : Alimentation carnée en Gaule du sud (VIIe s. av. J.-C.-XIVe s.),
Publications de l'Université de Provence, Aix-en-Provence.
• Columeau P., 2004 : « Aspects de la consommation de la viande en Provence au cours des
périodes anciennes : les données de l'archéozoologie » dans Provence historique.
Alimentation et cuisine en Provence, tome LIV – fascicule 218, Marseille, p. 413 à 429.
• Gateau F., 1996 : L'Etang de Berre, Carte Archéologique de la Gaule 13/1, Académie des
Inscriptions des Belles Lettres, Ministère de la Culture, Ministère de l'Enseignement
Supérieur et de la Recherche avec l'Association pour les Fouilles Archéologiques Nationales,
Fondation Maison des Sciences de l'Homme, Paris.
• Helmer D., 1987 : Fiches descriptives pour les relevés d'ensembles osseux d'animaux,
Fiches d'Ostéologie animale pour l'Archéologie, Série B (1), CRA/CNRS, Valbonne.
• Helmer D., 2000 : « Discrimination des genres Ovis et Capra à l'aide des prémolaires
inférieures 3 et 4 et interprétation des âges d'abattage : l'exemple de Dikili Tash (Grèce) »
dans Anthropozoologica n°31, p. 29 à 38.
• Lignereux Y. et Peters J., 1996 : « Techniques de boucherie et rejets osseux en Gaule
Romaine » dans Anthropozoologica n°24, Paris, p. 45 à 98.
• Méniel P., 1987 : Chasse et élevage chez les Gaulois, Errance, Paris.
• Pavaux C., 1976 : Ostéologie des mammifères domestiques, Document pédagogique de
l'Ecole Vétérinaire de Toulouse, 3 fascicules.
• Poplin F., 1976 : « A propos du nombre des restes et du nombre d'individus dans les
échantillons d'ossements », dans Cahiers du Centre de Recherches Préhistoriques, Paris I, 5,
p. 61 à 74.
• Poplin F., 1981 : « Un problème d'ostéologie quantitative : calcul d'effectif initial d'après
appariement. Généralisation aux autres types de remontages et à d'autres matériels
archéologiques », dans Revue d'archéométrie, Actes du XX symposium international
d'archéométrie 26 – 29 mars 1980, Volume II, Paris.
• Schmid E., 1972 : Atlas of animal bones for prehistorians archaeologists and quaternary
geologists, Elvesier Publishing Company, Amsterdam, Oxford, New York.
• Stouff L., 1970 : Ravitaillement et alimentation en Provence au XIVe et XVe siècles., Mouton
et Co., Paris-La Haye.
20• Trément F., 1999 : Archéologie du paysage. Les étangs de Saint-Blaise (Bouches-du-Rhône),
Editions de la Maison des Sciences de l'Homme, DAF 74, Paris.
• Verdin F., Bouet A. et Aubagnac G., 1992 : «Lançon, Constantine», dans Bilan Scientifique
1991, SRA-PACA, Aix-en-Provence, p. 110-111.
• Verdin F., 1993 : « L'occupation protohistorique du chaînon de la Fare. Recherches en cours
», dans Archéologie et environnement : de la Sainte-Victoire aux Alpilles, Publications
Université de Provence, Aix-en-Provence, p.137 à 147.
• Verdin F., 2001 : « L'oppidum de Constantine (Lançon-de-Provence) : un exemple
d'établissement de hauteur réoccupé durant l'Antiquité tardive », dans RAN 34, éditions de
l'Association de la RAN, Montpellier, p. 105 à 121.
• Verdin F., 2005 : « Lançon-Provence. Oppidum de Constantine », dans Bilan scientifique
2004, SRA-PACA, Aix-en-Provence, p.154.
• Verdin F., 2008 : Oppidum de Constantine, Fouille programmée 13 juillet – 3 août 2008,
Service Régional de l'Archéologie de PACA.
• Verdin F., 2009 : Oppidum de Constantine, Fouille programmée 12 juillet – 2 août 2009,
Service Régional de l'Archéologie de PACA.
• Von den Driesch A., 1976 : A guide to the measurement of animal bones from Archaeological
Sites, Peabody Museum Bulletins, 1, Harvard University.
21Vous pouvez aussi lire

















































