Université Pierre & Marie Curie Paris 6 - Jussieu
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Habilitation à Diriger des Recherches | Simon Scheuring
Université Pierre & Marie Curie
Paris 6 - Jussieu
Mémoire
Présenté pour l’obtention du diplôme
d’Habilitation à Diriger des Recherches
Spécialité: Science de la Vie et de la Santé
par
Simon Andreas Scheuring
Simon Scheuring
Institut Curie
UMR-CNRS 168
11 rue Pierre et Marie Curie
75231 Paris Cedex 05
France
simon.scheuring@curie.fr
Tel.: ++33 1 42 34 67 75
Fax.: ++33 1 40 51 06 36
Paris – Août 2005
1/53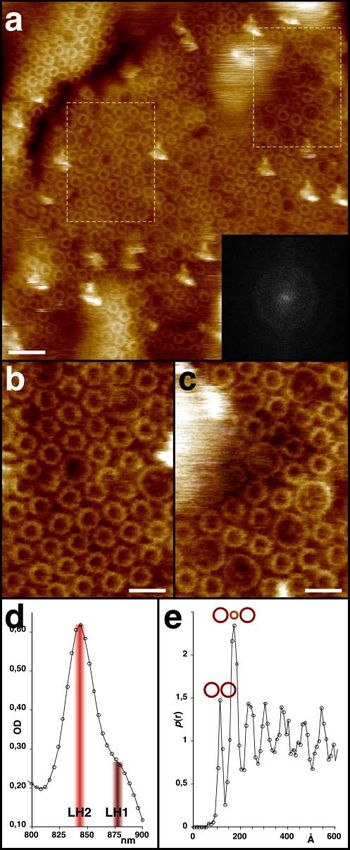
Habilitation à Diriger des Recherches | Simon Scheuring
2/53
Habilitation à Diriger des Recherches | Simon Scheuring
“Human subtlety will never devise an invention more beautiful, more
simple or more direct than does Nature, because in her inventions,
nothing is lacking and nothing is superfluous.”
Leonardo da Vinci (1452-1519)
painter, sculptor, anatomist, architect, engineer, inventor, geometer, philosopher
3/53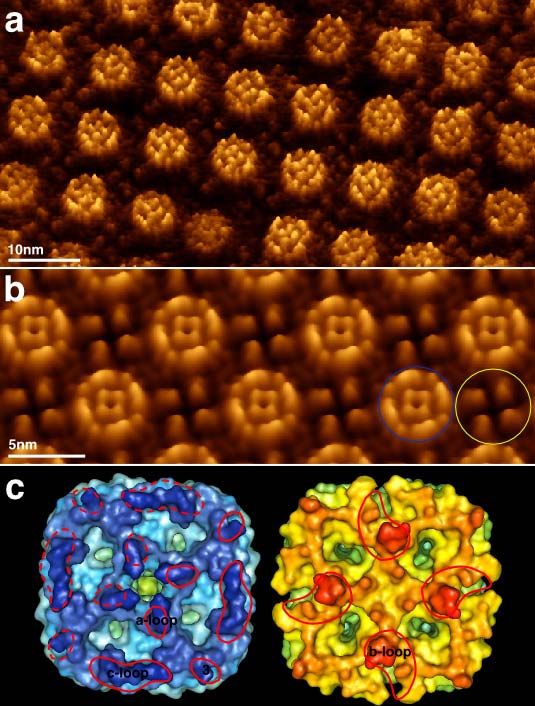
Habilitation à Diriger des Recherches | Simon Scheuring
4/53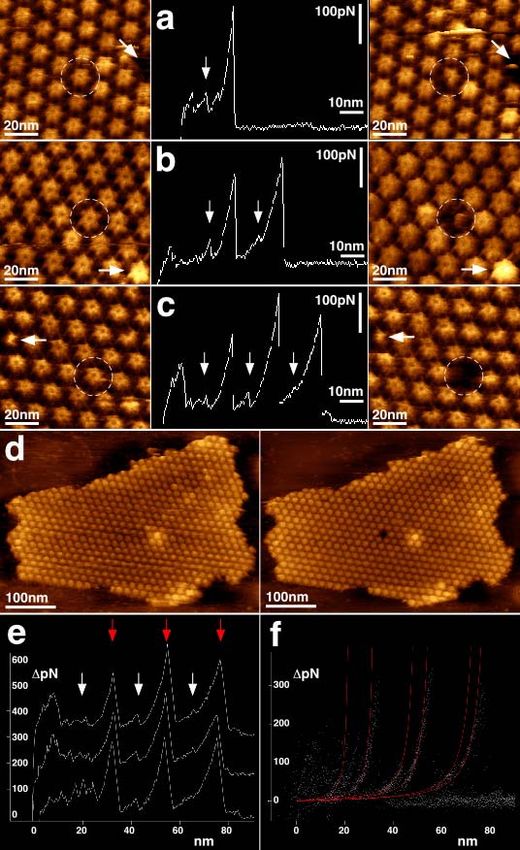
Habilitation à Diriger des Recherches | Simon Scheuring
INDEX:
1. RÉSUMÉ (FRANÇAIS) ................................................................................................................... 6
2. ABSTRACT (ENGLISH) ................................................................................................................. 7
3. INTRODUCTION............................................................................................................................. 8
3.1. LES PROTÉINES MEMBRANAIRES ET LEUR ANALYSE STRUCTURALE .......................................... 8
3.2. LE MICROSCOPE À FORCE ATOMIQUE – UN OUTIL PUISSANT POUR LA RECHERCHE SUR LES
PROTÉINES MEMBRANAIRES ...................................................................................................... 10
4. RAPPORT SCIENTIFIQUE ......................................................................................................... 12
4.1. ASPECTS TECHNIQUES DE L’ AFM ............................................................................................ 12
4.2. ASPECTS EXPÉRIMENTAUX ET RATIONNELS DU MICROSCOPE À FORCE ATOMIQUE (AFM)..... 12
4.3. IMAGERIE PAR MICROSCOPIE À FORCE ATOMIQUE (AFM) ....................................................... 13
4.4. APPLICATION ............................................................................................................................. 15
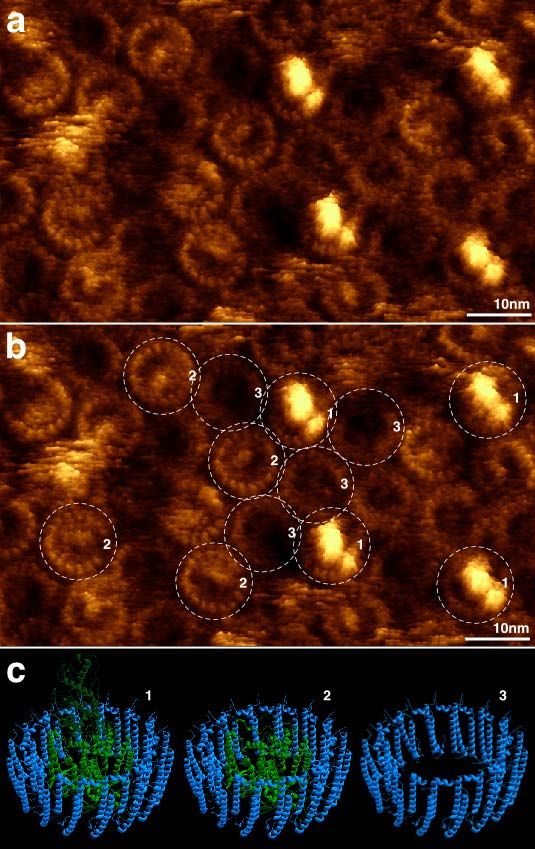
4.4.1. Imagerie à haute résolution – détection de contours de boucles individuelles .................. 15
4.4.2. Identification de domaines protéique par imagerie AFM et protéolyse.............................. 18
4.4.3. Imagerie et nano-manipulation - la pointe d'AFM comme un nano-outil........................... 19
4.4.4. Imagerie et mesure de force – le AFM en tant que sonde de force. .................................... 22
4.4.5. Imagerie à haute résolution – architecture supra-moléculaire en membrane native......... 24
4.4.6. Imagerie à haute résolution de super-complexes en membranes natives ........................... 29
5. PROJET............................................................................................................................................ 32
5.1. IMAGERIE À HAUTE RÉSOLUTION DES MEMBRANES NATIVES ET ÉLUCIDATION DES
ASSEMBLAGES PROTÉIQUES SUPRA-MOLÉCULAIRES DANS CELLES-CI. ................................... 33
5.2. COMBINAISON DE L'IMAGERIE À HAUTE RÉSOLUTION AVEC LES MESURES DE SPECTROSCOPIE
DE FORCE POUR ÉLUCIDER LES FORCES INTER- ET INTRA-MOLÉCULAIRES ENTRE ET À
L'INTÉRIEUR DE PROTÉINES MEMBRANAIRES. .......................................................................... 34
5.3. IMAGERIE DE CHANGEMENTS DE CONFORMATIONS FONCTIONNELLEMENT PERTINENTS DE
PROTÉINES MEMBRANAIRE. ...................................................................................................... 35
5.4. DÉVELOPPEMENT DE NOUVEAUX SUPPORTS AFM POUR L'IMAGERIE DE PROTÉINES
MEMBRANAIRES FONCTIONNELLES, DANS UNE MEMBRANE NON SUPPORTÉE SÉPARANT DEUX
COMPARTIMENT AQUEUX.......................................................................................................... 36
6. CURRICULUM VITAE................................................................................................................. 38
6.1. EDUCATION ................................................................................................................................ 38
6.2. TEACHING .................................................................................................................................. 38
6.3. TRAINEES ................................................................................................................................... 39
6.4. PUBLICATIONS ........................................................................................................................... 40
6.4.1. Publications – Research Articles .......................................................................................... 40
6.4.2. Publications – Review Articles.............................................................................................. 42
6.4.3. Publications – Didactic Writings.......................................................................................... 42
6.5. VISITING SCIENTIST AND SCIENTIFIC SCHOOLS ....................................................................... 42
6.6. MEETINGS AND INVITED PRESENTATIONS ................................................................................ 43
6.7. REVIEWER .................................................................................................................................. 46
6.8. EDITORIAL BOARD MEMBER ..................................................................................................... 46
7. ANNEXE 1 - REMERCIEMENTS............................................................................................... 47
8. ANNEXE 2 – REFERENCES........................................................................................................ 48
5/53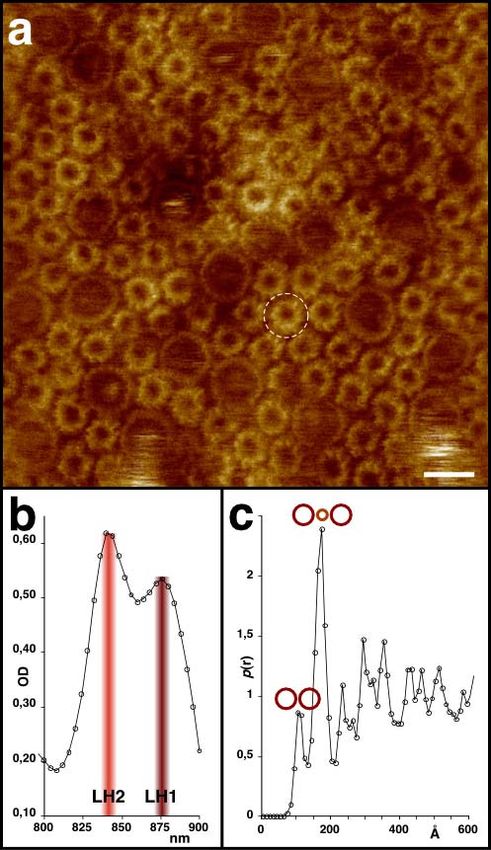
Habilitation à Diriger des Recherches | Simon Scheuring
1. Résumé (Français) dans lesquels la pointe de l’AFM est
utilisée pour analyser une couche entière
Avec ce rapport d’HDR, je montre
de protéines, des sous unités de protéine
que j’ai réussi à obtenir des résultats
unique provenant d’un complexe
scientifiques nouveaux et significatifs et protéique, ou des boucles individuelles
que je suis capable de planifier et
de surface.
poursuivre un projet de recherche
4. l’AFM comme outil pour mesurer des
original de façon indépendante. Ce
forces : Dans cette sous-section, les
rapport comporte deux axes majeurs: (i)
possibilités d’utilisation de l’AFM pour
un rapport scientifique qui contient les des mesures de forces afin d’élucider les
principaux résultats que j’ai obtenus, (ii)
forces inter- et intramoléculaires sur des
une description de projets dans la quelle
protéines individuelles sont démontrées
je présente mes plans de recherche pour et discutées.
le futur.
5. l’AFM comme seul outil possible pour
(i) J’ai décidé de présenter le
étudier, à haute résolution, les
rapport scientifique en termes de
membranes natives non cristallines
possibilités techniques plutôt qu’en
contenant de multiples complexes de
termes de résultats obtenus. Après avoir protéines différentes: Cette recherche a
introduit ce que sont les protéines
complété nos connaissances sur la
membranaires et la technique de
photosynthèse bactérienne du point de
microscopie à force atomique (AFM) de vu de l’architecture supramoléculaire des
façon générale, la section développant
membranes natives.
les résultats de recherche obtenus est
(ii) J’ai divisé le projet de
divisée est cinq points représentant les
recherche future en quatre points qui
différentes applications de l’AFM.
diffèrent en termes d’approche technique
1. l’AFM comme technique d’imagerie mais dont les contenus se chevauchent
pure qui permet d’acquérir des
d’un point de vue pertinence biologique.
informations structurales sur des boucles
1. le point le plus important est le
individuelles de protéines présentes à la développement de l’AFM pour
surface de membrane : Dans cette sous-
l’imagerie des membranes natives de
section, je me concentre sur le travail
plus en plus complexes.
effectué sur AqpZ. Je compare la
2. pour combiner l’imagerie et la
topographie obtenue par AFM (1999)
spectroscopie de force par AFM. Cet
avec la structure obtenue par rayons X
aspect peut objectivement être intégré au
(2003), ce qui permet d’évaluer
premier point de façon à accéder aux
rétrospectivement la fiabilité de
forces intramoléculaires entre complexes
l’imagerie par AFM et l’interprétation protéiques dans des systèmes natifs.
des données de façon complètement non
3. l’imagerie sera utilisée pour accéder
biaisée.
aux changements de conformation de
2. en combinant l’imagerie par AFM protéines membranaires, liés à leurs
avec le clivage protéolytique pour
fonctions.
assigner les côtés cytoplasmique et
4. en relation avec le troisième point, de
périplasmique des protéines
nouveaux supports d’AFM seront
membranaires et pour déterminer la
développés pour être capable d’imager
localisation des domaines peptidiques. des membranes non-supportées séparant
3. l’AFM comme un nano-outil pour
deux chambres aqueuses. De tels
manipuler des objets biologiques: Dans
supports permettront d’observer la
cette sous-section, je discute l’utilisation structure de protéines membranaires
de la pointe d’AFM comme un nano-
ainsi que leurs changements de
outil pour manipuler la structure des
conformation liés à leurs fonctions dans
protéines en appliquant des forces sur le
un état aussi proche que possible de
cantilever. Trois exemples sont donnés
l’état natif.
6/53
Habilitation à Diriger des Recherches | Simon Scheuring
2. Abstract (English) individual surface loops are displaced.
Forth, the AFM as a force measure
With this HDR report, I give
tool: In this subtopic, the possibilities of
account that I have achieved significant
using the AFM for force measurements
and novel scientific results and that I am to elucidate the inter- and intra-
able to plan and pursue an independent
molecular forces on individual proteins
original research project.
is demonstrated and discussed.
This report has two major axes: (i)
Fifth, the AFM as a unique tool to
a scientific report in which I describe
study non-crystalline native membranes
some of the major achieved scientific containing multiple different protein
result, and (ii) a project description in
complexes at high-resolution: This
which I present my future research plans.
research has complemented the field of
bacterial photosynthesis with structural
(i) I have decided to present the
views on the super-molecular
scientific report in terms of technical
architecture within native membranes.
possibilities rather than in terms of
results achieved. After introducing
(ii) I have divided the future
membrane proteins and the technique research project into four subtopics,
atomic force microscopy (AFM) in
differing in terms of the technical
general, the section documenting the
approach but with some overlap
research results is divided in five concerning the biological relevance.
subtopics representing different
First, and of major importance, the
applications of the AFM.
development of the AFM for imaging
First, the AFM as a pure imaging
more and more complex native
technique to acquire structural
membranes.
information on individual loops on Second, to combine imaging and
membrane protein surfaces: In this
force spectroscopy measurements by
subtopic I focus on the work performed
AFM. This aspect can obviously be
on AqpZ; I compare the AFM integrated with the first subtopic, in
topography (1999) with the X-ray
order to assess the intramolecular forces
structure (2003) allowing to
between protein complexes in native
retrospectively evaluate AFM imaging
systems.
reliability and data interpretation
Third, imaging will be used to
completely unbiased.
assess functional related conformational
Second, combining AFM imaging
changes of membrane proteins.
with proteolytic cleavage to assign
Fourth, and closely related to the
membrane protein sidedness and the third subtopic, novel AFM supports will
localization of peptide domains.
be developed in order to be able to
Third, the AFM as a nano-tool to
image non-supported membranes
manipulate biological objects: In this separating two aqueous chambers. Such
subtopic, I discuss the use of the AFM
supports will allow observing membrane
tip as a nano-tool to manipulate protein
protein structure and functional related
structure by employing loading forces to
conformational changes in an as close to
the cantilever. Three examples are given
native state as possible.
in which the AFM tip is used to dissect
an entire protein layer, single protein
subunits from a protein complex, or
7/53
Habilitation à Diriger des Recherches | Simon Scheuring
3. Introduction hautement hydrophobes, qui sont le plus
probablement intégrées dans des
Dans ce rapport d’HDR, je vais
membranes de cellules (Wallin and von
détailler mes contributions et mes succès
Heijne, 1998).
dans le développement et l’utilisation du
microscope à force atomique (AFM)
comme outil pour étudier les protéines
membranaires. De plus, je présenterai les
futures directions dans lesquelles je
souhaite axer mes recherches en utilisant
l’AFM pour obtenir de nouvelles
informations sur les protéines
membranaires et leur assemblée.
3.1. Les protéines membranaires et Fig 1) Abondance (axe-Y) en fonction de
leur analyse structurale l’hydrophobicité (axe- X) de produits de gènes
Les protéines membranaires des génomes de E. Coli, M. jannaschii, and H.
sapiens. Les protéines hautement hydrophobes (à
intrinsèques ou intégrales sont définies droite du graphe) représentent les protéines
comme des protéines qui pénètrent, ou, membranaires.
la plupart du temps, traversent la
bicouche lipidique de membranes Ce résultat intriguant souligne
biologiques. Les structures des protéines, l’extrême importance des protéines
qui se cloisonnent dans des lipides plutôt membranaires pour les êtres vivants. Les
que de rester en solution, ont des protéines membranaires font la
propriétés chimiques spécifiques. Elles connexion entre le cytoplasme et
sont riches en acides aminés l’espace extracellulaire de chaque
hydrophobes qui sont exposés et sont cellule, forment des jonctions entre
restreintes à leur structure secondaire. cellules ou jouent un rôle important dans
Une conséquence de ces propriétés les compartiments intracellulaires. C’est
physicochimiques est qu’une protéine pourquoi, dans les bactéries, de telles
membranaire intégrale ne peut être mise molécules sont impliquées dans le
en solution qu’en étant solubilisée en transport, la sécrétion et les processus
présence de détergents. C’est pour cette bioénergétiques. Les organismes
raison que les protéines membranaires pluricellulaires nécessitent aussi la
sont difficiles à étudier. communication entre les cellules qui les
Les génomes de plusieurs constituent. En conséquence, un grand
organismes ont été criblés pour connaître nombre de protéines membranaires ont
les protéines codées par les cadres de évolué, fonctionnant comme récepteurs
lecture ouverts (« open reading frames » dans le trafic intracellulaire ou dans
ORFs), et ces cadres de lecture ouverts l’adhésion cellulaire, et dans la
ont été traduits en séquences d’acides reconnaissance. L’évolution a aussi créé
aminés. La discrimination des peptides des canaux et des transporteurs
s’est faite en fonction de leur hautement spécifiques qui sont essentiels
hydrophobicité (Fig. 1). Ceci a conduit à pour la survie des systèmes biologiques.
deux familles de produits: les protéines Aussi, la suppression de plusieurs de ces
cytoplasmiques hydrophiles et les protéines est mortelle ou conduit à de
protéines membranaires qui contiennent graves maladies (Fig. 2).
de larges domaines hydrophobes (par Cependant, en dépit des
exemple représentant des hélices trans- informations obtenues par les techniques
membranaires). Par cette simple de biochimie, biophysique et biologie
approche, il a été démontré que 20 - 30 moléculaire, notre compréhension des
% des gènes codent pour des protéines phénomènes membranaires est largement
8/53
Habilitation à Diriger des Recherches | Simon Scheuring
limitée en manque d’information révèlent des traits qui sont comparés à
structurale. En effet, en comparaison des volumes calculés avec une résolution
avec les protéines solubles, seulement de l’ordre de 10 Å à partir de structures à
quelques structures de protéines très haute résolution. D’autres
membranaires ont été résolues par limitations viennent du fait que les
cristallographie par rayons-X et chevauchements de domaines
électrons (http://www.rcsb.org/pdb/; membranaires qui sont encore encastrés
http://www.mpibp-frankfurt.mpg.de/ dans des micelles de détergent sont
michel/public/memprotstruct.html). souvent très mal résolus dans les
reconstructions en ME de particule
isolée. De plus, le rapport signal sur bruit
en ME est si faible que la taille des
molécules est critique pour un
alignement ultérieur des particules. Quoi
qu’il en soit, cette technique peut fournir
Fig 2) Schéma partiel représentant les fonctions des informations structurales importantes
de protéines membranaires. Les classes de sur les supercomplexes. Les cartes de
canaux et de protéines membranaires faisant densités de tels complexes qui ont une
intervenir des processus bioénergétiques sont
omises. résolution moyenne peuvent être
superposées aux structures obtenues par
Les informations structurales sur rayons-X avec une très haute résolution
les protéines membranaires peuvent être (Bibby et al., 2001; Dudkina et al.,
obtenues en utilisant différentes 2005). (ii) L’analyse par cristallisation à
techniques dont les avantages et les 2D peut, dans le meilleur des cas, aboutir
limitations vont être brièvement à un modèle atomique d’une protéine.
discutées maintenant : Les cartes de densités obtenues par
- la microscopie électronique (ME): La cristallographie en ME avec une
microscopie électronique de protéines résolution de 4 Å semble être suffisante
membranaires peut être réalisées soit par pour tracer un modèle structural
analyse de particules uniques de raisonnable (Henderson et al., 1990;
protéines membranaires solubilisées, soit Kühlbrandt et al., 1994; Murata et al.,
par cristallisation à 2D de protéines 2000). En outre, la projection à 2D et les
membranaires reconstituées dans des cartes de densités à 3D à toutes les
bicouches lipidiques. (i) L’analyse de résolutions intermédiaires peuvent être
particules uniques peut, en principe, obtenues, donnant éventuellement des
toujours être réalisée, même avec de très réponses importantes sur la
petites quantités (de l’ordre du stochiométrie et le regroupement en
microgramme). Le principe est basé sur hélices. Un facteur limitant est la
le fait que les molécules individuelles croissance de cristaux 2D. En effet, bien
sont dispersées sur les grilles de que certaines règles aient été
microscopie électronique avec une découvertes, les détails de la
orientation aléatoire et qu’un volume à cristallisation à 2D restent énigmatiques.
3D peut être calculé à partir de toutes les La quantité de protéines nécessaire pour
différentes vues en projection (Frank et réaliser correctement cette analyse est de
al., 1987; Frank et al., 1996). Cette l’ordre du milligramme (Engel et al.,
approche est jusqu’à maintenant 1992; Kühlbrandt, 1992; Rigaud et al.,
fortement limitée en termes de résolution 2000).
obtenue (bien que la communauté - cristallographie par rayons X: Un
scientifique ne se soit même pas encore faisceau de rayon X est diffracté sur un
mise d’accord sur un seul critère de cristal à 3D qui a poussé à partir de
résolution!). Les meilleurs volumes à 3D protéines membranaires. Les figures de
diffraction obtenues sont ensuite
9/53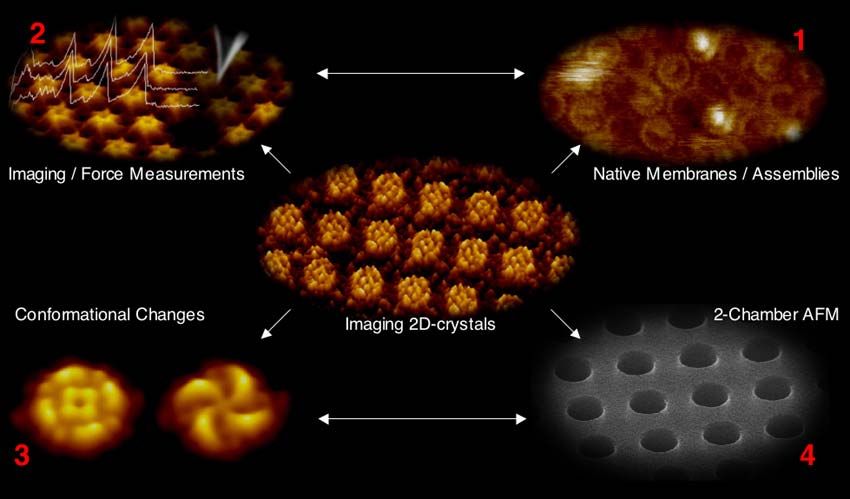
Habilitation à Diriger des Recherches | Simon Scheuring
interprétées. La cristallographie par jours, à une température élevée.
rayons-X a permis d’obtenir la plupart Toutes ces considérations
des structures connues de protéines témoignent des énormes besoins
membranaires. Les meilleurs modèles nécessaires aux techniques d’analyses
sont basés sur des données obtenues structurales alternatives et
avec une résolution inférieure à 2 Å. Les complémentaires pour acquérir des
facteurs limitant sont les suivants : informations sur les protéines
Premièrement, la quantité de protéines membranaires. L’AFM représente une de
nécessaires dans cette approche est ces alternatives. En outre, je vais
relativement grande (quelque dizaine de montrer que, à cette date, l’AFM est le
milligrammes). Ceci limite en particulier seul outil capable d’accéder aux
l’élucidation de protéines eucaryotes arrangements supramoléculaires dans les
dont la surexpression est critique. membranes natives avec une résolution
Deuxièmement, le facteur le plus de l’ordre du nanomètre.
limitant est la difficulté de faire croître
des cristaux à 3D cohérents. Encore une 3.2. Le microscope à force atomique
fois, bien que quelques règles aient été – un outil puissant pour la
identifiées, ce processus reste largement recherche sur les protéines
empirique. Troisièmement, l’information membranaires
sur la phase n’est pas inclue dans les Le microscope à force atomique
spots de diffraction ; elle doit donc être (AFM; schématisé dans la figure 3);
acquise par des artifices secondaires en (Binnig et al., 1986) permet de faire des
utilisant par exemple des cristaux à 3D images de protéines membranaires avec
contenant des métaux lourds, ou en une résolution latérale supérieure à 10 Å
déterminant la phase si la structure est et latérale de l’ordre de 1 Å. La haute
déjà partiellement connue. Maintenant, résolution de l’AFM peut directement
les chercheurs qui effectuent de la donner des informations sur
cristallographie par rayons-X ont a leur l’organisation de domaines extra-
disposition de multiples techniques membranaires comme des boucles
(goûte assise, goûte suspendue, phase reliant des hélices, sur l’insertion des
lipidique cubique) qui permettent protéines membranaires dans la bicouche
d’améliorer leur taux de réussite en lipidique et enfin sur les changements de
cristallisation (Caffrey, 2003). conformation (Müller et al., 1999b;
- la résonance magnétique nucléaire Schabert et al., 1995; Scheuring et al.,
(RMN). La RMN est une technique très 2001a; Scheuring et al., 2003a). La
puissante pour la biologie structurale, combinaison de l’imagerie par AFM
mais qui, jusqu’à maintenant et avec les expériences de protéolyse
probablement dans le futur proche aussi, permet l’identification de domaines
est considérée comme inappropriée pour protéiques et la définition des deux côtés
l’analyse structurale de protéines des protéines membranaires (Scheuring
membranaires. Même si quelques succès et al., 1999b; Scheuring et al., 2001b).
ont été reportés sur la structure de la En appliquant des forces
chaîne des C-alphas de petits beta-barrel supplémentaires à la pointe de l’AFM
porins (Fernandez et al., 2004), aucun durant le processus d’imagerie à haute
résultat n’a encore été publié sur résolution, il est possible de nano-
l’élucidation de la structure d’une large manipuler des objets biologiques. Ce
protéine membranaire hélicale. La RMN “mode” peut être utiliser pour disséquer
nécessite de grandes quantités de des membranes entières ou des sous
protéines qui doivent être marquées par unités individuelles de complexes
des isotopes. De plus, l’échantillon ne multiprotéiques, ou bien pour déplacer
doit pas être affecté durant une durée de façon non destructive des boucles
allant de plusieurs heures à plusieurs
10/53Habilitation à Diriger des Recherches | Simon Scheuring
individuelles à la surface d’une protéine permet de corréler les mesures de forces
membranaire (Müller et al., 1995; avec des changements topographiques
Scheuring et al., 1999b; Scheuring et al., (structurals). De cette façon, les forces
2002b; Scheuring et al., 2003b; inter- et intramoléculaires peuvent être
Scheuring et al., 2004c). déterminées (Müller et al., 1999a;
Oesterhelt et al., 2000; Scheuring et al.,
2002b).
Plus récemment, il a été montré
que l’imagerie à haute résolution peut
être réalisée sur des membranes natives
contenant une grande variété de
protéines (Bahatyrova et al., 2004;
Scheuring et al., 2003b; Scheuring et al.,
2004b; Scheuring et al., 2004c;
Scheuring and Sturgis, 2005). Ceci
permet, pour la première fois, de donner
un aperçu de l’assemblée supra-
moléculaire de protéines membranaires
dans un système natif avec une
résolution de l’ordre du nanomètre.
Les développements techniques de
Fig. 3) Schéma d’un AFM. Une pointe est fixée l’AFM biologiques sont très
sur un cantilever mou. En dessous, l’échantillon prometteurs: il existent des prototypes
est scanné dans les directions X, Y et Z grâce à d’AFM qui permettent l’acquisition
un piézoélectrique. La déflection du cantilever,
due aux ondulations de la surface, est enregistrée
d’images avec une vitesse et donc une
par l’intermédiaire d’un laser qui est focalisé sur résolution temporelle comparable à celle
la face arrière de la pointe. Le signal de de la vidéo, des prototypes avec des
déflection est injecté dans un système de boucle cantilevers courts qui sont considérable-
de rétroaction qui compense la déflection par un ment plus sensibles car les constantes de
mouvement du piézo. La mise en commun des
positions en X et en Y, ainsi que le déplacement
raideur sont plus faibles et le bruit
en Z permet de décrire la topographie de thermique est diminué. Enfin, de
l’échantillon. nouveaux supports biocompatibles sont
construits. Une meilleure compréhension
L’AFM peut aussi être utilisé pour du mécanisme d’imagerie de contact et
effectuer des mesures de forces des modes d’imagerie en l’absence de
permettant de remonter à des forces contact permanent dans les liquides va
inter- et intramoléculaire avec une renforcer l’utilité de l’AFM comme outil
sensibilité inférieure à 10 pN (Moy et d’investigation des systèmes biologiques
al., 1994; Rief et al., 1997a). L’imagerie natifs.
à haute résolution et les mesures de
forces sur des molécules individuelles
peuvent être combinées. Cette approche
11/53Habilitation à Diriger des Recherches | Simon Scheuring
4. Rapport scientifique balayage de l’échantillon placé en
dessous du stylet (attaché au cantilever
4.1. Aspects techniques de l’ AFM
flexible) pendant que le piézo déplace
Les éléments clés d’un AFM, l’échantillon verticalement pour garder
représentés schématiquement dans la la déflection du cantilever constante. Le
figure 4, sont : le cantilever avec un système optique permet de résoudre des
stylet de forme pyramidale qui touche déflections de l’ordre de 0,1 nm, ce qui
l’échantillon, un système optique qui est correspond à des forces variant
constitué d’un laser et d’une photodiode typiquement de 10 à 50 pN. Avec des
multi quadrants (qui permet de détecter instruments modernes, il est possible
les déflections du cantilever), un appareil d’utiliser un mode de contact stable avec
piézo-électrique qui translate l’échan- des forces de l’ordre de 100 pN, à
tillon de façon relative par rapport à la condition que l’échantillon soit en
pointe dans les directions X, Y et Z, et solution (Müller et al., 1999b).
un ordinateur qui pilote le microscope et
enregistre les contours des surfaces (Fig. 4.2. Aspects expérimentaux et
4a). rationnels du microscope à force
atomique (AFM)
L’utilisation de l’AFM en mode de
contact induit de la friction ; les
échantillons ont besoin de bien adhérer
sur des surfaces de mica fraîchement
clivées. Autrement, les forces de friction
peuvent être minimisées en faisant
osciller le cantilever verticalement. En
Fig. 4) a) Un faisceau laser est réfléchi sur la utilisant ce « mode oscillant », des
face arrière du cantilever de l’AFM, permettant échantillons peu adsorbés peuvent être
d’enregistrer la position relative du cantilever sur
un détecteur multi-quadrants. Le signal est traité
observés bien que la résolution soit
par le contrôleur qui pilote le piézo dans les moins bonne (Hansma et al., 1994;
directions X et Y, et compense la déflection du Schabert and Rabe, 1996).
cantilever par un déplacement dans la direction Z Il a été montré, en utilisant
de l’échantillon. Cette compensation de la différent échantillons biologiques avec
déflection de la pointe, qui influence
alternativement cette déflection à chaque
différentes densités de charges
position X-Y, est appelée la boucle de surfaciques (Butt, 1991b, 1992b;
rétroaction. L’utilisateur interagit avec le Israelachvili, 1991), que l’adsorption de
contrôleur via un ordinateur, il peut changer les l’échantillon est contrôlée par la nature
paramètres d’acquisition et lire les mouvements et la concentration en électrolytes dans le
du piézo correspondant à l’image de et par le
contrôleur. Plus la boucle de rétroaction réagit
tampon (Alshakhshir et al., 1995; Müller
vite, plus la détection du contour de la surface est et al., 1997; Roth and Lenhoff, 1993).
précise. b) Gros plan sur la cellule fluide (fluid J’ai contribué au processus d’établisse-
cell). L’échantillon est déposé sur du mica qui ment des conditions d’imagerie sur
est collé sur une plaque de téflon (qui protège le différents échantillons avec différentes
piézo), elle-même collée sur une plaque
magnétique. La pointe et l’échantillon se
charges de surface. De façon
trouvent de façon permanente en solution lors intéressante, les interactions entre le
des mesures. stylet et l’échantillon impliquent, dans
une large mesure, les mêmes forces que
L’instrument est utilisé à celles qui sont engagées entre
température ambiante avec des solutions l’échantillon et le support (des forces
tampons dont la composition peut être électrostatiques et de Van der Waals), et
changée au cours de l’expérience (Fig. qui peuvent être contrôlées en changeant
3b). Un topographe est enregistré par les conditions de tampon (Müller et al.,
12/53Habilitation à Diriger des Recherches | Simon Scheuring
1999b). L’immobilisation sur du mica, forces de van der Waals attractives
ou sur d’autres surfaces chargées, (Müller et al., 1999b; Scheuring et al.,
nécessite ainsi des conditions de tampon 2001a). Ces forces peuvent être décrites
différentes par rapport à celles de de façon quantitative, ce qui permet
l’imagerie. Pour l’adsorption, la d’optimiser les conditions d’enregistre-
longueur de Debye doit être minimisée De façon alternative, pour des
pour permettre aux forces attractives de échantillons hydrophobes comme les
Van der Waals de prédominer (Müller et cristaux à 2D qui ont poussé sur des
al., 1997) (Fig. 5). monocouches lipidiques, j’ai développé
une autre méthode de préparation qui
utilise des supports hydrophobes. Dans
ce cas, du graphite pyrolitique hautement
orienté (highly oriented pyrolytic
graphite HOPG) peut être utilisé comme
support, avec la limitation que le HOPG
n’est pas très plat sur des grandes
distances mais expose des terrasses
planes d’une taille maximale de l’ordre
de quelques microns (Scheuring et al.,
1999a).
Des progrès significatifs ont été
effectués dans beaucoup de laboratoires
Fig. 5) a) Représentation schématique de pour optimiser la préparation des
l’interaction entre la pointe de l’AFM et échantillons (supports d’échantillon
l’échantillon. La pointe pyramidale interagit avec stables, composition des tampons pour
un large étendu de l’échantillon par des forces
électrostatiques répulsives à longues distances. contrôler les interactions échantillon-
En même temps, le sommet de la pointe interagit substrat et échantillon-stylet), ainsi que
à très courtes distances avec l’échantillon. b ) les conditions d’imagerie d’objet de
Représentation vectorielle des contributions des matière molle biologique (courbes force-
forces à longues distances répulsives distance pour ajuster la force appliquée
électrostatiques et des forces à courtes distances
attractives de van der Waals qui se cumulent à la au cantilever inférieure à 100 pN,
force appliquée par la pointe. Dans le cas idéal, calibration de la vitesse de balayage et
la force appliquée ne doit pas excéder ~100 pN et des paramètres de la boucle de
doit s’annuler avec les autres forces présentes. c) rétroaction).
Représentation schématique des courbes force-
distance obtenues sur des objets biologiques. En
haut: Courbe de force dans le cas où les forces à 4.3. Imagerie par microscopie à force
longues distances prédominent. Le cantilever atomique (AFM)
fléchit lorsqu’il est encore à une grande distance
de la surface. Milieu: Courbe de force idéale Une fois que l’interaction entre la
pour l’imagerie à haute résolution. Les pointe et l’échantillon est comprise, un
différentes forces s’équilibrent jusqu’au moment paramètre crucial pour l’imagerie par
où le contact physique entre la pointe et l’objet AFM est la planéité des membranes
est établi. La boucle de rétroaction va alors réagir adsorbées et la tension de ces objets
rapidement sur la déflection du cantilever en
utilisant des forces appliquées minimales. En adsorbés. Des membranes qui ne sont
bas: Très proche de la surface, les forces pas complètement planes, ou bien des
attractives sont présentes. Cette attraction va vésicules qui n’ont pas complètement
attirer constamment la pointe sur l’échantillon collapsée peuvent présenter de multiples
causant la destruction de l’objet biologique. problèmes techniques et les forces
appliquées peuvent être mal estimées.
Pour l’imagerie, cependant, les
Des structures verticales peuvent être
électrolytes doivent être choisis de façon
« comprimées » verticalement quand la
à ce que les forces électrostatiques
pointe pousse sur des régions mal
répulsives soient contrebalancées par les
13/53Habilitation à Diriger des Recherches | Simon Scheuring
attachées. Ceci peut être observées sur domaines protéiques qui dépassent peu
les courbes force-distance, qui présentent de la membrane (< 2nm) se superposent
des faibles répulsions sur des distances bien avec les représentations des
relativement larges à partir du point de surfaces calculées à partir des structures
contact initial. En conséquence, la hautement résolues (Engel and Müller,
« compression » de l’objet contribue à la 2000), il paraît déraisonnable
constante de raideur, et la boucle de d’interpréter les détails structuraux des
rétroaction qui détermine la déflection de topographies de domaines plus larges.
la pointe ne pilotera pas le piézo de Le problème devient encore plus
façon appropriée pour obtenir une image prononcé quand la géométrie de la
avec un contraste maximum. De plus, les pointe dévie de sa forme hémisphérique.
membranes qui ne sont pas bien Ce phénomène, appelé de « pointe
attachées peuvent donner des images asymétrique » ou de « double pointe »,
avec des « trainées», ce qui est peut interférer avec la fiabilité de la
probablement du au faible déplacement détection de contours qui s’étend de
de la membrane durant le balayage, au détails submoléculaires à des structures
moins en mode de contact d’imagerie. globulaires d’une taille de plusieurs
De telles « trainées » peuvent ressembler nanomètres. Cet effet rend difficile
à la dérive du piézo, mais peuvent en l’analyse de molécules non-
être distinguées car la direction des oligomériques (en particulier dans des
« trainées » suit la direction des lignes de membranes natives où ni maillage
balayage, alors que la dérive du piézo régulier ni symétrie cristallographique
entraîne une distorsion uniforme dans les peuvent être utilisés pour estimer la
images obtenues par balayage aller et les symétrie de la pointe) ou de molécules
images obtenues par balayage retour. dont aucune information structurale n’est
Ils existent d’autres paramètres disponible par ailleurs.
cruciaux pour l’imagerie par AFM et qui Un autre aspect du mécanisme
sont reliés d’une part à la physique des d’imagerie est la non-homogénéité de la
mécanismes de détection de contours résolution obtenue dans un topographe à
employés, par exemple la boucle de AFM. Le fait que la résolution n’est pas
rétroaction, et d’autre part à la mobilité égale en chaque point d’une image est
des structures de protéines qui dépassent une conséquence directe de la
de la membrane. Cette mobilité convolution de la pointe et de
augmente avec la hauteur des domaines l’utilisation d’une boucle de rétroaction.
qui dépassent de la membrane. En tenant Par exemple, on peut imaginer une
compte de ses deux effets, on s’attend situation où une particule bien résolue
généralement à voir le contraste des qui dépassent peu de la membrane est à
contours diminuer lorsque la hauteur des proximité d’une structure globulaire
structures qui dépassent augmente. sujette de façon importante à la
Lorsque des domaines globulaires convolution de la pointe et donc à la
dépassent particulièrement sont mauvaise précision de la détection. Dans
observés, la géométrie de la pointe, qui cette situation, la structure des régions
peut être assimilée en première directement à côté d’un domaine qui
approximation à une demi-sphère avec dépasse très fortement de la membrane
un rayon défini, est convoluée avec la doit être considérée comme
topographie de la surface de façon « fantaisiste », puisque le contour
significative: la mesure de la hauteur enregistré représente plus l’interaction
d’un domaine qui dépasse entre le côté de la pointe et la structure
particulièrement de la membrane peut globulaire voisine que le contact entre
être précise, mais la mesure du diamètre l’extrémité de la pointe et la surface
est élargie du fait de la convolution de la placée en dessous. Toutes ces
pointe. Alors que les topographies de considérations sont vraies lorsqu’on
14/53Habilitation à Diriger des Recherches | Simon Scheuring
considère que l’extrémité de la pointe est 2) (Gonçalves et al., 2005b; Scheuring et
en contact physique avec l’objet. La al., 2001b; Scheuring et al., 2003a),
situation devient plus compliquée si l’on représentent un système idéal pour tester
veut prendre en considération l’influence les rayons des pointes d’AFM (Fig. 6).
des propriétés physico-chimiques En première approximation, le LH2 est
particulières des surfaces à chaque point considéré comme un cylindre ouvert
de contact entre l’échantillon et la pointe parfait avec un diamètre D de 50 Å, et la
(Israelachvili, 1991; Israelachvili and pointe comme une hémisphère avec un
Wennerstrom, 1996). rayon au niveau de l’extrémité r, qui doit
Une autre limitation cruciale pour être déterminé. Quand la pointe est
l’imagerie par AFM à haute résolution a positionnée au centre du cylindre de
un rapport avec le rayon de la pointe. Il LH2, elle va pénétrer autant que possible
doit être souligné que, bien que les dans le cylindre, avec la règle évidente
fournisseurs indiquent que ce rayon est qu’une pointe très pointue entrera plus
de 10 à 50 nm, les topographes de profondément dans la molécule en forme
surfaces biologiques planes peuvent être de cylindre qu’une pointe plus arrondie
enregistrés avec une résolution (Fig. 6a). Tant que la profondeur
supérieure à 10 Å. mesurée d est inférieure à la hauteur
totale h et la moitié du diamètre D/2 de
la molécule cylindrique, elle peut être
utilisée pour déterminer le diamètre r
(Fig. 6b).
En tenant compte du fait que les
bords de la molécule sont couverts d’une
couche organisée de tampon, le cylindre
réel avec lequel la pointe interagit est
probablement plus étroit que la structure
elle-même. En conséquence, le rayon de
la pointe obtenu par le calcul précédent
est une « large estimation ». A partir des
mesures effectuées sur les complexes
LH2 de différentes bactéries
photosynthétiques (Gonçalves et al.,
2005b; Scheuring et al., 2001b;
Scheuring et al., 2003a; Scheuring et al.,
Fig. 6) a) Représentation schématique d’une 2004b), et en tenant compte des
extrémité de la pointe arrondie (‘blunt tip’) et considérations mentionnées
d’une extrémité pointue (‘sharp tip’) positionnée précédemment, nous avons calculé que
au-dessus d’une molécule cylindrique (barrel-
shaped molecule). Une extrémité pointue peut
le rayon d’une pointe d’AFM à haute
pénétrer plus profondément au centre de la résolution est de l’ordre de 2.5 nm. La
molécule cylindrique qu’une extrémité arrondie. perte de temps nécessaire pour trouver
b) Rappels sur la façon de calculer le rayon de la une pointe dont l’extrémité est aussi
pointe (r) à partir de la profondeur de pénétration pointue peut être considérée comme une
(d) et le diamètre (D) de la molécule cylindrique.
autre limitation de l’imagerie par AFM à
C’est pourquoi, les pointes haute résolution.
employées la plupart du temps doivent
avoir une aspérité de l’ordre du 4.4. Application
nanomètre. Les molécules en forme de 4.4.1. Imagerie à haute résolution –
cylindre avec de petits diamètres, détection de contours de boucles
comme les complexes collecteurs de individuelles
lumière LH2 (light-harvesting complex
Parmi les sujets de recherche
15/53Habilitation à Diriger des Recherches | Simon Scheuring
auxquels j’ai participé sur l’imagerie à 2000). Cette structure a donné des
haute résolution de protéines réponses aux questions les plus urgentes
membranaires, qui fournit des comme sur le rôle du triplet d’acides
informations structurales sur celles-ci, aminés très conservé dans le filtre du
(Scheuring et al., 1999a; Scheuring et canal ou sur la rupture du réseau de
al., 1999b; Scheuring et al., 2001a; liaisons hydrogènes par l’orientation des
Scheuring et al., 2001b; Scheuring et al., molécules d’eau dans le canal à eau. Plus
2002a), je vais discuter plus en détail ici tard, la structure d’un membre de la
le cas du canal à eau d’Escherichia coli famille des canaux de glycerol a été
l’aquaporineZ (AqpZ). J’ai pu analyser résolu : GlpF de E. coli (Fu et al., 2000).
la surface de l’AqpZ dans un Depuis, d’autres structures de AQP1
environnement natif en 1999, soit un an (Sui et al., 2001) et AQP0 (Gonen et al.,
avant que toute structure de l’aquaporine 2004; Harries et al., 2004) ont été
soit disponible. La structure de l’AqpZ obtenues par critallographie électronique
elle-même a été résolue en 2003. Une ou de rayon-X.
comparaison de la structure à haute Dans Escherichia coli, un canal à
résolution maintenant connue avec la eau a été identifié par clonage
topographie obtenue par AFM, et son d’homologie, nommé AqpZ. Le AqpZ
interprétation, permet d’évaluer de façon recombinant, qui porte une étiquette
non biaisée la fiabilité de l’imagerie histidine, a été produit et il a été montré
ainsi que les limites de l’interprétation qu’il était actif (Calamita et al., 1998).
des topographies. Des cristaux à 2D avec des tailles allant
Les aquaporines sont des canaux jusque 5 µm ont été assemblés à partir de
omniprésentes dans les membres des cet AqpZ recombinant (Ringler et al.,
bactéries, des plantes, des champignons 1999). L’AFM a été utilisé pour mesurer
et des animaux. Elles sont hautement la topographie de surface des cristaux de
spécifiques à l’eau ou aux petits solutés AqpZ dans un environnement natif. La
hydrophiles non chargés et sont définition des côtés de AqpZ a ainsi été
impliquées dans la régulation de déterminée par imagerie de cristaux à 2D
l’osmose. L’analyse d’hydropathies des reconstitués à partir de AqpZs portant
premiers membres indiquait six une étiquette poly-histidine N-terminale,
domaines membranaires et deux boucles et par imagerie des cristaux-2D
inhabituelles (Agre et al., 1993; Gorin et identiques obtenus après protéolyse des
al., 1984; Jung et al., 1994). Depuis ce extrémités N-terminales (Scheuring et
temps, près de deux cents gènes al., 1999b). Les résolutions latérale et
d’aquaporines ont été séquencées, et verticale des images sont 8 Å et 1 Å
parmi celles-ci 10 chez les humains. respectivement (Fig. 7a). A cette
Presque toutes ont les motifs hautement résolution et avec un haut rapport signal
conservés NPA dans leurs boucles. sur bruit, des boucles individuelles
(Heymann and Engel, 2000). surplombant la surface ont été observées
Approximativement la moitié de ces pour chaque monomère des tétramères
canaux protéiques sont exclusivement de AqpZ. La topographie moyenne (Fig.
sélectif pour l’eau et ne permettent pas le 7b) a été comparée avec la prédiction de
passage de petits solutés hydrophiles. la structure basée sur la séquence
D’autres canaux facilitent pourtant le d’AqpZ indiquant que cette protéine
passage de ces petites molécules comme contient six hélices transmembranaires,
le glycérol ou l’urée. connectées avec trois boucles du côté
La première structure d’aquaporine extracellulaire et deux boucles du côté
à avoir été résolue est celle de AQP1 ; cytoplasmique, ainsi que deux extrémités
elle a été réalisée par crystallographie terminales cytoplasmiques. En accord
électronique à partir de cristaux à 2D avec ceci, trois protrusions ont été
hautement ordonnés (Murata et al., trouvées sur la surface extracellulaire
16/53Habilitation à Diriger des Recherches | Simon Scheuring
(Fig. 7b, cercle bleu) du monomère de d’ordre 4 a un volume d’environ 1300
AqpZ, une proche du centre de symétrie Å3, la petite protrusion à la périphérie en
d’ordre 4, ainsi qu’une petite et une plus a un d’environ 1000 Å3, tandis que la
allongée à la périphérie. protrusion allongée a un volume
d’environ 3200 Å3. La protrusion unique
observée par monomère sur la surface du
côté cytoplasmique (Fig. 7b, cercle
jaune) a un volume d’environ 1200 Å3.
En comparant la topographie
obtenue par AFM avec la structure
récente de AqpZ obtenue avec une
résolution de 2.5 Å par cristallographie à
rayons-X (Savage et al., 2003), il est
maintenant possible d’évaluer,
rétrospectivement, la précision de la
topographie et de l’interprétation des
données. Pour ce faire, la surface
moléculaire de la structure d’AqpZ a été
calculée (Fig. 7c). La précision de la
surface obtenue par AFM est surprenante
comparée à celle de la structure par
rayons-X avec une résolution de 2.5 Å,
sur les deux surfaces périplasmique (Fig.
Fig. 7) a) Topographe AFM à haute résolution 7c, gauche) et cytoplasmique avec les
d’un cristal à 2D d’AqpZ. Les canaux à eau extrémimités terminales retirées (Fig. 7c,
forment des tétramères insérés vers le haut et
vers le bas par rapport à la surface de la
droite). Sur la surface périplasmique, la
membrane. Les boucles individuelles des canaux boucle a culminant avec Pro30 (et
à eau tétramériques sont résolues dans l’image s'étendant de Gly28 à Leu32), la fin de
(donnée brute). b) Topographie moyenne d’un l'helice 3 et le début de la boucle c
cristal à 2D d’AqpZ. Les surfaces sont culminant avec Thr107 (et s'étendant de
encerclées, la forme de couronne extracellulaire
en bleu et la forme de croix cytoplasmique en
Gly105 à Phe109), et la longue boucle c
jaune. c) Comparaison de la topographie obtenue à la périphérie du tetramère sont
par AFM et de la structure récente de AqpZ délimitées de manière crédible. Du fait
obtenue par rayons-X. La surface moléculaire de sa longue séquence en acides aminés
calculée à partir de la structure obtenue par et de son volume important dans la
rayons-X est représentée. La surface
extracellulaire est montrée sur la gauche (en
topographie, la boucle c périphérique a
bleu), et la surface cytoplasmique sur la droite été correctement assignée. La petite
(en jaune). La surface moléculaire est colorée protrusion centrale, logeant la boucle a, a
avec des intensités croissantes suivant la hauteur été mise en évidence comme telle
des domaines protéiques qui dépassent de la (Scheuring et al., 1999b). Sur la surface
membrane. Les boucles qui connectent des
hélices, qui ont une taille d’environ 5 acides
cytoplasmique, la structure X révèle une
aminés (voir la boucle centrale a) sont petite hélice de 3 acides aminés dans la
déterminées de façon fiable. boucle b (His56, Ile57, Ser58) comme
étant la structure protudant le plus – elle
Comme, à cette date, aucune est située en amont de la boucle b qui
structure à haute résolution de descend dans le canal à eau pour former
l’aquaporine n’est disponible, des calculs le filtre. La topographie AFM de la
de volume sur les protrusions ont été surface cytoplasmique, étant donné que
effectués pour associer la topographie les termini ont été enlevés par des
aux boucles reliant les hélices. La protéolyses, contient les boucles b et d
protrusion proche du centre de symétrie sur cette surface (Scheuring et al.,
17/53Vous pouvez aussi lire