Afférences cholinergiques au noyau sous-thalamique et leur altération dans la maladie de Parkinson - Mémoire Maya Chebl Maîtrise en neurobiologie ...
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Afférences cholinergiques au noyau sous-thalamique et
leur altération dans la maladie de Parkinson
Mémoire
Maya Chebl
Maîtrise en neurobiologie - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© Maya Chebl, 2019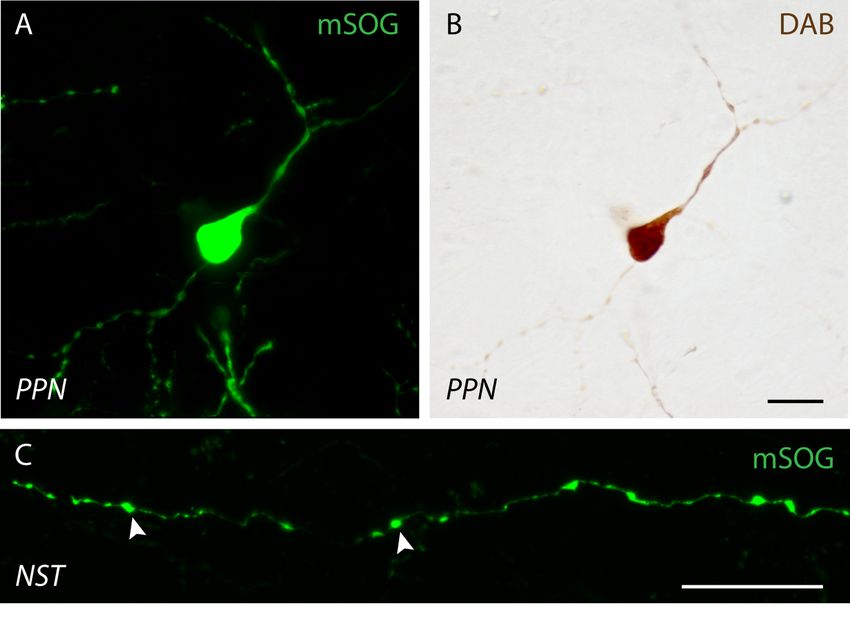
Résumé
Il existe dans le cerveau un regroupement de structures sous-corticales que l’on nomme
ganglions de la base (GB) et qui contrôle de nombreuses fonctionnalités psychomotrices.
L’apparition de débalancements neurochimiques dans l’une de ces structures est suffisante
pour induire des phénotypes pathologiques extrêmement déstabilisants. L’un des syndromes
les plus fréquemment rencontrés est la maladie de Parkinson (MP), une affection des GB
dont la principale déterminante neuropathophysiologique implique principalement les
neurones dopaminergiques (DA) de la substance noire pars compacta (SNc). La
dégénérescence marquée de ces neurones ne peut pourtant pas, à elle seule, expliquer
l’ensemble des symptômes dont font montre les patients. L’usage thérapeutique du
précurseur de la DA, la L-dopa, ne se montre pas infaillible face aux troubles posturaux
caractéristiques de la MP. Récemment, des études anatomiques et électrophysiologiques
suggèrent que l’acétylcholine (ACh) pourrait jouer un rôle déterminant dans ce désordre
neurologique. Ces travaux mettent l’emphase sur la dégénérescence des neurones ACh du
noyau pédonculopontin, une structure du tronc cérébral dont les axones ascendants
s’arborisent massivement dans le thalamus, mais aussi dans le noyau sous-thalamique (NST),
une composante-clé des GB fortement dérégulée dans la MP. Ces données nous ont orientés
vers l’étude de l’innervation ACh du NST en conditions normale et pathologique dans le tissu
humain post-mortem. Nous avons eu recours à l’immunohistochimie et à la stéréologie pour
obtenir des estimations non-biaisées du nombre de varicosités axonales ACh dans la région
d’intérêt. Les résultats obtenus mettent en évidence, et ce pour la première fois, une baisse
significative de la neurotransmission ACh dans le NST des sujets MP, une diminution
important affectant tous les secteurs du NST. Ces résultats soulignent l’importance de tenir
compte de l’impact fonctionnel de l’innervation ACh dans le NST et considérer son
implication possible dans les dérèglements de l’activité neuronale du NST qui survient dans
la MP.
ii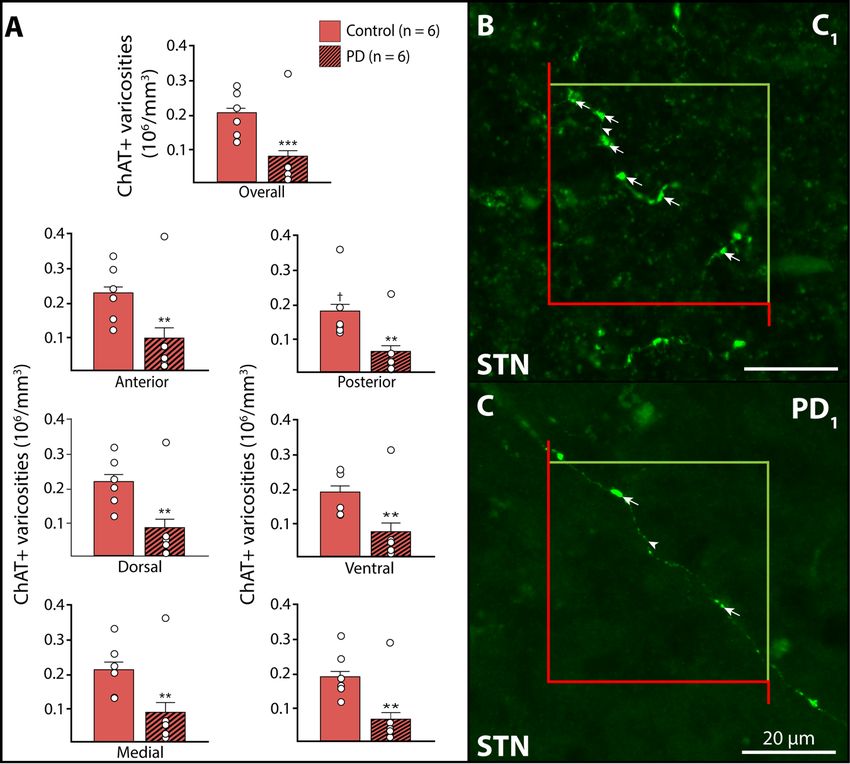
Abstract
The basal ganglia (BG) – a set of subcortical structures lying at the basis of the cerebral
hemispheres – are known to play a crucial role in the control of psychomotor functions. A
single neurochemical imbalance in one of the components of the BG can induce severe motor
disabilities that range from hyperkinesia to hypokinesia. One of the most frequently
encountered syndromes associated with BG malfunction is Parkinson’s disease (PD). The
pathological hallmark of PD, clinically characterized by tremor, bradykinesia and rigidity, is
the death of the dopaminergic (DA) neurons of the substantia nigra pars compacta (SNc).
However, the marked reduction of the DA neurons does not, by itself, explain the entire
symptomatic spectrum experienced by the patients. In fact, the main pharmacological
treatment of PD, which is based on the administration of L-dopa, the metabolic precursor of
DA, to restore the endogenous DA store, does not alleviate all symptoms of this
neurodegenerative disease, postural instabilities being particularly resistant to L-dopa
therapy. Recently, numerous anatomical and electrophysiological studies have revealed that
acetylcholine (ACh) might also be involved in the pathogenesis of this neurodegenerative
disorder. Of particular significance was the report of a loss of the ACh neurons contained in
the peduculopontine nucleus, an upper brainstem nucleus that projects massively to the
thalamus, but also to the subthalamic nucleus (STN), a key component of the BG that is
markedly dysregulated in PD. These findings led us to analyze post-mortem brain tissue
collected from normal individuals and PD patients in order to determine quantitatively the
strength of the ACh innervation in the STN in normal and pathological conditions. Unbiased
estimations of the number of ACh axonal varicosities in the different sectors of the STN were
gathered by applying stereological methods to material immunostained for an ACh marker.
Our results have provided a first direct evidence for a significant reduction of the ACh
neurotransmission in the STN of parkinsonian patients, a decrease that is evident in all
regions of the STN. These findings suggest that the ACh input plays an important role in the
normal functioning of the STN and that it might even be involved in the induction of the
abnormal neuronal activity that characterizes this nucleus in PD.
iii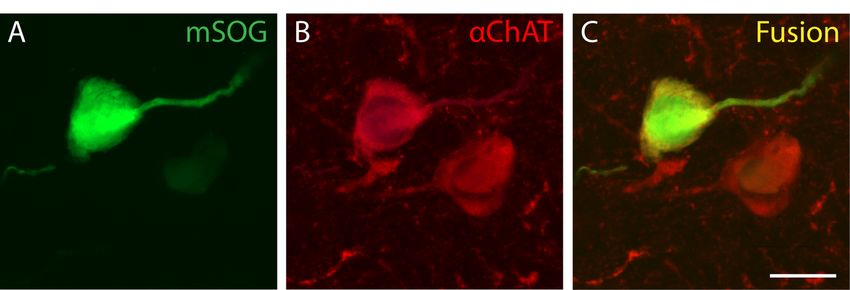
Table des matières
Résumé ..................................................................................................................................................... ii
Abstract ................................................................................................................................................... iii
Table des matières ................................................................................................................................... iv
Liste des figures ....................................................................................................................................... vi
Liste des abréviations ..............................................................................................................................vii
Remerciements ....................................................................................................................................... viii
Avant-propos ............................................................................................................................................ix
INTRODUCTION GÉNÉRALE .............................................................................................................. 1
1.1. Préambule ......................................................................................................................................... 1
1.2. Les ganglions de la base ................................................................................................................... 2
1.2.1. Le striatum ............................................................................................................................... 2
1.2.2. La substance noire ................................................................................................................... 4
1.2.3. Le globus pallidus ..................................................................................................................... 5
1.2.4. Le noyau sous-thalamique ....................................................................................................... 6
1.3. L’acétylcholine dans le système nerveux central ............................................................................ 11
1.3.1. Origine des afférences cholinergiques ................................................................................... 11
1.3.2. Caractéristiques de l’innervation cholinergique du noyau sous-thalamique......................... 13
1.3.3. Fonctions de l’innervation cholinergique............................................................................... 16
1.3.4. Types de récepteurs ............................................................................................................... 19
1.4. La maladie de Parkinson ................................................................................................................. 21
1.4.1. Historique............................................................................................................................... 21
1.4.2. Épidémiologie ........................................................................................................................ 22
1.4.3. Stigmates neuropathologiques traditionnels ......................................................................... 23
1.4.4. Dégénérescence cholinergique .............................................................................................. 25
1.4.5. Traitements ............................................................................................................................ 26
1.5. Objectif ........................................................................................................................................... 30
1.6. Hypothèse ....................................................................................................................................... 30
1.7. Approches méthodologiques .......................................................................................................... 30
iv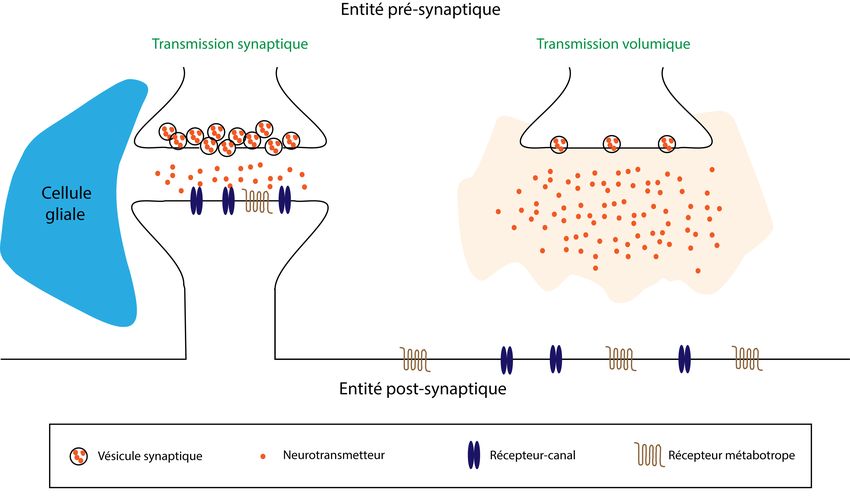
CHAPITRE 1 – THE CHOLINERGIC AFFERENT PROJECTIONS TO THE SUBTHALAMIC
NUCLEUS IN PARKINSON’S DISEASE ............................................................................................. 33
2.1. Résumé ........................................................................................................................................... 34
2.2. Abstract .......................................................................................................................................... 35
2.3. List of abbreviations........................................................................................................................ 36
2.4. Introduction .................................................................................................................................... 37
2.5. Material and methods .................................................................................................................... 39
2.5.1. Human brain tissue ................................................................................................................ 39
2.5.2. Immunohistochemistry .......................................................................................................... 39
2.5.3. Quantitative assessment of ChAT-immunoreactive axon varicosities in the STN.................. 40
2.5.4. Statistical analysis .................................................................................................................. 42
2.6. Results............................................................................................................................................. 42
2.6.1. ChAT immunoreactivity of the STN and morphological aspects of ChAT+ STN axons ........... 42
2.6.2. STN regional distribution of ChAT+ axon varicosities in normal condition ............................ 43
2.6.3. Density of ChAT+ axon varicosities in Parkinson’s disease .................................................... 43
2.7. Discussion ....................................................................................................................................... 44
2.7.1. ACh innervation of the normal brain ..................................................................................... 44
2.7.2. ACh innervation of the STN in PD .......................................................................................... 45
2.7.3. Neuropathological correlates ................................................................................................ 47
2.8. Acknowledgements ......................................................................................................................... 48
2.9. Table and figures ............................................................................................................................ 49
CONCLUSIONS ..................................................................................................................................... 54
ANNEXE - ANALYSES ULTRASTRUCTURALE ET TRIDIMENSIONNELLE DE SEGMENTS
AXONAUX CHOLINERGIQUES GRÂCE À LA PHOTOCONVERSION DE LA PROTÉINE
MINISOG CHEZ LA SOURIS CHAT-CRE.......................................................................................... 58
A.1. Introduction .................................................................................................................................... 59
A.2. Protocole expérimental ................................................................................................................... 60
A.2.1. Construction du vecteur viral exprimant miniSOG ................................................................ 60
A.2.2. Animaux et chirurgies stéréotaxiques.................................................................................... 61
A.2.3. Préparation des solutions pour la perfusion transcardiaque ................................................. 61
A.2.4. Perfusion transcardiaque et dissection des cerveaux ............................................................ 62
A.2.5. Préparation des solutions pour la photo-oxidation de miniSOG ........................................... 63
A.2.6. Protocole de photo-oxidation de miniSOG ............................................................................ 64
A.2.7. Protocole pour la microscopie électronique à transmission (TEM) ....................................... 64
A.2.8. Protocole pour la microscopie électronique à balayage (FIB-SEM) ....................................... 65
A.3. Résultats représentatifs .................................................................................................................. 67
A.4. Discussion ....................................................................................................................................... 67
Références ............................................................................................................................................... 75
v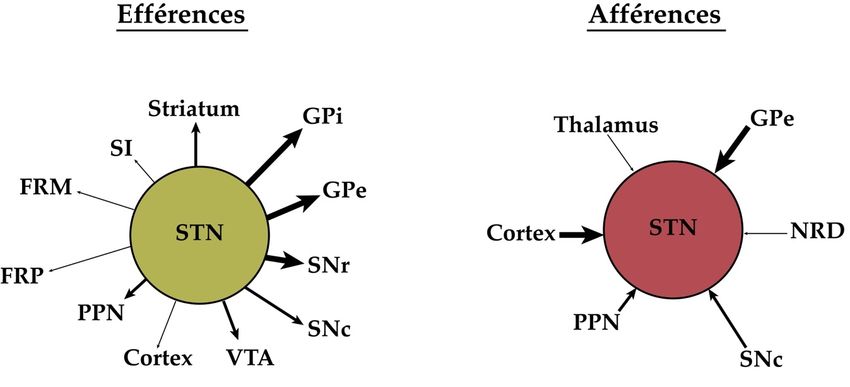
Liste des figures
Figure 1.1. Organisation classique des circuits des ganglions de la base..................................3
Figure 1.2. Anatomie et organisation topographique du noyau sous-thalamique.....................8
Figure 1.3. Vue d’ensemble des principales connexions anatomiques existant au sein du
noyau sous-thalamique..........................................................................................................10
Figure 1.4. Un neurotransmetteur relâché au niveau d’une synapse tripartite ou dans le milieu
extracellulaire loin de sa cible................................................................................................15
Table 2.1. Clinical information of the human post-mortem brains used in the study..............49
Figure 2.1. Experimental approach used for the unbiased stereological count of ChAT+ axon
varicosities in the STN...........................................................................................................50
Figure 2.2. Differences in the density of ChAT+ immunostained axon varicosities between
PD and age-matched controls................................................................................................52
Figure A.1. Construction du virus exprimant miniSOG selon le système cre-lox dans les
neurones cholinergiques du PPN de la souris........................................................................60
Figure A.2. Représentation schématique de la procédure expérimentale menant à la photo-
oxidation de miniSOG...........................................................................................................70
Figure A.3. Dans les souris injectées, l’immunomarquage avec l’anticorps contre la choline
acétyltransférase (ChAT) indique que miniSOG est exclusivement exprimé dans les neurones
cholinergiques du PPN..........................................................................................................72
Figure A.4. Exemple d’une photo-conversion de miniSOG en un précipité de chromogène
DAB......................................................................................................................................73
Figure A.5. Micrographie électronique mettant en évidence des éléments neuronaux marqués
à la DAB................................................................................................................................74
vi
Liste des abréviations
5-HT Sérotonine
6-Hydroxydopamine 6-OHDA
AADC Acide L-aminé décarboxylase
ACh Acétylcholine
AChE Acétylcholinestérase
ChAT Choline acétyltransférase
CL Corps de Lewy
D1 Récepteur dopaminergique 1
D2 Récepteur dopaminergique 2
D5 Récepteur dopaminergique 5
DA Dopamine
DIL Dyskinésies induites par la L-dopa
GB Ganglions de la base
GPe Segment externe du globus pallidus
GPi Segment interne du globus pallidus
L-dopa L-3,4-hydroxyphénylalanine
LDT Noyau tegmentaire latérodorsal
MA Maladie d’Alzheimer
mAChRs Récepteurs muscariniques
MP Maladie de Parkinson
MPTP 1-méthyl-4-phényl-1,2,3,6-tétrahydropyridine
nAChRs Récepteurs nicotiniques
nbM Noyau basalis de Meynert
NPS Neurones de projection du striatum
NST Noyau subthalamique
PPN Noyau tegmentaire pédonculopontin
SNc Substance noire pars compacta
SNr Substance noire pars reticulata
SRAA Système réticulaire activateur ascendant
TCSP Trouble comportemental du sommeil paradoxal
TEP Tomographie par émission de positrons
TH Tyrosine hydroxylase
VTA Aire tegmentaire ventrale
ZI Zona incerta
vii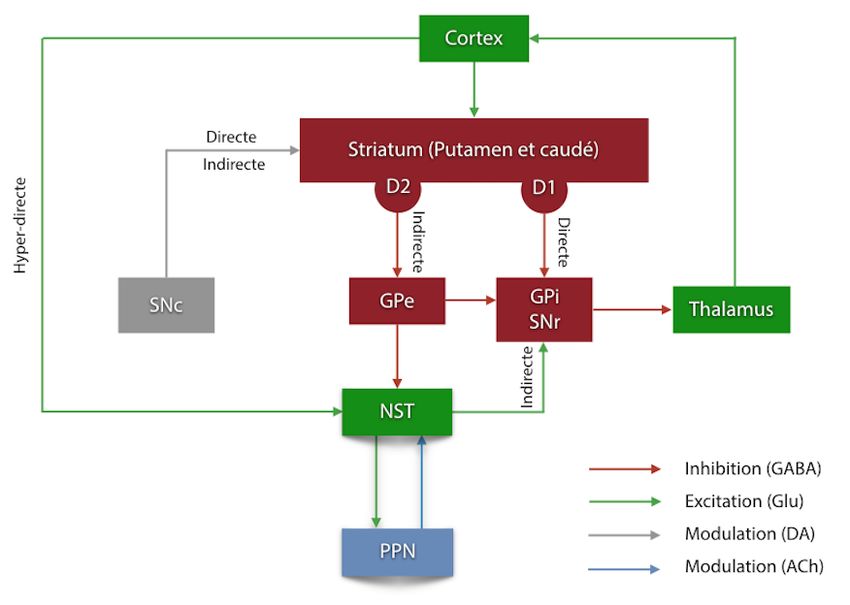
Remerciements
Je voudrais tout d’abord adresser un lot immense de gratitude à mon directeur de recherche,
Dr. Martin Parent, qui s’est toujours montré accessible et disponible pour me recevoir dans
son bureau et répondre à mes interrogations et mes doutes. Ses mots d’encouragement et sa
patience ont été des plus appréciés. J’aimerai également remercier mon co-directeur de
recherche, Dr. Christophe Proulx, pour tous les judicieux conseils qu’il m’a fournis.
Je voudrais en outre démontrer ma reconnaissance envers Dr. André Parent. Merci de m’avoir
inculqué une part de votre sagesse scientifique et humaine. Sachez que votre passion pour les
neurosciences a inspiré tou(te)s les étudiant(e)s qui ont croisé votre chemin.
Merci à tous les membres du laboratoire, présents et passés. Merci à Marie-Josée Wallman,
ton support technique mais particulièrement, ton support moral avec ta bonne humeur, m’ont
été indispensables pour traverser toutes les étapes de cette maîtrise. Merci à Dr. Dave
Gagnon, sur qui je pouvais toujours compter. Merci à Dr. Laurent Goetz, Dymka Coudé,
Francis Desmeules, Damon DePaoli, Vincent Coulombe, Lydia Saidi et Tiago David. Vous
avez su égayer mes journées au laboratoire.
Finalement, je voudrai remercier mes parents, Anna et Zouheir, et mon frère, Nicolas. Votre
amour inconditionnel et votre écoute, même à des centaines de kilomètres de distance, ont
été le fruit de cet accomplissement. Merci de ne jamais avoir baissé les bras devant moi.
viiiAvant-propos
Les travaux présentés dans ce mémoire visent à ajouter aux connaissances que nous
possédons sur le système cholinergique dans les ganglions de la base et à apporter une
nouvelle perspective concernant les changements qui lui sont propres dans la maladie de
Parkinson. Mes travaux auront permis d’élaborer sur la complexité déjà fort bien établie
entourant ce désordre neurologique du mouvement.
Ce mémoire s’articule en premier lieu autour d’une introduction générale couvrant
l’organisation des composantes des ganglions de la base, le système cholinergique dans le
système nerveux central et une description exhaustive de la pathophysiologie de la maladie
de Parkinson.
Le deuxième chapitre fait l’objet d’un article qui sera soumis au Journal of Chemical
Neuroanatomy. Je serai la première auteure de cet article, ayant réalisé toutes les
expérimentations, l’analyse des résultats et la rédaction du manuscrit. J’ai toutefois reçu la
précieuse aide technique de Marie-Josée Wallman et Dr. Dave Gagnon tout au long de la
réalisation de cette étude. J’aimerai également mentionner que ce projet n’aurait pu être
effectué sans l’implication de mon directeur de recherche dans son élaboration, la correction
du manuscrit et ses conseils dont je n’aurais pu me passer.
L’annexe jointe à ce mémoire correspond à l’ébauche d’une approche méthodologique
servant à la reconstruction de segments axonaux en microscopie électronique à balayage. J’y
élabore le protocole qui a été suivi ainsi que l’ensemble des résultats et les limitations
techniques qui doivent encore être revisitées.
ixINTRODUCTION GÉNÉRALE
1.1. Préambule
Les êtres humains sont constamment à la poursuite du changement. Ce désir, ou besoin, est
grandement propulsé par l’envie de surmonter de nouveaux défis, de dévier des activités
routinières qui font souvent partie intégrante de nos vies ou encore de satisfaire à notre
curiosité, pour n’en citer que quelques-uns. Dans ce cas, le changement provient d’une
motivation intrinsèque et les conséquences qui en découlent sont adoptées avec un plaisir
inhérent à la personne. Toutefois, il arrive que le changement s’impose à nous et oblige ainsi
notre comportement à être modulé de telle sorte qu’il réponde aux nouvelles exigences. Au
cours de cette instance, nous nous retrouvons dépourvus de nos moyens et cela peut mener
ultimement à une situation de crise. Le fonctionnement du cerveau peut être perçu de manière
similaire. L’acquisition de nouvelles connaissances, cet apprentissage qu’une personne
décide par elle-même de poursuivre, engendre un changement dans la force de la
communication neuronale au sein de certaines structures cérébrales (on parle souvent de
plasticité neuronale). L’adaptation cellulaire qui s’ensuit est fort bien reçue, car la finalité qui
en résulte est considérée comme étant favorable. En revanche, des changements non désirés
se font aussi prévaloir et perturbent l’homéostasie cellulaire, cet équilibre du milieu que le
cerveau cherche à maintenir au meilleur de ses capacités. Les maladies neurodégénératives,
comme la maladie d’Alzheimer (MA) ou de Parkinson (MP), sont des exemples concrets de
cette situation. La perte de populations neuronales ciblées se traduit par un changement que
le cerveau peine à compenser correctement. Un déséquilibre à petite et grande échelles se fait
ressentir et les conséquences sont fâcheuses. Naturellement, nous envisageons la possibilité
de les minimiser. Pour ce faire, il est primordial de comprendre les systèmes impliqués qui
produisent les symptômes apparents. C’est dans cette optique que j’en viens à présenter le
premier volet de ce mémoire. Il comprendra un exposé qui vise à expliquer la transmission
neuronale de l’acétylcholine (ACh) en lien étroit avec les ganglions de la base (GB), cet
ensemble de noyaux sous-corticaux dont les fonctions sont d’orchestrer et exécuter des tâches
motrices, limbiques et cognitives (Kim et Hikosaka, 2015). Le tout sera intégré à un contexte
pathologique, soit le cas de la MP, où les dérèglements du système ACh seront évalués et la
pertinence de son implication dans l’apparition des symptômes tentera d’être élucidée.
11.2. Les ganglions de la base
Comme il a été mentionné dans le court préambule ci-haut, les GB interviennent à des étapes
précises des systèmes moteur, exécutif, cognitif et limbique. Cela se fait au sein de circuits
intrinsèques qui se chevauchent considérablement et de facto rendent son organisation
générale beaucoup plus complexe, contrairement à ce qui avait été préalablement anticipé
avec les voies directe et indirecte seulement. Ces voies peignaient alors une organisation plus
simpliste, avec le striatum comme principale porte d’entrée. Depuis quelques décennies, les
préceptes entourant les GB ont été revistés et l’emphase sur de nouvelles connexions a vu le
jour (Afsharpour, 1985; Nambu, Tokuno et Takada, 2002) Il n’en demeure pas moins que les
éléments qui sont sous-jacents à cette organisation sont inchangés et ils feront ainsi l’objet
de cette première partie de l’introduction, dans laquelle une description détaillée sera
proposée pour chacun d’entre eux. Une emphase sera placée sur le noyau sous-thalamique,
puisque cette structure-clé des GB constitue le sujet principal de mon étude.
1.2.1. Le striatum
Le striatum constitue la principale porte d’entrée des GB puisque cette structure reçoit des
projections massives de l’ensemble du cortex cérébral. Il comprend une partie dorsale, elle-
même divisée par la capsule interne en noyau caudé et putamen chez les primates, qui se
charge du relais des informations motrices et sensorielles ainsi qu’une partie ventrale (noyau
accumbens) qui représente le territoire limbique (cette partie ne sera pas élaborée dans le
cadre de ce projet). Aux fins de ce mémoire, l’usage du terme striatum réfère essentiellement
au striatum dorsal, soit la partie correspondant au territoire sensorimoteur (Parent et Hazrati,
1995a).
Le striatum renferme une diversité importante de neurones, tant en termes d’organisation
morphologique que de contenu neurochimique. La population neuronale la plus abondante
est constituée de neurones épineux de taille moyenne, qui représentent les neurones de
projection du striatum (NPS) (Difiglia, Pasik et Pasik, 1980; Smith et Bolam, 1990). Ce sont
des neurones GABAergiques qui reçoivent sur leurs épines, concentrées au niveau de la
partie distale des dendrites, la plus grande partie des afférences extrinsèques du striatum,
2incluant celles provenant du cortex cérébral, du thalamus et de la substance noire pars
compacta (SNc) (Parent et Hazrati, 1995a). Deux sous-populations principales de NPS ont
été caractérisées en fonction de l’expression complémentaire de certains neuropeptides et
récepteurs dopaminergiques. La première arbore une double immunoréactivité pour la
substance P et la dynorphine, et exprime à sa surface le récepteur dopaminergique de type 1
(D1). Ces neurones projettent au segment interne du globus pallidus (GPi) et à la substance
noire pars reticula (SNr). Ils constituent, de ce fait, le substrat morphologique de la voie
striatofuge directe des GB. En contrepartie, l’autre sous-population a été identifiée par le
biais de son immunoréactivité à l’enképhaline et l’expression membranaire du récepteur
D2. Les axones de ces neurones se projettent sur le segment externe du globus pallidus (GPe).
Ces neurones constituent par conséquent le premier relais de la voie indirecte des GB. Plus
récemment, une troisième sous-population de NPS a été identifiée, ces neurones étant dotés
de récepteurs D1 et D2, et exprimant uniquement la dynorphine (Gagnon et al., 2017; Lee et
al., 2004). En plus des récepteurs dopaminergiques, les neurones striataux expriment à leur
surface des récepteurs ACh et glutamatergiques.
3Figure 1.1. Modèle classique de l’organisation anatomique et fonctionnelle des ganglions de
la base.
Le striatum renferme également une deuxième population de neurones qui diffèrent de la
première par le fait que leurs axones demeurent à l’intérieur de la structure ; il s’agit de la
famille des interneurones striataux. Bien que ces derniers soient présents en plus faible
proportion (1% chez les humains, 3 à 5% chez les rongeurs, et 20 % chez les primates)
(Graveland et DiFiglia, 1985; Kawaguchi, 1993; Mesulam, Mash, Hersh, Bothwell et Geula,
1992), ils interviennent à plusieurs niveaux dans le traitement de l’information qui est
acheminée au striatum (Gittis et Kreitzer, 2012). Deux populations d’interneurones sont
rencontrées dans cette structure : les interneurones GABAergiques de tailles moyennes et les
interneurones ACh à large diamètre, qui peuvent également se différencier par les
neuropeptides ou protéines qu’ils renferment.
La place du striatum dans l’organisation fonctionnelle des GB a reçu une attention croissante
en tant que centre intégrateur des afférences provenant en grande partie du cortex cérébral.
Ces informations se transmettent ensuite au sein d’une boucle fermée correspondant à la
boucle cortico-GB-thalamo-corticale (Parent et Hazrati, 1995a).
1.2.2. La substance noire
La substance noire est divisée en deux compartiments : la substance noire réticulée (SNr) et
la substance noire compacte (SNc). Leurs délimitations anatomiques (quoiqu’elles ne soient
pas strictes) sont surtout déterminées par le bagage neurochimique des neurones qui
composent ces deux structures. En effet, la SNr est habitée par de larges neurones
GABAergiques ; en contrepartie, une grande proportion du système dopaminergique du
mésencéphale est contenue dans la SNc. Ces neurones immunopositifs pour la tyrosine
hydroxylase (TH) sont aisément reconnus par la présence marquée de neuromélanine et par
une arborisation axonale dense et vaste.
Mis à part la divergence dans la nature de leur neurotransmetteur principal, ces deux
composantes sont aussi uniques du point de vue de leurs afférences. La SNr est fortement
4innervée par les neurones D1 du striatum (voie directe), comme il a été mentionné
préalablement, mais reçoit également des projections du GPe et du noyau sous-thalamique
(NST). Puisque les neurones de la SNr projettent au thalamus, cette structure est considérée
comme étant une porte de sortie des GB, l’autre étant le GPi avec qui elle partage des
similarités. À cette projection vers les noyaux moteurs du thalamus s’ajoute une projection
vers le collicule supérieur et le tegmentum mésencéphalique (Parent et Hazrati, 1995a). Les
propriétés électrophysiologiques des neurones de la SNr lui permettent d’entretenir une
inhibition tonique sur ses cibles et jouent ainsi un rôle important sur le contrôle fin du
comportement moteur (Deniau, Mailly, Maurice et Charpier, 2007). La SNc conserve des
connexions réciproques avec plusieurs structures. Nous pouvons faire mention ici du
striatum, du GPe, du NST et du noyau pédonculopontin (PPN). L’influence dopaminergique
provenant de la SNc a, quant à elle, un rôle substantiel à jouer dans l’apprentissage moteur
et la régulation des circuits de la récompense.
1.2.3. Le globus pallidus
Comme il a été exposé antérieurement, le globus pallidus comporte deux segments chez les
primates, qui se dissocient anatomiquement par la lame médullaire interne (étroit faisceau de
fibres myélinisées) ainsi que par la nature des afférences striatales qu’ils reçoivent.
Cependant, ils sont morphologiquement et phénotypiquement similaires, contenant tous deux
une population neuronale GABAergique.
Le GPi occupe une place équivalente à la SNr, ces deux structures agissant en tant que portes
de sortie de par leurs projections au tiers ventral du thalamus. D’ailleurs, la SNr est parfois
considérée comme une extension caudo-médiale du GPi. Toutefois, ces deux structures
acheminent des informations en parallèle et s’intègrent à des voies fonctionnellement
distinctes (Nambu, 2007; Parent et Hazrati, 1995a). En effet, le GPi contrôle le comportement
somato-moteur via une influence indirecte au cortex moteur alors que la SNr entretient des
connexions indirectes avec le cortex préfrontal et contrôle davantage les fonctions
associatives (Nambu, 2007). En outre, le GPi démontre une somatotopie interne où les
neurones confinés dans sa région déchargent en réponse au mouvement du membre qui lui
5est associé. Ceci serait le résultat d’une convergence des projections venant du GPe, du NST
et du striatum.
Bien que le GPe soit pourvu de neurones ayant une morphologie somatodendritique
semblable à ceux du GPi (Parent et Hazrati, 1995b), il lui est fonctionnellement distinct. Le
GPe occupe une position intermédiaire entre le striatum et le NST au sein de la voie indirecte
et l’action inhibitrice qu’il exerce sur le NST est par conséquent cruciale dans la régulation
de son activité. Cependant, l’action du GPe ne se limite pas à la voie indirecte puisque les
neurones qui le composent projettent également au GPi.
1.2.4. Le noyau sous-thalamique
Le sujet de recherche du présent mémoire s’articule autour de l’étude du NST. Il s’agira donc
ici de détailler les connaissances que nous possédons à son égard et de tenter, dans une
prochaine section, d’élucider notre compréhension de l’impact qu’ont les changements
neuropathologiques sur son fonctionnement.
1.2.4.1. Historique
Le NST humain a été décrit anatomiquement pour la première fois par Jules Bernard Luys
(1828-1897) dans son traité de 1865 intitulé Recherches sur le système cérébro-spinal, sa
structure, ses fonctions et ses maladies. Jules Bernard Luys était un anatomopathologiste,
aliéniste (ancien titre que l’on attribuait aux personnes traitant les patients psychiatriques) et
neurobiologiste (Parent, 2017). Il obtint le titre de médecin en 1857 après avoir défendu une
thèse portant sur la tuberculose et décroche le poste de médecin des hôpitaux de Paris en
1862 (Parent, 2002). Son parcours professionnel s’édifie autour de la caractérisation
anatomique, pathologique et fonctionnelle du système nerveux central (SNC) par l’usage de
la microscopie et de l’iconographie (qui a fait l’objet du premier atlas sur le cerveau humain,
publié en 1873).
La dénomination que Luys a choisi pour désigner le NST était la bandelette accessoire des
olives supérieures ou bandelette accessoire du noyau rouge de Stilling. Il la décrit comme
6« un amas de substance grisâtre, disposé sous forme linéaire, renflé dans sa portion médiane
et atténué à chacune de ses extrémités ». Auguste Henri Forel (1848-1931), dans la
description qu’il en fera en 1877, considère que le terme de bandelette utilisé par Luys est
inapproprié, puisqu’elle fait référence à la matière blanche, contraire à la nature réelle du
NST. Il décide de le renommer « corps de Luys » (corpus Luysii), rendant ainsi hommage à
son collègue français. Plus tard, dans un article publié en 1886 dans le journal L’Encéphale,
Luys fait l’éloge du NST en tant que « nouvelle région de substance grise située à la base de
l’encéphale » ayant « échappé à l’attention des anatomistes [français] » (Luys, 1886). Il
stipule que le NST est un centre de dispersion des afférences cérébelleuses et attribue d’ores
et déjà à cette structure une place pivot dans l’intégration des commandes motrices. Ces
spéculations seront plus tard confirmées par des expérimentations sur le NST qui pouvaient
générer une réponse motrice variable et de forte intensité suite à une destruction ou une
stimulation de cette structure nerveuse.
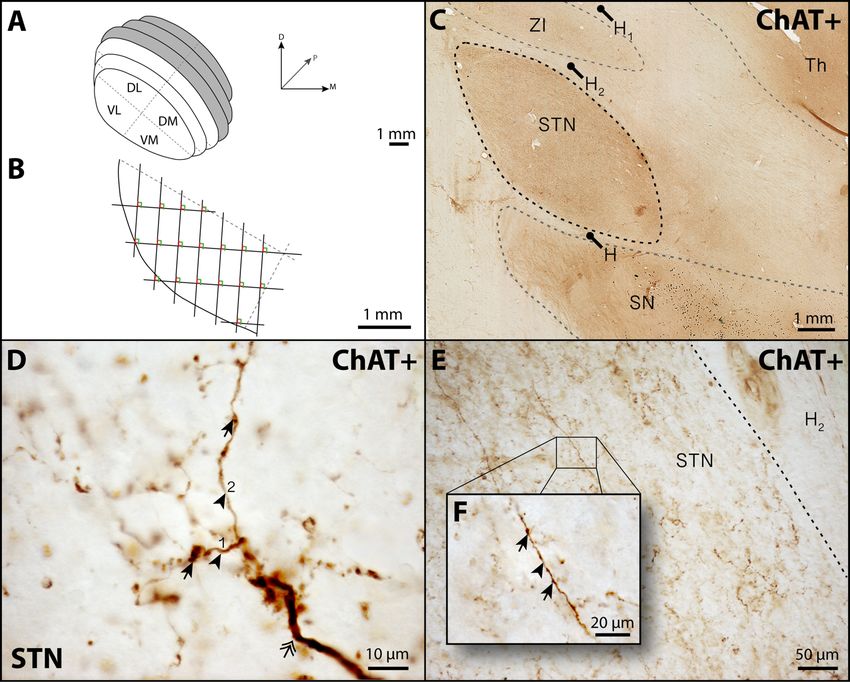
1.2.4.2. Délimitations anatomiques
Le NST est perçu comme un noyau fermé sur toute sa surface, à l’exception de sa partie
médiane qui ne démontre pas de bordure stricte et semble au contraire fusionner avec la
région latérale de l’hypothalamus. Cette vision est supportée par la présence de faisceaux de
fibres myélinisées sur son pourtour : sur sa face ventrale, il est longé par les champs de Forel
(H) et les pédoncules cérébraux alors que sur sa face dorsale, il est juxtaposé antérieurement
à la zona incerta (ZI) et postérieurement au faisceau lenticulaire (H2) ; latéralement, c’est la
capsule interne qui s’interpose entre le NST et les segments pallidaux (Fig 1.2A).
7Figure 1.2. Anatomie et organisation topographique du noyau sous-thalamique. A : Le noyau
sous-thalamique (STN) est délimité dorsalement par le faisceau lenticulaire (H2) appartenant
au champ de Forel. Sa limite latérale est séparée des segments pallidaux (GPe et GPi) et du
putamen (Put) par la capsule interne (IC). B : Le noyau sous-thalamique est divisé en trois
régions fonctionnelles le long de ses axes dorso-ventral, latéro-médian et rostro-caudal.
Modifiée de Hamani et al. (2004).
L’étude de Hardman et ses collègues est la première à proposer une comparaison détaillée
des structures des GB chez le rat, le marmoset, le macaque, le babouin et l’humain
(hamaniHardman et al., 2002). Le NST était facilement délimité dans le cerveau de toutes
ces espèces avec un marquage intense à la parvalbumine, une protéine liant le calcium et qui
est fortement exprimée dans ses neurones. La forme caractéristique du NST, telle que décrite
plus haut par Luys, et son orientation (dorso-latérale à ventro-médiane) semblent avoir été
conservées dans sa phylogénèse. Le NST est, en proportion, la plus petite structure des GB
et ce, chez les primates et non-primates. Des différences quant à la proportion de la densité
neuronale globale ont par ailleurs été notées entre les humains et les autres espèces. Cette
densité neuronale chez l’humain en comparaison aux autres espèces se traduit par un
recrutement plus important des neurones de relais dans la circuiterie des GB et impose
conséquemment une divergence fonctionnelle (Hamani et al., 2004).
81.2.4.3. Phénotypes neuronaux
La population neuronale la plus influente du NST utilise le glutamate comme principal
neurotransmetteur. Morphologiquement, il existe trois sous-classes de neurones, dont deux
appartiennent à la classe des neurones de Golgi de type I (neurones de projection). Ces deux
types de neurones de projection se distinguent l’un de l’autre par la forme de leur corps
cellulaire, qui peut être rond ou au contraire, fusiforme et allongé, ainsi que l’étendue de leur
arborisation dendritique, qui diffère en termes de diamètre et de nombre de branchements
(Pearson, Norris et Phelps, 1985). Ces neurones relâchent du glutamate dans leurs cibles et
comptent pour plus de 90% de la population neuronale contenue dans le NST humain
(Levesque et Parent, 2005). Le NST est donc l’unique composante excitatrice des GB et c’est
justement pour cette raison qu’il est considéré comme une force propulsive au sein de
l’organisation anatomique et fonctionnelle des GB.
La troisième sous-classe neuronale a été considérée comme une population neuronale à part
entière un peu plus tard, avec des travaux qui ont permis de la classifier selon ses
caractéristiques morphologiques distinctives (Rafols et Fox, 1976; Yelnik et Percheron,
1979). Ces neurones présentent une aire somatique plus modeste, des extensions dendritiques
plus restreintes et qui sont surtout concentrées autour du corps cellulaire. L’ensemble de ces
observations a donné naissance à la famille des interneurones du NST, dont la nature
GABAergique a éventuellement été confirmée chez l’humain (Levesque et Parent, 2005).
1.2.4.4. Connexions anatomiques
Le NST est une structure sous-corticale contenant des neurones dont le mode de
communication se fait via la relâche de glutamate. Le NST est donc la seule source excitatrice
des GB et projette massivement vers les structures de sortie des GB. La figure 1.3 illustre les
principales projections qui émanent du NST (à gauche) ainsi que celles qui y convergent (à
droite). Ce dernier schéma évoque notamment la diversité des projections afférentes, faisant
référence à la DA, l’ACh, la sérotonine (5-HT), le GABA et le glutamate. Pour cette raison,
les neurones du NST sont habités, au niveau post-synaptique, par une grande variété de
récepteurs. Chez le macaque, le récepteur D5, partageant une plus forte affinité pour la DA
9que le récepteur D1, semble prédominer (Galvan et al., 2014). Son activation engendre une
augmentation de la fréquence d’apparition des trains de décharge, qui pourrait lui valoir une
implication fonctionnelle dans certaines conditions pathologiques (déplétion en DA, par
exemple). Les sous-types de récepteurs de 5-HT, 5-HT1A, 5-HT2C et 5-HT4, jouent également
un rôle dans les niveaux d’excitabilité des neurones glutamatergiques du NST (Miguelez,
Morera-Herreras, Torrecilla, Ruiz-Ortega et Ugedo, 2014). Le récepteur NMDA exprimant
la sous-unité GluN2D, quant à lui, est retrouvé à des concentrations considérablement élevées
et régule positivement l’amplitude du courant excitateur post-synaptique (Swanger et al.,
2015). Nous mentionnerons au passage la présence des récepteurs ACh dans cette structure
d’intérêt, dont le rôle et la nature seront divulgués dans la prochaine section de ce chapitre.
Finalement, le NST entretient une somatotopie fonctionnelle qui est fortement dépendante
des projections qu’il reçoit en provenance de différentes régions corticales (Coude, Parent et
Parent, 2018; Parent et Hazrati, 1995b). Le NST peut donc être réparti en un secteur
sensorimoteur dans sa région dorsolatérale, un secteur associatif dans sa région ventrale et
un secteur limbique au niveau de sa pointe médiane (Fig. 1.2B).
Figure 1.3. Vue d’ensemble des principales connexions anatomiques existant au sein du
noyau sous-thalamique. L’épaisseur des flèches dépeint l’envergure des projections, une
flèche plus épaisse correspondant à des projections plus importantes. Abréviations :
VTA = Aire tegmentaire ventrale, GPe = Globus pallidum externe, GPi = Globus pallidus
interne, FRM = Formation réticulaire mésencéphalique, FRP = Formation réticulaire
10pontique, NRD = Noyau du raphé dorsal, PPN = Noyau pédonculopontin, SI = Substantia
innominata, SNc = Substance noire pars compacta, SNr = substance noire pars réticulata.
1.3. L’acétylcholine dans le système nerveux central
Les facultés cognitives sont médiées par un large éventail de processus électriques et
chimiques au sein des systèmes nerveux central et périphérique. La communication
neuronale s’articule autour de la conversion d’un influx nerveux électrique à la relâche de
neurotransmetteurs dans la fente synaptique, dont l’action spécifique est déterminée par les
familles de récepteurs exprimées en grande partie au niveau de la membrane post-synaptique.
On retrouve une grande variété de neurotransmetteurs dans le cerveau et tous exercent des
rôles fondamentaux dans l’exécution de fonctions qui ont trait à la cognition, la motricité, les
perceptions sensorielles, les états de vigilances et le tout, pour le bon fonctionnement
neuronal. Certains auront des effets excitateurs (glutamate) ou inhibiteurs (GABA et
glycine), alors que d’autres agiront différemment en fonction du récepteur sur lequel se lie le
neurotransmetteur (c’est le cas notamment de la dopamine (DA), de la sérotonine (5-HT) et
de l’ACh). L’emphase sera ici portée sur le rôle de l’ACh dans le système nerveux central,
mais aussi sur ses interactions avec les autres systèmes chimiques qui s’influencent
mutuellement dans certaines circonstances.
1.3.1. Origine des afférences cholinergiques
L’ACh est exprimée dans plusieurs structures du cerveau et son implication a été soulevée
dans les voies ascendantes et descendantes. Ces structures ACh ont été classifiées par
Mesulam et ses collaborateurs (Ch1 à Ch6) : le noyau septal médian (Ch1), le noyau du
membre vertical de la bandelette diagonale de Broca (Ch2), le noyau du membre horizontal
(latéral) de la bandelette diagonale de Broca (Ch3), le noyau basalis de Meynert (Ch4), le
noyau tegmentaire pédonculonpontin (PPN, Ch5) et le noyau tegmentaire latérodorsal (LDT,
Ch6) (Mesulam et al., 1983). Les secteurs Ch4 à Ch6 recevront une attention singulière dans
11ce chapitre, notamment en raison de leur connectivité réciproque avec les GB (Mesulam et
al., 1992).
1.3.1.1. Noyau basalis de Meynert (Ch4)
Le noyau basalis de Meynert (nbM), situé dans le prosencéphale, dorsalement au nerf
optique, est la source primaire d’ACh vers le cortex cérébral et l’amygdale (Mesulam et al.,
1983). De son innervation en direction des régions corticales émergea l’hypothèse
cholinergique, à laquelle on accorde encore aujourd’hui une grande importance dans le déclin
cognitif de la population gériatrique (Bartus, Dean, Beer et Lippa, 1982). Étant donné que
l’étude de ce noyau et de ses projections ne fait pas l’objet de ce projet de mémoire, il ne sera
pas considéré en détail dans cette section. En revanche, son implication sera à nouveau
évoquée dans l’origine de la dégénérescence ACh dans la MP.
1.3.1.2. Noyaux tegmentaires pédonculopontin (Ch5) et dorsolatéral (Ch6)
Le PPN et le LDT sont deux structures situées près de la jonction pontomésencéphalique et
dont les délimitations anatomiques ne sont parfois pas clairement définies (Mesulam et al.,
1983). Toutefois, dans la littérature, un consensus règne sur la question de leurs principales
bordures anatomiques. Pour le PPN, elles ont été définies comme suit : il est longé par le
lémnisque médian sur sa face latérale ; médialement, par la décussation du pédoncule
cérébelleux supérieur ; dorsalement, il est rejoint par le champ rétrorubral sur sa face
antérieure et par le noyau cunéiforme sur sa face postérieure ; ventralement, il est circonscrit
par la formation réticulaire pontique. Finalement, ses neurones peuvent s’apposer à ceux de
la SN et du locus cœruleus (Steckler, Inglis, Winn et Sahgal, 1994). Le LDT est quant à lui
situé en périphérie de la substance grise périaqueducale.
Contrairement au LDT, le PPN démontre un gradient de concentration de neurones ACh qui
s’étend de façon croissante sur son axe rostro-caudal. Ainsi, sa région rostrale réfère à la
division diffuse (pars dissipita) alors que sa région caudale correspond à la région compacte
(pars compacta). Des neurones arborant d’autres profils neurochimiques ont aussi été
12identifiés ; notons à cet égard la présence de neurones GABAergiques et glutamatergiques,
et une proportion de neurones co-exprimant l’ACh et le glutamate (Clements, Toth, Highfield
et Grant, 1991; Lavoie et Parent, 1994a). Bien qu’ils puissent opérer de manière sensiblement
différente, le PPN et le LDT, regroupant les neurones appartenant aux classes Ch5 et Ch6 du
système ACh central, respectivement, composent le système réticulaire activateur ascendant
(SRAA), connu pour contribuer à un grand nombre de fonctions essentielles à la survie,
incluant le contrôle moteur et les divers états de vigilance et de conscience (Meyer, 1970;
Vertes et Martin, 1988).
Ces deux regions du tronc cérébral envoient la plus grande partie de leurs terminaisons
axonales ACh au thalamus, plateforme pivot qui promeut à son tour l’action indirecte de ces
derniers sur la régulation de l’activité corticale (Moruzzi et Magoun, 1949). Ils entretiennent
d’autres connexions ascendantes avec plusieurs composantes des GB (striatum, SNc, GPi et
STN), le système limbique (noyau accumbens) et les collicules supérieurs, en plus de
projections vers la moelle épinière pour un contrôle fin du patron de locomotion (Gut et
Winn, 2016; Wang et Morales, 2009).
Pour l’heure, la place du PPN dans les nombreux systèmes fonctionnels est sans cesse
investiguée et revisitée. Alors qu’il était de prime abord considéré pour ses fonctions dans le
SRAA, la recherche sur son rôle dans l’attention et la cognition a connu récemment une forte
émergence. Plus encore, son implication dans la locomotion ne serait pas seulement confinée
à l’exécution d’une tâche motrice, mais à la sélection d’une action (Gut et Winn, 2016). En
d’autres termes, le PPN est capable de produire, de concert avec les autres acteurs des GB,
une réponse rapide à l’arrivée d’un stimulus. Ses projections réciproques avec le NST, qui
reçoit des connexions en provenance du cortex (voie hyper-directe, Fig. 1.1), rendent compte
de son rôle dans la sélection rapide des commandes motrice, associative et limbique.
1.3.2. Caractéristiques de l’innervation cholinergique du noyau sous-thalamique
Les premières traces de l’existence d’une biologie cholinergique datent du tout début du 20e
siècle (1906) et proviennent de travaux effectués par Reid Hunt (1870-1948) et René de
13Mortemer Taveau (1876-date de décès inconnue). Ces derniers ont noté un effet de la choline
et ses dérivés (notamment l’ACh) sur la réduction globale de la pression artérielle chez des
lapins. Suite à cette découverte, deux autres collègues, Otto Loewi (1873-1961) et Henry
Dale (1875-1968), ont également contribué de manière significative à la compréhension de
la neurotransmission cholinergique. En effet, Loewi a été le premier à émettre l’hypothèse
de la nature chimique du message neuronal par le biais d’une expérience des plus élégantes
dont l’idée lui est venue au cours de son sommeil. Le paradigme expérimental consistait à
placer un cœur de grenouille avec son nerf vague intact dans une chambre de perfusion et un
cœur de grenouille dénervé dans une autre chambre, avec son propre perfusat. Dans la
première condition, une stimulation du nerf vague provoquait un ralentissement de la
fréquence cardiaque (résultat attendu, car ce phénomène était déjà bien connu à l’époque).
La découverte s’est proclamée quand Loewi a transféré la solution de perfusion du cœur
innervé dans la deuxième chambre. Effet remarquable : la réduction de la fréquence
cardiaque était également perçue au niveau du coeur dénervé. Quant à lui, Dale a démontré
que l’effet transitoire de l’ACh sur sa cible était dû à l’action d’une estérase présente dans le
milieu qui métabolise l’ACh. Grâce à ces travaux de grande envergure, ils se sont vus tous
les deux décerner le prix Nobel de la Médecine en 1936. Ce n’est pas avant 1968, en
revanche, que l’on a eu la confirmation de la présence de cette molécule au sein du SNC,
chez le rat. Plus tard, et encore aujourd’hui d’ailleurs, un intérêt particulier est porté sur les
caractéristiques ultrastructurales de l’innervation ACh qui nous pourvoit d’informations
supplémentaires, essentielles à une compréhension raffinée de son fonctionnement
moléculaire et son rôle dans le cerveau. Un nombre d’évidences, telles que décrites grâce à
l’usage de la microscopie électronique à transmission, suggère que l’ACh exerce une
influence volumique sur ses cibles, c’est-à-dire que sa relâche au niveau des varicosités
axonales (structure qui définit un regroupement de vésicules qui contiennent les
neurotransmetteurs) est largement asynaptique (absence de spécialisation membranaire
synaptique, qui se traduit notamment par l’absence d’une densité post-synaptique, Fig. 1.4)
(Descarries et Mechawar, 2000). Ceci implique que l’ACh a un effet diffus sur son
environnement et peut manifestement atteindre des cibles éloignées (situées dans une sphère
dont le rayon avoisine les 10 µm), expliquant entre autres la grande diversité de récepteurs
ACh exprimés dans le cerveau.
14Vous pouvez aussi lire

















































