LES CHAMPIGNONS PARASITES (CHYTRIDES) DANS LES LACS D'AUVERGNE
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Serena Rasconi
Directeur de thèse: Télesphore Sime-Ngando
LES CHAMPIGNONS PARASITES (CHYTRIDES) DANS LES LACS D’AUVERGNE
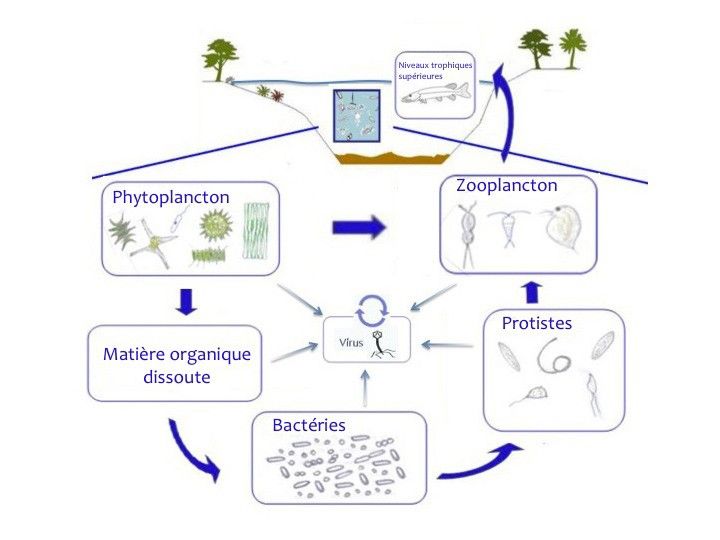
Un lac est un écosystème complexe, comprenant une zone littorale où les interactions avec les
sédiments sont importantes, et une zone strictement aquatique, dite zone pélagique. Dans cette
dernière zone, la biodiversité est très importante et principalement représentée par des virus et des
organismes microscopiques (phytoplancton, zooplancton et autres micro-organismes). Ces
microorganismes, via les multiples interactions qui les relient, gouvernent le fonctionnement de
l’écosystème lacustre entier. Plus en détail, les micro-algues (phytoplancton) réalisent la
photosynthèse et constituent les producteurs primaires de la matière organique. Cette nouvelle
biomasse produite peut être consommée directement par le zooplancton via la prédation, ou
également en tant que matière organique dissoute, produite sous forme d’exsudats excrétés suite à
la photosynthèse, et qui constituent la ressource nutritive primaire des bactéries dites hétérotrophes.
Ces bactéries sont ensuite consommées par des microorganismes bactérivores de plus grande taille
(protistes), qui sont à leur tour les proies du zooplancton métazoaire. L’ensemble de ces
microorganismes pélagiques forment ainsi une chaine trophique complexe (dite microbienne;
Pomeroy 1974, Azam et al 1983) dont le fonctionnement est essentiel au transfert de matière et
d’énergie entre maillons trophiques, grâce au rôle clé des prédateurs (Fig. 1).
Fig. 1 La chaine trophique microbienne dans les milieux pélagiques et les différents microorganismes impliqués.
En raison de leur taille, ces microprédateurs sont difficiles à étudier à l’aide de méthodes
microscopiques classiques. En conséquence, la connaissance de leur diversité et de leurs rôles
écologiques dans le fonctionnement des écosystèmes aquatiques a longtemps été limitée. Ces
limites méthodologiques sont en train d’être repoussées avec le développement des techniques de
la biologie moléculaire, notamment celles des d’empreintes génétiques qui, aujourd’hui, permettent
de relier identité et fonction chez les microorganismes non cultivables. Il faut préciser que ces
microorganismes non cultivables représentent l’essentiel (plus de 99%) des microorganismes de
notre environnement. On peut ainsi déterminer des microorganismes inconnus par affiliation
1
phylogénétique de leurs séquences ADN, par rapport à celles de micro-organismes déjà décrits
présentes dans les bases de données moléculaires internationales.
C’est ainsi que des études récentes, initiées dans les lacs d’Auvergne (Lefranc et al 2005, Lepère et
al 2008, Lefèvre et al 2007, 2008), ont permis de se rendre compte de la diversité et des fonctions
écologiques potentielles associées à la communauté des protistes hétérotrophes qui, jusque là,
étaient tous considérés comme prédateurs des bactéries dans les modèles de fonctionnement de la
chaine trophique microbienne des écosystèmes aquatiques. Un de résultat le plus remarquable de
ces études a été la mise en évidence de l’omniprésence de champignons (Eumycètes)
microscopiques, dont le sous-groupe dominant était celui des Chytridiomycètes (chytrides).
Les travaux effectués au cours de cette thèse, ont clairement démontré l’omniprésence, la diversité
et l’importance quantitative et fonctionnelle des chytrides dans les écosystèmes lacustres. Ces
champignons sont caractérisés par un cycle de vie complexe, considéré comme une adaptation au
mode de vie pélagique, puisqu’il comporte un stade infectieux, le sporange, qui se développe fixé
sur les algues planctoniques qui constituent leurs hôtes dont ils se nourrissent. Au terme de sa
croissance, le sporange produit les stades de dissémination constitués par des zoospores
flagellées, dont les flagelles assurent la mobilité nécessaire à la recherche d’un nouvel hôte en
milieu aqueux.
Dans la littérature scientifique, les chytrides sont décrits comme des parasites d’algues. Ils ont été
observés dès le milieu du siècle dernier à l’aide de techniques de microscopie classique (Canter
1950, 1953a, b, 1972) et, relativement plus récemment, par microscopie électronique (Beakes et al.
1992a, b, 1993). Leur présence a été rapportée dans des aires biogéographiques contrastées (Kobb
1966, Van Donk et Ringelberg 1983, Sen 1987a, b) et serait caractéristique des milieux lacustres.
Cependant, ces études sont essentiellement descriptives (morphologie et cycle de développement
des espèces), et ne resituent pas le rôle des chytrides dans le fonctionnement des réseaux
trophiques microbiens. En fait, des outils de routines adaptées à l’étude du rôle de ces organismes
font défaut dans le contexte de l’écologie aquatique, en raison de leur petite taille et de la complexité
des formes de vie.
C’est ainsi que dans un premier temps, les objectifs de la thèse ont été donc d’utiliser les
technologies récentes pour développer de nouveaux outils permettant d’identifier, de quantifier et de
déterminer la prévalence et l’intensité d’infection des chytrides parasites du phytoplancton dans les
écosystèmes aquatiques, en tenant compte des différentes formes de vie. Des outils adaptés ont
été mis au point en couplant les méthodes de microscopie et de la biologie moléculaire.
Pour l’étude de la phase parasitaire (les sporanges), nous avons testé un colorant spécifique de la
chitine qui est un composant de la paroi cellulaire des champignons. Après excitation sous ultra-
violet, ce permet d’identifier et de distinguer les chytrides des autres micro-organismes présents
dans le milieu (Fig. 2). Une estimation quantitative rapide et fiable de ces parasites dans des
échantillons environnementaux a ainsi pu être réalisée. De plus, le protocole proposé (Rasconi et al
2009) permet également une saisie d’images de bonne qualité (Fig. 3), exploitables pour la
détermination à la fois des parasites et de leurs hôtes à partir de critères morphologiques.
Fig.2. La méthode de coloration développée permet de distinguer facilement les champignons parasites des autres
organismes, grâce à la forte fluorescence bleue.
2
Fig. 3. Le protocole mis au point pour la détection et la quantification des sporanges de chytrides parasites a fait l’objet
d’une publication dans une revue scientifique (Rasconi et al 2009) et une des photos prise au microscope a été retenue
pour la couverture du numéro où l’article est paru.
3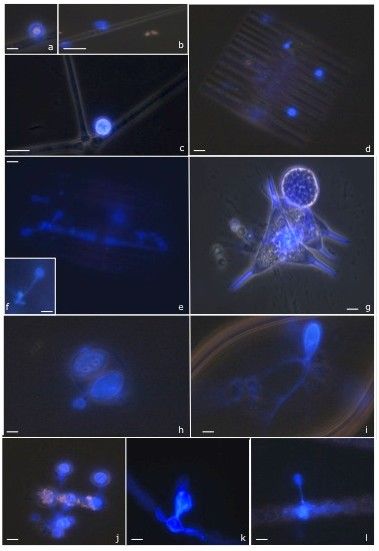
Contrairement aux sporanges, le stade de dissémination est dépourvu de chitine et ne possède pas
de composants cibles pour des colorants spécifiques. De plus, ces propagules sont de très petite
taille et ne présentent pas de caractéristiques morphologiques pouvant permettre de les différencier
d’autres microorganismes flagellés présents dans le milieu aquatique. Les outils moléculaires offrent
une alternative pour pallier à ces inconvénients. C’est ainsi que nous avons mis au point une sonde
moléculaire spécifique des chytrides, fonctionnant par hybridation d’un brin complémentaire de
l’ARN ribosomal des chytrides présent dans l’environnement (Fig. 4). L'ARN ribosomal est une
molécule hautement conservée, présente dans les cellules de tous les organismes, qui est produite
à partir d’une matrice d’ADN. Avec cette technique de marquage fluorescent par la sonde
moléculaire mise au point, a été possible d’identifier et de compter les zoospores au microscope
(travail effectué en collaboration avec M. Jobard, Jobard et al 2010) (Fig. 5).
Fig. 4. La méthode de coloration utilisée rend fluorescentes les zoospores des champignons qui deviennent ainsi visibles
sous la lumière du microscope.
Fig. 5. Le protocole mis au point pour la détection et la quantification des zoospores de champignons parasites a fait l’objet
d’une publication dans une revue scientifique (Jobard et al 2010).
Une fois les outils méthodologiques développés et validés, l’objectif de l’étude a été de décrire la
dynamique saisonnière et l’incidence des communautés de chytrides parasites sur le
développement des successions du phytoplancton lacustre. Deux lacs contrastés ont fait l’objet de
ces suivis: le lac Pavin qui est un lac profond et relativement pauvre en nutriments (lac oligo-
mésotrophe), et le lac d’Aydat qui est un lac peu profond et riche en nutriments (lac eutrophe).
L’étude a été conduite en 2007 et les échantillons ont été prélevés tous les 15 jours au point central
des lacs.
4
Pour ce qui concerne l’étude de la phase parasitaire, l’exploitation de la méthode de coloration
développée et des clés taxonomiques disponibles dans les ouvrages de référence (Sparrow 1960,
Canter 1950, Canter and Lund 1951) ont permis d’identifier les espèces rencontrées sur des bases
morphologiques et de mettre en évidence que plusieurs espèces d’algues étaient infectées dans les
lacs étudiés (Fig. 6).
Fig. 6. Photographies de chytrides (stade sporanges) parasitant des algues prises au microscope pendant le suivi
saisonnier dans le lac Pavin et le lac d’Aydat. L’observation des champignons parasites a abouti à l’identification sur bases
morphologiques de la plupart des espèces présentes dans les échantillons récoltés, ainsi que des différentes phases de
leur cycle de vie. Fig. a-f différents stade de développement des parasites sur des algues diatomées dans le lac Pavin (a-
d) et le lac d‘Aydat (e, f). Fig. g-h exemple de Chlorophytes parasitées dans le lac Pavin (stade mature); Fig. i-l exemple de
Cyanobactéries parasitées dans le Lac d’Aydat (i-k stade mature, l début de l’infection). Echelle 10 µm.
5
Ce travail a permis de relier la dynamique des parasites à celle de leurs hôtes et d’évaluer
l’incidence du parasitisme en utilisant des paramètres tels que la proportion d’individus infectés dans
une population (prévalence de l’infection) et le rapport entre le nombre de parasites et le nombre
d’individus infectés dans la population hôte (intensité de l'infection). La comparaison des
dynamiques saisonnières a montré que la période de présence maximale des chytrides dans le lac
Pavin est le printemps et que le groupe d’algues le plus attaqué par les parasites est celui de
diatomée. Dans le lac d’Aydat les parasites de diatomées sont également plus abondant au
printemps. Cependant, le pic d’abondance des chytrides a été observé en automne, lors de la
prolifération des Cyanobactéries. Dans les deux lacs, au moment de l’année ou les parasites
prolifèrent, ils peuvent influencer et limiter le développement des populations phyoplanctoniques
hôtes, en changeant, profondément, la structure saisonnière des peuplements phytoplanctoniques
qui est à la base de l’organisation biologique des écosystèmes lacustres.
La dernière partie de ce travail a été consacrée à l’étude de la phase de dissémination des
chytrides, à l’aide des outils moléculaires mises au point. Les zoospores des chytrides ont pu être
visualisés et leurs dimensions évaluées, démontrant une forte homogénéité dans les deux lacs
étudiés. De plus, tout comme pour la phase parasitaire, la dynamique saisonnière a été déterminée
à partir d’échantillons naturels. La comparaison entre l’abondance des zoospores et celles des
sporanges a mis en évidence l’importance des zoospores comme sources de nouvelles infections
du phytoplancton et comme ressources nutritives pour le zooplancton.
Un des résultats remarquables et récurrent dans les deux lacs a été l’importance de la taille des
hôtes pour le développement de l’infection, aussi pour la phase parasitaire que pour les zoospores.
Une corrélation positive a été observée entre la taille moyenne des espèces hôtes et le pourcentage
des individus infectés dans la population. Les comptages et observations ont également fait ressortir
que la production de zoospores (nombre de zoospores / sporange) était également positivement
corrélée à la présence d’algues de grandes tailles. Ces résultats impliquent que la miniaturisation
chez les hôtes constitue un véritable refuge contre les chytridiomycoses.
D’un point de vue écologique, la préférence de parasites des algues de grandes tailles peut avoir
des conséquences importantes pour le fonctionnement des écosystèmes lacustres et les cycles
biogéochimiques associés. Du fait de leur taille, ces algues sont peut utilisées comme proies par le
zooplancton, ce qui limite l’efficacité du transfert de carbone de ce compartiment algal vers les
niveaux trophiques supérieurs. Ainsi, dans les modèle contemporains du fonctionnement des
écosystèmes aquatiques, leur biomasse sédimenterait et serait donc, en grande majorité, ‘perdue’
pour les processus pélagiques. Ce travail a apporté la preuve que les parasites utilisent une partie
de ces algues non seulement comme ressource nutritive, mais aussi pour la production de
zoospores qui sont des ressources nutritives de qualité pour le développement du zooplancton
métazoaire. En effet, on sait que les zoospores de chytrides sont riches en éléments essentiels,
notamment en acides gras polyinsaturés et en cholestérol. Nos recherches aboutissent donc à une
nouvelle vision du fonctionnement des écosystèmes aquatiques, et ont été depuis reprises par
différents laboratoires dans le monde (Nascimento et al 2010, Gleason et al 2010, Marano et al
2011). Contrairement aux idées reçues, les efflorescences phytoplanctoniques, dont certaines sont
nuisibles comme celles des cyanobactéries filamenteuses, ne seraient donc pas que de simples
impasses trophiques. Nos travaux ouvrent donc des perspectives intéressantes en apportant une
alternative biologique aux traitements chimiques de ces efflorescences. En effet, nous avons pu
montrer, récemment, que ces traitements, notamment par le sulfate de cuivre, avait des effets
néfaste sur l’ensemble des maillons et liens trophiques de l’écosystème lacustre (Le Jeune et al
2006, 2007).
6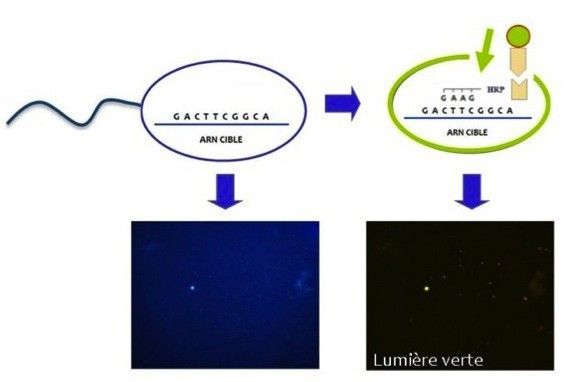
Fig. 7. Une synthèse bibliographique sur le rôle des champignons dans le réseau trophic microbien lacustre a été rédigée
pendant la thèse et acceptée pour être publiée dans une revue scientifique (Rasconi et al 2011). De suite une
schématisation du rôle des chytrides dans le réseau trophique aquatique.
Pour conclure, les développements méthodologiques récents, tels que les techniques de biologie
moléculaire, ont amélioré considérablement notre connaissance de la composition taxonomique des
organismes cellulaires de petite taille. Cela a permis de ‘redécouvrir’ la présence des Eumycètes
dans les milieux pélagiques lacustres, où les Chytridiomycètes ont été reconnus comme le groupe le
plus important. Il était donc important de préciser la diversité et l’importance quantitative et
fonctionnelle des ces microorganismes pour les replacer dans le contexte des réseaux trophiques
microbiens dans les écosystèmes aquatiques. En effet, une connaissance approfondie de la
diversité et de la structure des communautés biologiques est nécessaire pour améliorer la
compréhension des processus écologiques qui sont à la base du fonctionnement des écosystèmes
naturels. A partir de là, il est important de noter que l’association de la biologie moléculaire à un
savoir-faire permettant des analyses de qualité en microscopie, ainsi qu’une expérience de
systématicien (spécialiste de la reconnaissance d’espèces à partir de critères morphologiques), aura
permis d’aller au-delà d’études simplement descriptives.
L’ensemble de ce travail, mené par une approche écosystémique, a donc permis d’intégrer les
chytrides dans le fonctionnement des milieux pélagiques et d’évaluer l’incidence du parasitisme
comme stratégie trophique dans le contexte général des réseaux trophiques microbiens. Ainsi, avec
cette vision renouvelée des écosystèmes aquatiques, les champignons parasites ne constituent pas
seulement un facteur de régulation de leurs hôtes, mais représentent une véritable force écologique
dans le fonctionnement de l’écosystème.
7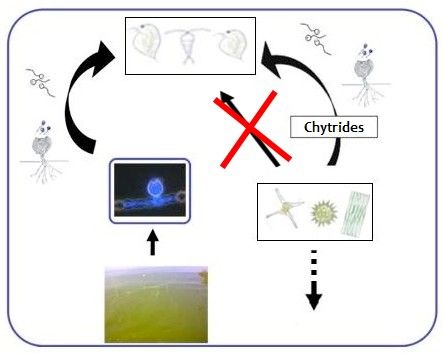
Références bibliographiques
Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F. 1983. The Ecological Role of Water-Column Microbes
in the Sea. Marine Ecology Progress Series. 10:257-263.
Beakes GW, Canter HM, Jaworski GHM. 1992a. Comparative ultrastructural ontogeny of zoosporangia of Zygorhizidium
affluens and Z. planktonicum, chytrid parasites of the diatom Asterionella formosa. Mycological Research 96:1047-1059.
Beakes GW, Canter HM, Jaworski GHM. 1992b. Ultrastructural study of operculation (discharge apparatus) and zoospore
discharge in zoosporangia of Zygorhizidium affluens and Z. planktonicum, chytrid parasites of the diatom Asterionella
formosa. Mycological Research 96:1060-1067.
Beakes GW, Canter HM, Jaworski GHM. 1993. Sporangium differentiation and zoospore fine-structure of the chytrid
Rhizophydium planktonicum, a fungal parasite of Asterionella formosa. Mycological Research 97:1059-1074.
Canter HM, Lund JWG 1951. Studies on plankton parasites. III. Examples of the interaction between parasitism and other
factors determining the growth of diatoms. Annals of Botany 15:359-371.
Canter HM. 1950. Fungal parasites of the phytoplankton .I. Studies on British chytrids X. Annals of Botany 14:263-289.
Canter HM. 1953a. Annotated list of British aquatic chytrids. Transactions of the British Mycological Society 36:278-303.
Canter HM. 1953b. Fungal parasites of the phytoplankton. I. Annals of Botany 14:263-289.
Canter HM. 1972. A guide to the fungi occurring on planktonic blue-green algae. Pages 145-159, in Taxonomy and Biology
of Blue-Green Algae, TV Desikachary Ed. University of Madras, Madras, India.
Gleason FH, Schmidt SK, Marano AV. 2010. Can zoosporic true fungi grow or survive in extreme or stressful
environments?. Extremophiles 14:417-425.
Jobard M, Rasconi S, Sime-Ngando T. 2010. Fluorescence in situ hybridization of zoosporic fungi (chytrids) in aquatic
environments: testing with clone-FISH and application to natural samples using CARD-FISH. Journal of Microbiological
Methods 83, 236-43.
Koob DB. 1966. Parasitism of Asterionella formosa Hass by a chytrid in two lakes of Rawah wild area of Colorado. Journal
of Phycology 2:41-45.
Le Jeune AH, Charpin M, Deluchat V, Briand JF, Lenain JF, Baudu M, Amblard C. 2006. Effect of copper sulphate
treatment on natural phytoplanktonic communities, Aquatic Toxicology 80:267-280.
Le Jeune AH, Charpin M, Sargos D, Lenain JF, Deluchat V, Ngayila N, Baudu M, Amblard C. 2007. Planktonic microbial
community responses to added copper. Aquatic Toxicology 83:223-237.
Lefèvre E, Bardot C, Noel C, Carrias JF, Viscogliosi E, Amblard C, Sime- Ngando T. 2007. Unveiling fungal zooflagellates
as members of freshwater picoeukaryotes: evidence from a molecular diversity study in a deep meromictic lake.
Environmental Microbiology 9:61-71.
Lefèvre E, Roussel B, Amblard C, Sime-Ngando T. 2008. The molecular diversity of freshwater picoeukaryotes reveals
high occurrence of putative parasitoids in the plankton. PLoS One 3:e2324.
Lefranc M, Thenot A, Lepere C, Debroas D. 2005. Genetic diversity of small eukaryotes in lakes differing by their trophic
status. Applied and Environmental Microbiology 71:5935-5942.
Lepere C, Domaizon I, Debroas D. 2008. Unexpected importance of potential parasites in the composition of the
freshwater small-eukaryote community. Applied and Environmental Microbiology 74:2940-2949.
Marano AV, Pires-Zottarelli CLA, Barrera MD, Steciow MM, Gleason FH. 2011. Diversity, role in decomposition, and
succession of zoosporic fungi and straminipiles on submerged decaying leaves in a woodland stream. Hydrobiologia
659:93-109.
Nascimento CA, Gomes EPC, Pires-Zottarelli CLA. 2010. Occurrence and distribution of zoosporic organisms in water
bodies from Brazilian Cerrado areas. Mycologia 10:139v1-10139.
Pomeroy LR. 1974. The Ocean's Food Web, A Changing Paradigm. BioScience 24:99-504.
Rasconi S, Jobard M, Jouve L, Sime-Ngando T. 2009. Quantitative ecology of chytrid epidemics: Use of calcofluor white for
diagnosing, identifying and counting phytoplanktonic fungal parasites. Applied and Environmental Microbiology 75:2545-
2553.
Rasconi S, Jobard M, Jouve L, Sime-Ngando T. 2011. Parasitic fungi of lake phytoplankton: Ecological role and
implications for freshwater microbial food webs. Aquatic Microbial Ecology. 62:123-137.
8Sen B. 1987a. Fungal parasitism of planktonic algae in Shearwater. I. Occurrence of Zygorhizidium affluens Canter on
Asterionella formosa Hass in relation to the seasonal periodicity of the alga. Archiv für Hydrobiologie 76:101-27.
Sen B. 1987b. Fungal parasitism of planktonic algae in Shearwater. II. A study of the chytrid parasites of the diatom
Fragilaria crotonensis Kitton. Archiv für Hydrobiologie 76:129-44.
Sparrow FK. 1960. Aquatic phycomycetes, 2nd ed. University of Michigan Press, Ann Arbor, MI.
Van Donk E, Ringelberg J.1983. The effect of fungal parasitism on the succession of diatoms in Lake Maarsseveen (The
Netherlands). Freshwater Biology 13:241-251.
9Vous pouvez aussi lire

















































