EFFETS DE BACTÉRIOCINES SUR LE PATHOGÈNE PORCIN - STREPTOCOCCUS SUIS MÉMOIRE GENEVIÈVE LEBEL - CORPUS UL
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Effets de bactériocines sur le pathogène porcin
Streptococcus suis
Mémoire
Geneviève LeBel
Maîtrise en microbiologie
Maître ès Sciences (M.Sc.)
Québec, Canada
© Geneviève LeBel, 2014
I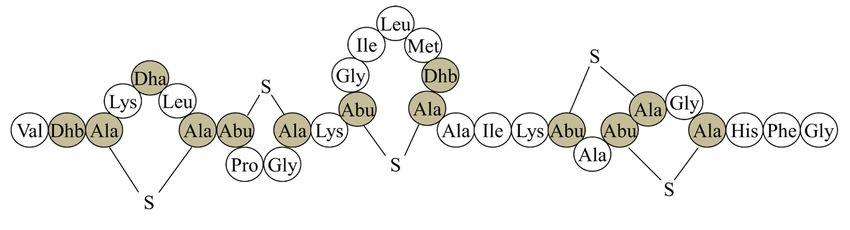
II
RÉSUMÉ
Le but de cette étude était d’évaluer le potentiel antimicrobien de bactériocines pour le
contrôle du pathogène porcin Streptococcus suis sérotype 2. Pour répondre au premier
objectif de ce projet, la susceptibilité de S. suis à la nisine, une bactériocine produite par
Lactococcus lactis, seule ainsi qu’en combinaison avec différents antibiotiques, a été
démontré. Pour réaliser le second objectif, la purification et la caractérisation d’une
bactériocine produite par une souche non-virulente de S. suis sérotype 2 ont été effectuées.
La bactériocine a d’abord été purifiée par HPLC pour ensuite être caractérisée au niveau
biochimique, ainsi que génique. Des études plus approfondies permettront d’évaluer la
capacité de la nisine, de la suicine 90-1330 ou de leur souche productrice respective à
prévenir des infections expérimentales à S. suis chez le porc en vue d'une potentielle
application thérapeutique et préventive.
III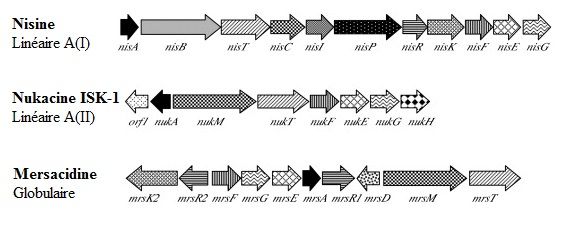
IV
TABLE DES MATIÈRES
RÉSUMÉ .............................................................................................................................. III
TABLE DES MATIÈRES ..................................................................................................... V
LISTE DES FIGURES ....................................................................................................... VII
LISTE DES TABLEAUX ................................................................................................... IX
LISTE DES ABRÉVIATIONS ........................................................................................... XI
REMERCIEMENTS ........................................................................................................... XV
1. INTRODUCTION ....................................................................................................... 1
1.1. Streptococcus suis ............................................................................................... 1
1.1.1. Généralités .................................................................................................... 1
1.1.2. Facteurs de virulence .................................................................................... 3
1.1.3. Pathologies .................................................................................................... 4
1.2. Contrôle des infections à Streptococcus suis ...................................................... 7
1.2.1. Vaccination ................................................................................................... 7
1.2.2. Administration d’antibiotiques ..................................................................... 7
1.2.3. Résistance aux antibiotiques ......................................................................... 8
1.3. Bactériocines ....................................................................................................... 9
1.3.1. Généralités .................................................................................................... 9
1.3.2. Classification .............................................................................................. 10
1.3.2.1. Classe I : lantibiotiques ......................................................................... 11
1.3.2.1.1. Biosynthèse et organisation génique .............................................. 12
1.3.2.1.2. Mode d’action ................................................................................ 13
1.3.2.1.3. Nisine A ......................................................................................... 15
1.3.2.2. Classe II................................................................................................. 17
1.3.2.3. Classe III ............................................................................................... 17
1.3.2.4. Classe IV ............................................................................................... 17
1.3.3. Applications des bactériocines.................................................................... 18
2. PROBLÉMATIQUE .................................................................................................. 19
2.1. Hypothèse de recherche .................................................................................... 19
2.2. Objectifs ............................................................................................................ 19
3. MATÉRIEL ET MÉTHODES................................................................................... 21
3.1. Effet antibactérien de la nisine vis-à-vis S. suis................................................ 21
V3.1.1. Souches bactériennes et conditions de croissance...................................... 21
3.1.2. Activité antagonisme de Lactococcus lactis envers S. suis ........................ 21
3.1.3. Détermination des concentrations minimales inhibitrices (CMIs) et des
concentrations minimales bactéricides (CMBs) ........................................................ 23
3.1.4. Cinétique de l’activité bactéricide de la nisine........................................... 23
3.1.5. Microscopie électronique à transmission de cellules de S. suis traitées à la
nisine 24
3.1.6. Interactions synergiques de la nisine avec des antibiotiques ..................... 24
3.2. Identification, caractérisation et purification d’une bactériocine produite par
une souche non-virulente de S. suis .............................................................................. 25
3.2.1. Souches bactériennes et conditions de croissance...................................... 25
3.2.2. Test de production de bactériocines sur milieu solide ............................... 25
3.2.3. Effet de la source de carbone et de sa concentration sur la production de la
bactériocine par S. suis 90-1330 ................................................................................ 25
3.2.4. Purification de la bactériocine produite par S. suis 90-1330 ...................... 26
3.2.5. Analyse par électrophorèse (SDS-PAGE).................................................. 27
3.2.6. Caractérisation de la bactériocine .............................................................. 27
3.2.7. Perméabilisation de la membrane .............................................................. 28
3.2.8. Spectre d’activité de la bactériocine purifiée ............................................. 29
3.2.9. Séquençage des acides aminés ................................................................... 29
3.2.10. Identification du groupe de gènes putatifs codant pour la bactériocine de
S. suis 90-1330 .......................................................................................................... 30
3.2.11. Distribution du gène sslA chez les souches de S. suis ................................ 30
3.2.12. Évaluation in vitro de la sécurité de S. suis 90-1330 ................................. 31
4. RÉSULTATS ............................................................................................................ 33
4.1. Effet antibactérien de la nisine vis-à-vis S. suis ............................................... 33
4.2. Identification, caractérisation et purification d’une bactériocine produite par la
souche non-virulente de S. suis 90-1330....................................................................... 39
5. DISCUSSION ........................................................................................................... 55
5.1. Effet antibactérien de la nisine vis-à-vis S. suis ............................................... 55
5.2. Identification, caractérisation et purification d’une bactériocine produite par la
souche non-virulente de S. suis 90-1330....................................................................... 58
6. CONCLUSION ......................................................................................................... 65
7. BIBLIOGRAPHIE .................................................................................................... 67
VILISTE DES FIGURES
Figure 1 : Microscopie électronique à transmission de S. suis sérotype 2. ............................ 2
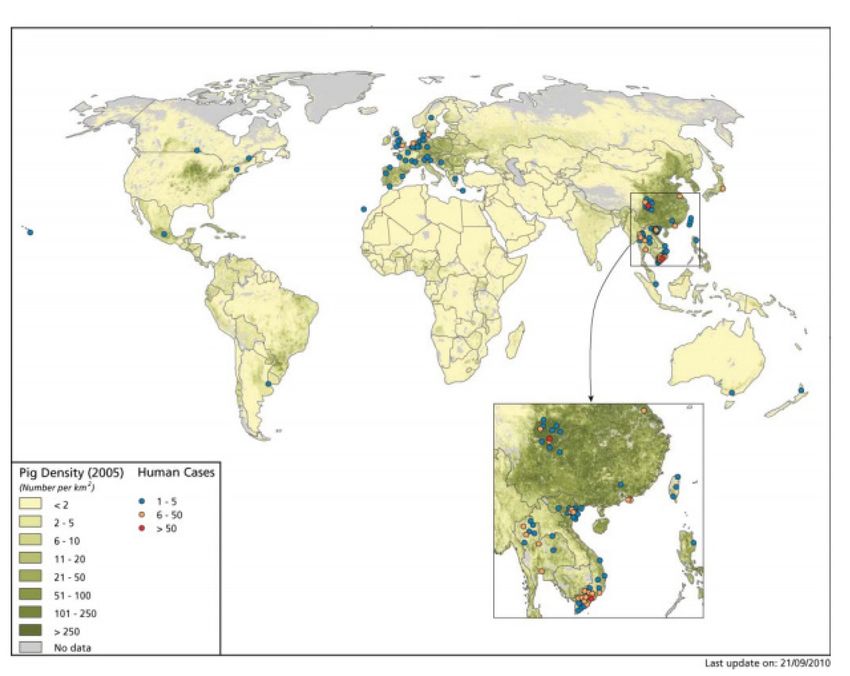
Figure 2 : Carte du monde représentant des cas d’infections à S. suis avec comme fond la
densité de la population porcine. ............................................................................................ 6
Figure 3 : Classification universelle des bactériocines proposée par Heng et Tagg............. 11
Figure 4 : Organisation génique de lantibiotiques linéaires et globulaires ........................... 13
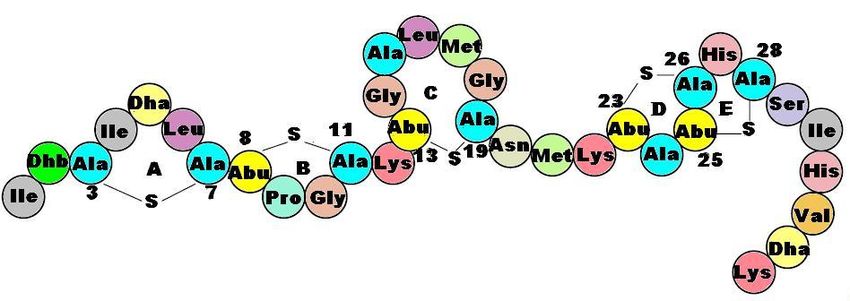
Figure 5 : Structure primaire de la nisine. ............................................................................ 16
Figure 6 : Cinétique de la viabilité de S. suis MGGUS2 en présence de nisine. .................. 35
Figure 7 : Microscopie électronique à transmission de S. suis MGGUS2 traité à la nisine. 37
Figure 8 : Zones d’inhibition produites par les souches S. suis 90-1330 et MGGUS13, deux
ST28, contre des isolats de S. suis provenant de porcs malades et appartenant soit au ST1
ou au ST25. ........................................................................................................................... 40
Figure 9 : Analyse de la fraction purifiée de la bactériocine produite par S. suis 90-1330 sur
gel Tris-Tricine par SDS-PAGE. .......................................................................................... 43
Figure 11 : Organisation génétique du groupe de gènes putatifs de la suicine 90-1330. ..... 49
Figure 12 : Comparaison de la séquence en acides aminés de la suicine 90-1330 avec
d’autres lantibiotiques ........................................................................................................... 50
Figure 13 : Structure primaire déduite de la suicine 90-1330............................................... 51
Figure 14 : Détection du gène de structure sslA de la suicine 90-1330 chez différents isolats
de S. suis. .............................................................................................................................. 52
Figure 15 : Détection du gène sly (suilysine), epf (extracellular protein factor) et gdh
(glutamate déshydrogénase) chez S. suis 90-1330 et P1/7 ................................................... 53
VIIVIII
LISTE DES TABLEAUX
Tableau 1 : Souches de S. suis sérotype 2 utilisées dans cette étude. ................................... 22
Tableau 2 : Zones d’inhibition produites par la souche productrice de nisine L. lactis ATCC
11454 contre des souches de S. suis appartenant à différents «sequence types». ................. 33
Tableau 3 : Concentrations minimales inhibitrices (CMIs) et concentrations minimales
bactéricides (CMBs) de la nisine contre des souches de S. suis appartenant à différents
«sequence types». .................................................................................................................. 34
Tableau 4 : Concentrations minimales inhibitrices (CMIs) de différents antibiotiques pour
les souches de S. suis MGGUS2 (ST1) et 1102864 (ST25). ................................................ 38
Tableau 5 : Effet de la combinaison de la nisine A avec différents antibiotiques
conventionnels vis-à-vis S. suis MGGUS2 et 1102864. ....................................................... 39
Tableau 6 : Effet de différentes sources et concentrations de carbone sur les zones
d’inhibition produites par la souche non-virulente S. suis 90-1330 contre les souches
indicatrices virulentes S. suis 24 et MGGUS2. ..................................................................... 41
Tableau 7 : Stabilité et spectre d’activité de la bactériocine purifiée de S. suis 90-1330. .... 45
Tableau 8 : Pourcentages d’identité des séquences d'acides aminés déduites entre le groupe
de gènes de la suicine 90-1330 de S. suis et le groupe de gènes de la nisine U de S. uberis.
.............................................................................................................................................. 49
Tableau 9 : Concentrations minimales inhibitrices (CMIs) et concentrations minimales
bactéricides (CMBs) de la pénicilline et de l’amoxicilline contre la souche S. suis 90-1330.
.............................................................................................................................................. 53
IXX
LISTE DES ABRÉVIATIONS
%: Pourcentage
°C : Degré Celsius
µg : Microgramme
µL : Microlitre
µM : Micromolaire
A. pleuropneumoniae : Actinobacillus pleuropneumoniae
A. suis : Actinobacillus suis
ADN : Acide désoxyribonucléique
ARNt: Acide ribonucléique de transfert
ATCC : American type culture collection
B. bronchiseptica : Bordetella bronchiseptica
BAGEL : Bacteriocin genome mining tool
BLAST : Basic local alignment search tool
C. botulinum : Clostridium botulinum
CMB : Concentration minimale bactéricide
CMI : Concentration minimale inhibitrice
Da : Dalton
Dha : 2,3-didehydroalanine
Dhb : 2,3-dehydrobutyrine
DO660 : Densité optique 660 nm
E. coli : Escherichia coli
EPF : Facteur extracellulaire protéique (gène : epf)
FIC : Fractional inhibitory concentration
XIFICI : Fractional inhibitory concentration index GDH : Glutamate déshydrogénase GRAS : Generally recognized as safe H. parasuis : Haemophilus parasuis HEPES : Acide 4-(2-hydroxyéthyl)-1-pipérazine éthane sulfonique HPLC : High performance liquid chromatography kb : Kilobase kDa : Kilo Dalton L: Litre L. lactis : Lactococcus lactis LAB : Lactic acid bacteria Lan : Lanthionine M: Molaire m/v : masse (g) / volume (mL) = gramme / millilitre Melan : 3-méthyllanthionine MES : Acide 2-(N-morpholino)-éthane-sulfonique mg : Milligramme min : Minute mL : Millilitre MLST : Multilocus sequence typing mm : Millimètre mM : Millimolaire MRP : Muraminidase-released protein MWCO : Molecular weight cut off N: Normal XII
NAD : Nicotinamide adénine dinucléotide
NCBI : National center for biotechnology information
nm : Nanomètre
ORF : Open reading frame
P. multocida : Pasteurella multocida
pb : Paire de bases
PBS : Phosphate-buffered saline
PCR : Polymerase chain reaction
PVDF : Polyfluorure de vinylidène
S. agalactiae : Streptococcus agalactiae
S. aureus : Staphylococcus aureus
S. hyicus : Staphylococcus hyicus
S. salivarius : Streptococcus salivarius
S. suis : Streptococcus suis
S. uberis : Streptococcus uberis
SDS-PAGE : Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
SLY : Suilysine (gène : sly)
ST : Sequence type
TFA : Acide trifluoroacétique
THA : Todd Hewitt agar
THB : Todd Hewitt broth
U: Unité
v/v : Volume/volume
XIIIXIV
REMERCIEMENTS
Je désire tout d’abord remercier mon directeur de recherche, Dr Daniel Grenier, pour sa
disponibilité, son soutien et tous ses précieux conseils apportés au cours de ma maîtrise. Je
tiens également à souligner l’apport des membres de mon comité d’encadrement, composé
de Dr Michel Frenette ainsi que de Dr Steve Labrie.
J’aimerais également exprimer ma reconnaissance envers tous les membres du laboratoire
qui, par leurs conseils et leur intérêt à l’égard de ma recherche, m’ont permis de progresser
au cours de ces deux années. Je dois un merci tout particulier à Katy Vaillancourt qui, par
son aide, sa présence et sa bonne humeur a grandement contribué au succès de ce projet. Je
tiens aussi à remercier le support financier fourni par le Ministère de l’Agriculture, des
Pêcheries et de l’Alimentation du Québec (MAPAQ) et le Centre de Recherche en
Infectiologie Porcine et Avicole du Fonds Québécois de la Recherche sur la Nature et les
Technologies (FQRNT).
Je désire également terminer en remerciant ma famille pour les encouragements et la
confiance qu’elle m’a témoignés tout au long de mon parcours universitaire ainsi que mon
copain Christian, pour m’avoir soutenue tout au long de ce projet. Sans ses
encouragements, sa patience et son amour, il m’aurait été difficile de me rendre si loin.
XVXVI
1. INTRODUCTION
1.1. Streptococcus suis
1.1.1. Généralités
Streptococcus suis (S. suis) est une bactérie gram-positive et anaérobie facultative qui se
présente sous forme de coque isolée ou en courte chaîne. De plus, la cellule est entourée
d’une capsule composée de polysaccharides riches en acide sialique (Figure 1).
Auparavant, les souches de S. suis étaient classées dans les groupes de Lancefield R, S, RS
et T, en se basant sur la composition de la capsule. Depuis, la classification a été
réorganisée et les souches sont maintenant distribuées en différents sérotypes à partir des
antigènes capsulaires. À ce jour, 35 sérotypes ont été identifiés (sérotype 1 à 34 et sérotype
1/2). Parmi ceux-ci, le sérotype 2 est le plus fréquemment isolé chez les porcs malades (1-
3). Les souches du sérotype 2 peuvent également être subdivisées en différents «sequence
types» (STs) à l’aide du «multilocus sequence typing» (MLST). Cette méthode permet de
mettre en évidence et de comparer les différences retrouvées dans les séquences d’ADN de
sept gènes constitutifs. Lorsqu’un gène contenant une séquence différente est identifié, un
numéro d’allèle lui est attribué. Ensuite, en tenant compte du profil allélique obtenu pour
tous les gènes de ménage, un numéro de ST est attribué à chaque souche. Celles
appartenant à un même ST sont considérées comme provenant d’un même clone (4). En
Amérique du Nord, parmi les souches du sérotype 2, les STs les plus souvent isolés chez le
porc sont le ST1, le ST25 et le ST28. Des recherches récentes utilisant un modèle murin ont
montré que les souches qui appartiennent au ST1 sont les plus virulentes. Il a aussi été
observé que les souches du ST25 ont une virulence intermédiaire et que celles du groupe
ST28 sont faiblement ou non-virulentes (5).
1Figure 1 : Microscopie électronique à transmission de S. suis sérotype 2. La flèche indique la capsule de polysaccharides entourant la bactérie. Les infections à S. suis sont reconnues pour causer diverses maladies chez le porc. En effet, S. suis peut provoquer des septicémies, de l’arthrite, des endocardites, des pneumonies et des méningites chez l’animal (6-9). Par conséquent, cet important pathogène engendre de graves problèmes économiques et sanitaires dans l’industrie porcine mondiale. Chez le porc, ce microorganisme est naturellement présent dans les voies respiratoires supérieures et plus particulièrement au niveau des amygdales. C’est d’ailleurs dans cette région que la bactérie est la plus couramment retrouvée chez le porc en santé (10-12). En effet, le porc est souvent porteur d’au moins une souche de S. suis sans toutefois manifester de symptômes cliniques de la maladie (13). S. suis est également isolé de plus en plus fréquemment chez de nombreux autres animaux tels que la vache, le cheval, le chien, le chat ainsi que certains oiseaux (1, 14). La capacité de la bactérie à coloniser ces différentes espèces animales permet d’accroître et de faciliter sa dispersion mondiale. C’est donc pour cette raison qu’un suivi épidémiologie de la maladie est nécessaire pour mieux comprendre la propagation de S. suis. 2
1.1.2. Facteurs de virulence
À ce jour, la pathogénicité de S. suis chez le porc n’est pas encore complètement élucidée.
Le processus d’infection est divisé en quatre grandes étapes : 1) l’adhérence et l’invasion
des surfaces épithéliales de l’hôte, 2) la dissémination et la survie dans la circulation
sanguine, 3) l’activation des processus inflammatoires et du choc toxique, et 4) l’infection
du système nerveux central (3). Au cours des dernières années, plusieurs facteurs de
virulence potentiels ont été identifiés chez S. suis et certains rôles ont pu leur être assignés.
Ces facteurs vont aider la bactérie à infecter, coloniser et provoquer la maladie chez l’hôte
malgré les mécanismes de défense de celui-ci. Plus spécifiquement, la capsule constituée
d’acide sialique et d’acide N-acétylneuraminique est reconnue pour jouer un rôle important
dans la virulence de S. suis (15). En effet, des mutants de S. suis sans capsule se sont avérés
être non-virulents dans des modèles murins et porcins (16). La présence de la capsule
permet à la bactérie d’échapper aux macrophages et aux neutrophiles, l’aidant ainsi à
survivre et se disséminer chez l’hôte (17, 18).
En plus de la capsule, une hémolysine produite par S. suis, nommée suilysine (SLY),
semble également jouer un rôle important dans la virulence. En effet, les hémolysines sont
connues pour être toxiques envers certaines cellules de l’hôte. Il a d’ailleurs été démontré
que la suilysine augmentait la lyse et l’invasion de l’épithélium par S. suis en plus
d’empêcher la phagocytose de la bactérie (18-20). Deux autres potentiels facteurs de
virulence ont aussi été proposés, soit la «muraminidase-released protein» (MRP) et le
«extracellular protein factor» (EPF). La présence de ces facteurs est généralement associée
avec la virulence des souches en provenance d’Europe et d’Asie (2, 21). Leurs rôles exacts
sont toutefois méconnus et leur présence ne semble pas être essentielle à l’infection (22). À
vrai dire, peu de facteurs de virulence montrent une association directe entre la présence du
facteur et la virulence de la souche. Cependant, un lien a pu être établi entre la capacité
d’expression de certains facteurs de virulence et différents STs (ST1, ST25 et ST28) des
souches de S. suis sérotype 2. En effet, selon les différents STs, il y a présence ou absence
des gènes mrp, epf et/ou sly qui codent respectivement pour le MRP, le EPF et la suilysine.
Les souches ST1, considérées comme virulentes, sont généralement SLY+MRP+EPF+,
3tandis que les souches modérément virulentes (ST25) sont SLY-MRP-EPF- et celles appartenant au groupe ST28 (faiblement ou non-virulentes) sont SLY-MRP+EPF- (5). Toutefois, il est important de noter que l’identification de certains facteurs de virulence ne permet pas de faire un lien direct entre la présence du facteur et l’apparition de la maladie. En effet, certains facteurs de virulence vont être présents autant chez des souches virulentes que non-virulentes. 1.1.3. Pathologies Chez le porc, les infections à S. suis sont plus courantes dans les élevages regroupant une plus grande population d’animaux. Comme mentionné précédemment, S. suis se retrouve naturellement dans les amygdales de l’animal. La transmission de la bactérie à l’intérieur d’un troupeau est habituellement liée à la présence de porteurs sains (8). En général, la bactérie affecte plus souvent les porcelets en colonisant, peu après la naissance, les voies respiratoires supérieures de ces derniers. Plus particulièrement, les porcelets seront plus à risque d’être colonisés par S. suis au moment de la naissance et lors de l’étape du sevrage (23-25). Dans une moindre mesure, S. suis peut aussi être transmis horizontalement par des bioaérosols (26). Quoique l’infection soit plus commune chez les jeunes animaux, il arrive toutefois que les bêtes en phase d’engraissement et celles plus âgées développent des symptômes de la maladie. Les premiers symptômes observables sont la fièvre, l’anorexie et la dépression (27). Dans la majorité des cas, les porcs vont présenter une méningite même s’il est possible que l’animal puisse développer d’autres maladies comme la septicémie ou l’endocardite (27-29). S. suis affecte non seulement l’industrie porcine, mais constitue également un problème pour la santé publique, de par sa capacité à infecter l’humain. C’est en 1968, au Danemark, que le premier cas humain d’infection à S. suis a été signalé (30). Quoique l’incidence de la maladie soit beaucoup plus faible chez l’humain, la bactérie peut tout de même causer la méningite, l’endocardite, la cellulite, l’arthrite et la pneumonie (31-33). La bactérie se 4
transmet lors de contacts rapprochés ou d’expositions courantes avec les porcs ou les
produits dérivés du porc. C’est l’une des raisons pour lesquelles S. suis est considéré
comme un agent de zoonose important dans les pays asiatiques puisqu’il y a une plus
grande proximité des humains avec les animaux. En effet, deux épidémies à S. suis ont été
déclarées en Chine, une première en 1998 et une deuxième plus importante en 2005. Cette
dernière a affecté au moins 200 personnes dont 38 qui en sont décédées (34). Des cas de
S. suis ont aussi été rapportés en Europe et dans les Amériques (35). Il est important de
noter que cette zoonose est retrouvée à l’échelle internationale (Figure 2) et qu’elle ne doit
donc pas être négligée.
5Figure 2 : Carte du monde représentant des cas d’infections à S. suis avec comme fond la densité de la population porcine. La figure est tirée du livre «Atlas of Human Infectious Diseases» (36). 6
1.2. Contrôle des infections à Streptococcus suis
Afin d’obtenir des produits alimentaires provenant du porc qui sont sécuritaires pour la
consommation humaine, il est important de maintenir la santé et le bien-être des animaux.
Pour ce faire, il y a actuellement deux moyens permettant de prévenir et/ou de traiter les
infections à S. suis, soit l’utilisation d’antibiotiques ou la vaccination.
1.2.1. Vaccination
La vaccination est une méthode de contrôle des pathogènes qui a fait ses preuves au cours
des dernières décennies (37). Les vaccins qui sont présentement disponibles pour
immuniser les porcs contre S. suis sont faits à partir de suspensions de bactéries inactivées
que l’on nomme bactérines. Cependant, ce type de vaccins pose un problème face aux
espèces bactériennes qui présentent une grande diversité antigénique (comme dans le cas S.
suis) puisque la vaccination contre un sérotype n’assure la protection que contre ce sérotype
spécifique (38, 39). Dans le cas de S. suis, en plus de l’existence de plusieurs sérotypes,
ceux-ci se subdivisent en différents STs, ce qui complique davantage la mise au point d’un
vaccin et entraîne une efficacité très limitée de ce dernier (40, 41). Présentement, aucun
antigène commun aux différents sérotypes n’a encore été identifié. C’est pourquoi jusqu’à
ce jour la vaccination demeure un procédé peu utilisé. De plus, les résultats obtenus lors
d’expériences précédentes sur la vaccination des porcs montrent une certaine inconsistance
(29, 42). Il existe toutefois des moyens pour tenter d’augmenter l’efficacité des vaccins, tels
que la préparation de vaccins contenant plusieurs sérotypes différents (43). Une alternative
également couramment employée consiste en l’utilisation de vaccin autogène. Pour ce faire,
la souche causant la maladie est d’abord isolée puis utilisée chez l’animal comme un vaccin
(39).
1.2.2. Administration d’antibiotiques
Les antibiotiques sont une des plus grandes découvertes du 20e siècle et représentent un
moyen couramment utilisé pour contrer les infections bactériennes. Dans l’industrie
7porcine, les antibiotiques sont utilisés pour prévenir et traiter les infections. De plus, ils sont également employés comme facteur de croissance quoique plusieurs pays aient maintenant interdit ce type d’utilisation (44). Les antibiotiques généralement employés pour traiter les infections à S. suis sont des bêta-lactames comme la pénicilline G de même que des céphalosporines (45, 46). Malheureusement, dû à une utilisation inadéquate et abondante des antibiotiques au cours des dernières années, l’industrie fait maintenant face à un problème de résistances bactériennes. 1.2.3. Résistance aux antibiotiques L’apparition de résistances à la tétracycline et aux macrolides chez S. suis a commencé à être observé dans les années 1980 au Danemark (47). Depuis, partout dans le monde, des résistances à différents antibiotiques sont mises en évidence. En effet, des résistances aux lincosamides, aux macrolides, aux tétracyclines et aux sulphonamides ont été observées mondialement et parfois chez plus de 85 % des souches de S. suis (12). Au Québec, il a été démontré que les bactéries étaient particulièrement résistantes à la clindamycine (91 %) et à l’érythromycine (85 %) (48). Actuellement, les mécanismes de résistance aux antibiotiques chez S. suis ne sont pas encore bien compris. Toutefois, quelques études ont identifié certains facteurs génétiques déterminants dans le développement de résistances (49). En effet, la résistance de S. suis à la tétracycline serait généralement causée par des gènes de protection (tetM et tetO) empêchant la liaison de l’antibiotique aux ribosomes (50, 51). Pour ce qui est des résistances aux macrolides, elles seraient principalement causées par : 1) la présence des gènes erm codant pour une méthylase qui permet la modification du ribosome au site de fixation des ARNt et 2) la présence des gènes mef servant à l’expression des pompes à efflux (49). La résistance aux bêta-lactames (pénicilline, céphalosporine) est moins propagée et diffère beaucoup selon la provenance des souches (45). La résistance serait due à la modification des protéines liant la pénicilline (12). 8
Plusieurs antibiotiques utilisés dans l’industrie porcine sont les mêmes que ceux employés
pour traiter les infections chez l’humain. De ce fait, les résistances acquises par les
bactéries pathogènes du porc courent le risque d’être transférées aux bactéries commensales
de l’animal. De plus, il y a une possibilité que ces résistances soient aussi transférées à
l’humain. Effectivement, une étude réalisée au Vietnam a montré que des souches de S. suis
isolées chez des patients atteints de méningite présentaient des résistances à la tétracycline
(83,2 %), à l’érythromycine (20,2 %) et au chloramphénicol (3,3 %) (52). Les personnes
œuvrant dans l’industrie porcine se retrouvent donc dans une impasse puisqu’ils n’ont pas
d’alternative saine et efficace pour combattre les infections à S. suis. C’est pourquoi, la
recherche de nouveaux composés antimicrobiens est présentement nécessaire afin
d’identifier des moyens alternatifs de combattre la maladie.
1.3. Bactériocines
1.3.1. Généralités
En 1925, A. Gratia a été le premier à découvrir les bactériocines lors d’une expérience
concernant deux souches d’Escherichia coli. Il a observé qu’une des souches d’E. coli
produisait une substance, une colicine, capable d’inhiber l’autre souche (53). Depuis cette
expérience, environ 200 bactériocines ont été répertoriées dans la littérature selon la base de
données Bactibase (http://bactibase.pfba-lab-tun.org/main.php) (54). Elles sont décrites
comme étant des peptides antimicrobiens d’origine bactérienne qui permettent l’inhibition
de la croissance d’espèces proches de la souche productrice et parfois d’espèces plus
éloignées (55). La production de bactériocines procurerait un avantage écologique à la
souche productrice vis-à-vis des autres microorganismes qui colonisent le même
environnement (56). Il existe une grande hétérogénéité parmi les bactériocines connues à ce
jour. En effet, les bactériocines peuvent être produites autant par des bactéries gram-
négatives que gram-positives. De plus, des différences ont pu être mises en évidence en
regard de la taille des peptides antimicrobiens, de leurs modes d’action, de la cible
microbienne ainsi que de leurs mécanismes immunitaires (57). Les bactériocines produites
9par les bactéries gram-positives, et plus particulièrement par les bactéries lactiques (LAB), ont été très étudiées au cours des dernières décennies. L’une des raisons de cet intérêt pour les LAB est liée au fait que ce type de bactéries est beaucoup utilisé dans l’industrie alimentaire et que les seules bactériocines utilisées commercialement appartiennent à cette catégorie (58). Une étude préliminaire a permis de démontrer la présence de bactériocines chez S. suis (59). De plus, selon le poids moléculaire de ces dernières, elles feraient potentiellement partie de la classe des lantibiotiques. 1.3.2. Classification Au cours des dernières années, plusieurs schémas de classification ont été proposés pour répertorier les bactériocines. Tout d’abord, en 1993, Klaenhammer a proposé une première classification pour les bactériocines produites par des LAB. Comme illustré à la Figure 3A, ces dernières sont séparées en quatre grandes classes : les lantibiotiques, les peptides non- modifiés, les protéines de haut poids moléculaire sensible à la chaleur et les bactériocines complexes (57). Par la suite, en 2005, Cotter a suggéré une mise à jour du schéma de Klaenhammer en tenant compte des dernières avancées dans le domaine. Son schéma présenté à la Figure 3B permet de répartir les bactériocines en deux principaux groupes, les lantibiotiques et les non-lantibiotiques, ainsi qu’en bactériolysines (60). La classification qui sera présentée dans ce travail est celle proposée par Heng et Tagg en 2006 (Figure 3C) (61). Cette classification est considérée comme étant universelle puisqu’elle permet le classement des bactériocines provenant autant des bactéries gram-négatives que gram- positives. De plus, elle prend en considération les deux méthodes de répartition proposées antérieurement. Cette classification divise les bactériocines en quatre grandes classes : les lantibiotiques, les peptides non-modifiés, les protéines de haut poids moléculaire et les peptides cycliques. 10
Figure 3 : Classification universelle des bactériocines proposée par Heng et Tagg (C) basée
sur la classification des LAB par Klaenhammer (A) et la classification de Cotter et al. (B).
Figure traduite à partir du schéma de classification de Heng et Tagg (61).
1.3.2.1. Classe I : lantibiotiques
Les lantibiotiques sont des petits peptides ayant une taille inférieure à 5 kDa. Ils sont
reconnus pour leur grande stabilité à la chaleur et aux variations de pH. Cependant, la
caractéristique principale qui les différentie des autres bactériocines est la présence de ponts
thioéthers et d’acides aminés modifiés (Dha, Dhb) ajoutés post-traductionnellement (62).
Les ponts sont formés à partir de résidus lanthionine (Lan) ou 3-méthyllanthionine (Melan)
11d’où la provenance du nom lantibiotique (63). Ce groupe de bactériocines se divise en trois catégories selon la structure des peptides et leurs modes d’action : les linéaires (type A), les globulaires (types B) et les multi-composantes (type C) (61, 64). Les lantibiotiques dits linéaires sont en général des peptides cationiques ayant une forme allongée. Cette catégorie comprend des bactériocines comme la nisine A, l’épidermine et la lacticine 481. Les lantibiotiques linéaires sont en plus subdivisés en deux sous-classes, A(I) et A(II), selon leur structure. Les bactériocines globulaires, pour leur part, comprennent des peptides de forme sphérique chargés négativement comme la mersacidine et la cinnamycine (65, 66). Les peptides multi-composants sont des bactériocines dont l’activité antibactérienne est le résultat de la combinaison d’au moins deux peptides. La lacticine 3147 (67) et la cytolysine (68) se retrouvent dans cette catégorie. 1.3.2.1.1. Biosynthèse et organisation génique La synthèse des lantibiotiques se produit par voie ribosomique et a recourt à l’action de produits de plusieurs gènes organisés en «cluster» (69). Ces derniers peuvent se situer dans le chromosome (70), sur des éléments transposables ou non (71), ou sur un gros plasmide (72-74). Le symbole lan a été choisi pour nommer les gènes de biosynthèse des lantibiotiques. Une appellation plus précise est toutefois utilisée pour chaque lantibiotique. Par exemple, la terminologie nis est utilisée pour les gènes impliqués dans la production de la nisine. Un lantibiotique est formé à partir d’un pré-peptide constitué d’un peptide signal et d’un peptide mature inactif (75). Les différents «clusters» de gènes impliqués dans la formation du peptide vont servir à la production, à l’immunité (lanFEG, lanI, lanH) et à la régulation (lanR, lanK, lanQ) du lantibiotique (Figure 4). Parmi les gènes participant à la production, on retrouve des transporteurs ABC (lanT) ainsi que des gènes servant aux modifications post-traductionnelles (lanB, lanC, lanM, lanD et lanJ) et au clivage du peptide signal (lanP parfois associé à lanT) (65). Le pré-peptide, codé par le gène de structure lanA, subit deux 12
étapes de modifications post-traductionnelles. D’abord, la sérine est déshydratée en Dha et
la thréonine en Dhb. Ensuite, des ponts thioéthers sont constitués à partir des cystéines et
des résidus Dha/Dhb (76). C’est ainsi que les molécules lanthionine et méthyllanthionine
sont formées. Le peptide signal est ensuite clivé et le peptide mature est transporté à
l’extérieur de la cellule par le transporteur ABC. C’est à cette étape que le peptide actif est
obtenu (77).
Figure 4 : Organisation génique de lantibiotiques linéaires et globulaires. A : Nisine A,
peptide linéaire type A(I) ; B : Nukacine ISK-1, peptide linéaire de type A(II) ;
C : Mersacidine, peptide globulaire. Les fonctions similaires sont colorées ou hachurées de
la même façon. La figure est tirée de l’article d’Asaduzzaman et al. (78).
1.3.2.1.2. Mode d’action
Le mode d’action des lantibiotiques varie en fonction de la sous-classe dont ils font partie.
Les lantibiotiques linéaires (type A) ciblent des composantes de la membrane cellulaire
comme le lipide II, qui est un précurseur de la synthèse du peptidoglycane. En effet, il a été
démontré que la nisine, un lantibiotique linéaire, lie le lipide II (79). Ce dernier est un
13élément essentiel pour la formation de la paroi cellulaire et il est conservé parmi les bactéries. La liaison du lantibiotique et du lipide II entraine la formation de pores dans la membrane cellulaire. De plus, la bactériocine bloque également la biosynthèse de la paroi cellulaire en empêchant l’étape de transglycosylsation et en séquestrant le lipide II de son site actif situé au septum (80, 81). L’ensemble de ces mécanismes va entraîner la mort cellulaire qui sera causée par la perte du contenu cytoplasmique (acides aminés, ions, etc.) (82, 83). Il a été proposé que les deux modes d’action distincts de la nisine seraient l’une des raisons pour lesquelles peu de résistances ont été développées chez les bactéries malgré une utilisation régulière depuis les années 1980 (76, 84). Les lantibiotiques globulaires (type B) vont plutôt interrompre les fonctions de certaines enzymes nécessaires à la synthèse du peptidoglycane (66). Par exemple, la mersacidine inhibe la biosynthèse de la paroi cellulaire en formant un complexe avec le lipide II. Contrairement à la nisine, elle ne forme pas de pores dans la membrane cellulaire mais interfère avec l’étape de transglycosylation qui permet la polymérisation des chaînes glucidiques dans la formation du peptidoglycane (85). Ceci a pour conséquence d’empêcher les précurseurs du peptidoglycane associés lipide II d’être ajouté au peptidoglycane en croissance, entrainant graduellement la lyse de la cellule. Cependant, les lantibiotiques globulaires ne se lient pas tous au lipide II. En effet, la cinnamycine et la duramycine vont plutôt inhiber la phospholipase A2 et vont donc interférer avec la biosynthèse de la prostaglandine et des leucotriènes (69). Il faut toutefois savoir que dans plusieurs cas, la cible moléculaire de la plupart des lantibiotiques n’est pas encore identifiée. Les lantibiotiques multi-composants nécessitent l’action de deux peptides ou plus pour qu’il y ait une activité antibactérienne complète. La combinaison des deux peptides permet d’augmenter le spectre d’action des ces lantibiotiques (86). Présentement, seulement quatre lantibiotiques multi-composants ont été identifiés dans la littérature, soit la lacticine 3147, la plantaricine W, la staphylococcine C55 et la cytolysine (87). Le mode de fonctionnement 14
de la lacticine est celui qui a été le plus étudié. L’action inhibitrice de cette bactériocine
implique la participation de la lacticine 3147 A1 et de la lacticine 3147 A2. L’effet combiné
des deux peptides entraine la formation de pores dans la membrane cellulaire. Il y a alors
dépolarisation de celle-ci due à la fuite du contenu cellulaire qui comprend des ions K+ et
des ions phosphate. Il a été proposé que le peptide A1 agirait en se liant au lipide II,
permettant ainsi au peptide A2 de se lier et de former un pore dans la membrane
cytoplasmique de la cellule (86-88).
1.3.2.1.3. Nisine A
La nisine est un lantibiotique connu depuis déjà plusieurs années. En effet, son action
inhibitrice a été observée pour la première fois dans les ferments laitiers en 1928 (89).
Actuellement, cinq variations naturelles de la nisine (A, Z, Q, F, U) ont été décrites (90). À
l’exception de la nisine U produite par Streptococcus uberis, tous les autres types sont
produits par Lactococcus lactis. Parmi ces variations, la nisine A est celle qui a été la plus
étudiée et qui sera traitée dans ce travail. Cette dernière est un lantibiotique faisant partie
des lantibiotiques linéaires (type A(I)). Elle est composée de 34 acides aminés et de 5 ponts
thioéthers (Figure 5) (91). La nisine A montre une activité inhibitrice efficace contre
plusieurs bactéries gram-positives et sur certaine bactéries gram-négatives. En effet, elle est
capable d’inhiber plusieurs pathogènes alimentaires tel que Listeria monocytogenes, E. coli,
Staphylococcus aureus, Clostridium botulinum (92-94), des salmonelles (95) ainsi que
certains streptocoques (96-98). De plus, il a été montré que la nisine a une activité
inhibitrice permettant d’empêcher la formation de spores bactériennes (99, 100). Comme
mentionné précédemment, puisqu’elle fait partie des lantibiotiques linéaires, son activité
inhibitrice s’effectue par la liaison au lipide II, bloquant ainsi la biosynthèse de la paroi
cellulaire et entraînant la formation de pores dans la membrane cellulaire (81).
Depuis qu’elle a été approuvée par l’Union européenne en 1983 (101) et par la FDA en
1988 (102), la nisine est régulièrement utilisée comme agent de conservation dans
15l’industrie alimentaire (Nisaplin®) (93). Plus particulièrement, elle est beaucoup employée comme additif dans les aliments en conserve et sous vide pour empêcher la croissance de C. botulinum. La nisine sert également dans l’industrie laitière pour prolonger la durée de vie des produits. Même si son utilisation est largement répandue dans le domaine alimentaire, la nisine n’est que peu exploitée dans d’autres domaines tels que la santé, l’environnement et les biotechnologies. Plusieurs études ont toutefois amené des preuves pour une potentielle utilisation dans les nanotechnologies (103), pour les implants médicaux (104), en médecine dentaire (98, 105) et en médecine vétérinaire (97). En effet, une étude a permis de démontrer que la nisine est efficace contre le pathogène S. aureus, responsable de la mastite chez la vache. Il existe présentement un médicament (WIPE OUT® Immucell) fait à base de nisine qui permet de prévenir et de traiter les mammites chez la vache. Figure 5 : Structure primaire de la nisine. Les acides aminés modifiés sont identifiés comme Dha et Dhb. La structure comprend 34 acides aminés dont cinq ponts thioéthers (Ala-S-Ala = lanthionine, Abu-S-Ala = β- méthyllanthionine) identifiés avec les lettres A, B, C, D et E. La figure est tirée de la thèse de Olli Koponen (106). 16
1.3.2.2. Classe II
La classe II (peptides non-lantibiotiques) est constituée de peptides d’une taille inférieure à
10 kDa qui ne contiennent pas de peptides modifiés. Cette classe se subdivise en trois sous-
classes (IIa, IIb, IIc). La classe IIa, nommée pseudo-pédiocine, comprend des peptides
ayant une séquence consensus YGNGV en N-terminale en plus de posséder une activité
spécifique contre Listeria monocytogenes (55, 60, 107). La classe IIb contient des peptides
hétérogènes qui incluent des peptides linéaires et différents de la pédiocine. Enfin, la classe
IIc, les peptides multi-composants, renferme des bactériocines qui ont besoin d’au moins
deux peptides pour exercer une activité antibactérienne.
1.3.2.3. Classe III
La classe III de la classification de Klaenhammer, des protéines de grandes tailles (>10
kDa) sensibles à la chaleur (57), est conservée dans la classification de Heng et Tagg. Par
contre, cette classe est divisée en deux sous-classes de peptides : bactériolytiques et non-
lytiques. Les peptides bactériolytiques agissent au niveau du peptidoglycane, résultant en la
lyse de la cellule. Parmi ces bactériocines, la lysostaphine est le prototype à la base de cette
sous-classe. Les peptides non-lytiques, comme leur nom l’indique, ne lysent pas les
cellules. Le mode d’action précis de ces peptides n’est pas encore bien connu. Il a été
proposé que le mode d’action repose sur la dissipation de la force proton motrice entraînant
la mort de la cellule (108), tel que décrit précédemment pour d’autres bactériocines (109).
1.3.2.4. Classe IV
Les bactériocines de cette classe sont des peptides cycliques synthétisés par voie
ribosomale. Cette appellation est liée au fait que ces peptides sont modifiés post-
traductionnellement, ce qui entraîne la formation d’un lien covalent entre les extrémités N-
et C-terminales (108). Présentement, peu de bactériocines ont été identifiées dans cette
classe. L’une des plus étudiées est l’entérocine AS-48 produite par Enterococcus faecalis.
17Cette dernière est une bactériocine qui a une stabilité plutôt élevée due à sa forme cyclique. C’est également ce qui lui permettrait de conserver sa forme active (110). 1.3.3. Applications des bactériocines Présentement, seulement deux bactériocines, la nisine et la pédiocine, sont approuvées pour une utilisation commerciale dans l’industrie alimentaire. De plus, ces bactériocines sont considérées comme étant sans danger pour une utilisation en tant qu’agent de conservation et les souches productrices portent le statut «generally recognized as safe» (GRAS). La nisine est efficace contre plusieurs bactéries gram-positives ainsi que certaines gram- négatives. C’est pourquoi, en plus d’être employées dans l’industrie alimentaire, elle est aussi utilisée dans d’autres domaines. Par exemple, elle sert pour le traitement de la mastite chez la vache. En effet, le médicament WIPE OUT® Immucell fait à base de nisine permet de contrôler les deux principaux agents causant la maladie, S. aureus et Streptococcus agalactiae. D’autre part, une grande variété de bactériocines ayant démontré un potentiel pour une utilisation dans les domaines médicaux, alimentaires ou environnementaux ont été identifiées. Par exemple, certaines études ont amené des preuves pour une application potentielle dans le traitement des maladies de la peau (111), le contrôle de la carie dentaire (112) et le contrôle des infections de la gorge aux streptocoques (113). Il a aussi été proposé que les bactéries productrices de bactériocines pourraient avoir un rôle comme probiotiques (114). L’utilisation de bactériocines offre donc de grandes possibilités et demeure un sujet à explorer. 18
Vous pouvez aussi lire

















































