Blé, d'orge, de seigle et d'avoine
←
→
Transcription du contenu de la page
Si votre navigateur ne rend pas la page correctement, lisez s'il vous plaît le contenu de la page ci-dessous
Évaluation du pouvoir discriminatif d’anticorps créés
à partir de peptides synthétiques issus de gluten de
blé, d’orge, de seigle et d’avoine
Mémoire
David Poirier
Maîtrise en sciences des aliments - avec mémoire
Maître ès sciences (M. Sc.)
Québec, Canada
© David Poirier, 2021Évaluation du pouvoir discriminatif d’anticorps
créés à partir de peptides synthétiques issus de
gluten de blé, d’orge, de seigle et d’avoine
Mémoire
David Poirier
Maîtrise en sciences des aliments
Maître ès sciences (M.Sc.)
Sous la direction de :
Samuel Godefroy, directeur de rechercheRésumé
Les symptômes de la maladie cœliaque sont déclenchés par l'ingestion de céréales contenant
du gluten comme le blé, l'orge, le seigle et, dans de rares cas, l'avoine. Afin d’éviter ceux-ci,
les personnes touchées doivent adopter un régime sans gluten. Pour être déclaré sans-gluten,
un aliment ne doit pas contenir plus de 20 mg/kg de gluten. Ainsi, ces denrées doivent être
analysées afin de garantir leur innocuité. Actuellement, des tests immunochimiques basés sur
la reconnaissance de séquences protéiques de gluten sont utilisés. Cependant, des problèmes
de sur- et de sous-estimation de la teneur exacte en gluten surviennent en fonction de la source
de gluten analysée, c’est-à-dire selon la céréale. En cas de surestimation, cela réduit l’offre
alimentaire des personnes cœliaques, mais en cas de sous-estimation, de graves conséquences
affectent cette population. De plus, la réglementation canadienne exige une déclaration de la
source du gluten sur l'étiquetage des produits préemballés, ce qui ne peut être réalisé
adéquatement avec les moyens actuels.
Ce travail porte sur le développement de nouveaux anticorps visant à discriminer les sources
de gluten en utilisant des peptides synthétiques comme stratégie d'immunisation. Pour ce
faire, 14 peptides synthétiques sélectionnés de séquences protéiques de gluten ont chacun été
bioconjugués à une protéine porteuse et l'immunisation de lapins a été réalisée. Les anticorps
polyclonaux (pAbs) résultants distinguent avec succès le gluten de blé, d'orge et d'avoine.
Les pAbs dirigés contre le seigle réagissent également avec le gluten de blé et de seigle. Les
résultats obtenus démontrent que la discrimination des sources de gluten peut répondre à la
législation canadienne, mais aussi complémenter les tests immunochimiques actuels en
nivelant les problèmes de sur- et sous-estimation de la teneur en gluten et améliorer
davantage la sécurité des aliments destinés aux patients cœliaques.
iiAbstract
Symptoms of celiac disease are triggered by ingesting grains that contain gluten like wheat,
barley, rye, and, in rare cases, oats. In order to avoid symptoms, those affected should adopt
a gluten-free diet. To be declared gluten-free, a food must not contain more than 20 mg / kg
of gluten. Thus, these foods must be analyzed in order to guarantee their safety. Currently,
immunoassays based on the recognition of gluten protein sequences are used. However,
problems of overestimating and underestimating of the exact gluten content arise depending
on the type of gluten being analyzed (wheat, barley, rye or oat). If gluten is overestimated,
this reduces the food supply of celiac patients, but if underestimated, serious consequences
then affect this population. In addition, Canadian regulations require a declaration of the
source of gluten on the labelling of prepackaged products, which cannot be done adequately
with current means.
This work focuses on the development of new antibodies aimed at discriminating between
sources of gluten using synthetic peptides as immunization strategy. To proceed, 14 synthetic
peptides selected from gluten protein sequences were each bioconjugated to a carrier protein
and rabbit immunization was performed. The resulting polyclonal antibodies (pAbs)
successfully distinguish gluten from wheat, barley and oats. Rye pAbs react evenly with
wheat and rye gluten. The results obtained demonstrate that the discrimination of gluten
sources can meet Canadian legislation, but also complement current immunoassays by
levelling the problems of over and underestimation of gluten content and further improve the
safety of food intended for celiac patients.
iiiTable des matières
Résumé ................................................................................................................................... ii
Abstract .................................................................................................................................. iii
Table des matières ................................................................................................................. iv
Liste des figures .................................................................................................................... vii
Liste des tableaux ................................................................................................................ viii
Liste des abréviations, sigles, acronymes .............................................................................. ix
Remerciements ..................................................................................................................... xii
Avant-propos ....................................................................................................................... xiv
Introduction ............................................................................................................................ 1
1. Revue de littérature ............................................................................................................. 3
1.1-Les céréales nuisibles pour la population cœliaque...................................................... 4
1.2-Composition protéique des grains céréaliers contenant du gluten ............................... 5
1.3-Méthodes d’analyse du gluten ...................................................................................... 7
1.3.1-Chromatographie liquide à haute performance (HPLC) ........................................ 7
1.3.2-Spectrométrie de masse.......................................................................................... 7
1.3.3-Méthodes basées sur la génomique ........................................................................ 8
1.3.4-Méthodes immunochimiques ................................................................................. 8
1.3.5-Législation entourant l’analyse du gluten ............................................................ 10
1.4-Facteurs influençant la réponse des tests ELISA de gluten ........................................ 11
1.4.1-Extraction du gluten de la matrice alimentaire .................................................... 11
1.4.2-Standard de référence utilisé pour l’étalonnage ................................................... 12
1.4.3-Niveau de transformation de la matrice alimentaire ............................................ 12
1.4.4-Facteur de conversion pour l’obtention de la teneur totale de gluten .................. 13
1.4.5-L’anticorps utilisé dans le test ELISA ................................................................. 14
1.5-Anticorps actuellement utilisés dans les tests ELISA de gluten ................................. 14
1.5.1-Skerritt : anticorps contre ω-gliadine ................................................................... 14
1.5.2-G12 et α1 : anticorps contre 33-mer .................................................................... 15
1.5.3-α20 : anticorps contre l’α-gliadine ....................................................................... 15
1.5.4-R5 : anticorps contre le pentapeptide ................................................................... 16
iv1.5.5-Anticorps polyclonaux ......................................................................................... 16
1.6-Méthode de production des anticorps dirigés contre le gluten ................................... 17
1.6.1-Induction de la réponse immunitaire humorale ................................................... 17
1.7-Utilisation de peptides synthétiques comme immunogène ........................................ 18
1.7.1-Expositions antigéniques des protéines natives par rapport aux peptides
synthétiques ................................................................................................................... 18
1.7.2-Bioconjugaison des peptides synthétiques à une protéine porteuse..................... 19
2. But, hypothèse et objectifs................................................................................................ 21
2.1 But et hypothèse.......................................................................................................... 22
2.2 Objectifs ...................................................................................................................... 22
3. Évaluation du pouvoir discriminatif d’anticorps créés à partir de peptides synthétiques
issus de gluten de blé, d’orge, de seigle et d’avoine............................................................. 23
3.1 Résumé........................................................................................................................ 24
3.2 Abstract ....................................................................................................................... 25
3.3 Introduction ................................................................................................................. 26
3.4 Materials and methods ................................................................................................ 27
3.4.1 Materials ............................................................................................................... 27
3.4.2 Research and selection of sequences .................................................................... 27
3.4.3 Synthetization ....................................................................................................... 29
3.4.4 Bioconjugation of immunogen ............................................................................. 29
3.4.5 BSA bioconjugate ................................................................................................ 29
3.4.6 Immunization of rabbits ....................................................................................... 30
3.4.7 Preparation of gluten protein fractions ................................................................. 30
3.4.8 Titer determination by indirect ELISA ................................................................ 30
3.4.9 Sensitivity and specificity tests ............................................................................ 31
3.5 Results and discussion ................................................................................................ 32
3.5.1 Selection of the sequences.................................................................................... 32
3.5.2 Titer determination ............................................................................................... 34
3.5.3 Sensitivity and specificity tests ............................................................................ 36
3.6 Conclusions ................................................................................................................. 40
3.7 Acknowledgments ...................................................................................................... 41
vConclusion ............................................................................................................................ 42
Bibliographie ........................................................................................................................ 46
Annexe A – Séquences consensus de gluten ........................................................................ 57
viListe des figures
Figure 1: Principe de l'ELISA sandwich (A) et de l'ELISA compétitif (B) ......................... 10
Figure 2: Sites antigéniques (illustrés en rose) du virus de l'influenza HA1 lorsque
l'immunisation est réalisée avec le virus intact (à gauche) et lorsque l'immunisation est
réalisée à l'aide de plusieurs peptides synthétiques (à droite) .............................................. 18
Fig 3: Peptide selection flow chart ....................................................................................... 33
Fig 4: Titer determination by indirect ELISA against 1μg/ml of BSA-(Mal-PEG4-NHS)-
peptide (1-14).. ..................................................................................................................... 35
viiListe des tableaux
Tableau 1: Classification et proportion des protéines de stockage des Pooideae .................. 6
Tableau 2 : Séquence du 33-mer de l'α-2-gliadine ............................................................... 15
Table 3: Prolamins and glutelins from gluten-containing grains ......................................... 28
Table 4: Sensitivity tests by indirect ELISA. ....................................................................... 37
Table 5: Cross-reaction test by indirect ELISA.................................................................... 39
Table S1: Consensus sequences obtained from multiple alignments sequences (MSA) and
compilation ........................................................................................................................... 57
viiiListe des abréviations, sigles, acronymes
a.a. Acide aminé / amino acid
ADN Acide désoxyribonucléique / Deoxyribonucleic acid
ARN Acide ribonucléique / Ribonucleic acid
BSA Albumine de sérum bovin / Bovine Serum Albumin
CCH Concholepas concholepas hemocyanine /
Concholepas concholepas hemocyanin
CCMAS Codex Committee of Methods of Analysis and Sampling
CD Maladie cœliaque / Celiac disease
CS Séquence consensus / consensus sequence
cv. Cultivar
DTT Dithiothreitol
ELISA Essai d’immunoabsorption enzymatique /
enzyme-linked immunosorbent assay
FPLC Chromatographie liquide protéique rapide /
Fast Protein Liquid Chromatography
GP-HPLC Chromatographie en phase liquide à haute performance par
perméation de gel / Gel Permeation High Performance Liquid
Chromatography
GPT Type de protéine de gluten / Gluten Protein Type
GRAVY Moyenne d’hydropathie / Grand average of hydropathy
HMW-GS Sous-unité de gluten de haut poids moléculaire /
High Molecular Weight – Gluten Subunit
IRMM Institute for Reference Materials and Measurements
LC Chromatographie liquide / Liquid Chromatography
LMW-GS Sous-unité de gluten de bas poids moléculaire /
Low Molecular Weight – Gluten Subunit
LOQ Limite de quantification / Limit Of Quantification
mAbs Anticorps monoclonal / monoclonal antibody
Mal-PEG4-NHS Maleimido-Tetra(Ethylene Glycol)-Acetic Acid NHS
ixMALDI-TOF Ionisation laser assistée par une matrice–Analyseur à temps de vol /
Matric Assisted Laser Desorption Ionisation – Time-of-flight
MHC Major Histocompatibility Complex /
Complexe majeur d’histocompatibilité
MMW-GS Sous-unité de gluten de poids moléculaire moyen /
Medium Molecular Weight – Gluten Subunit
MS Spectrométrie de masse / Mass Spectrometry
MSA Alignment de séquences multiple / Multiple Sequence Alignment
NCBI National Center of Biotechnology Information
NMR Résonance magnétique nucléaire / Nuclear Magnetic Resonance
O.D. Densité optique / Optic Density
pAbs Anticorps polyclonaux / polyclonal antibodies
PARERA Plateforme d’Analyse des Risques et d’Excellence en
Réglementation des Aliments
PBS Tampon phosphate salin / Phosphate Buffered Saline
PCR Réaction de polymérisation en chaîne / Polymerase Chain Reaction
ppm partie par million / parts-per-million
RP-HPLC Chromatographie liquide à haute performance en phase inverse /
Reversed Phase High Performance Liquid Chromatography
sulfo-SMCC sulfosuccinimidyl 4-[N-maleimidomethyl]cyclohexane-1-
carboxylate
TCEP tris(2-carboxyethyl)phosphine
UV Ultraviolet
WGPAT Working Group on Prolamin Analysis and Toxicity
x« Je cultive mon jardin. Et dans ma vie
professionnelle comme dans mon carré de jardin,
j'ai bien l'intention d'exclure les navets ! »
Louis de Funès
xiRemerciements
Je ne pourrais commencer ce mémoire sans remercier toutes les personnes qui m’ont soutenu
autant professionnellement que personnellement dans ce projet. Premièrement, merci à
Samuel Godefroy pour son accueil dans son équipe de recherche ainsi que pour la latitude et
la confiance accordée dans toutes les phases de cette maîtrise.
Merci à Jérémie Théolier pour ses bons conseils et ces heures de réflexions communes dans
une branche de la biochimie qui nous était alors obscure et pleine de défis avant ce projet.
Merci à Virginie Barrere de s’être jointe à d’innombrables reprises à nos « brainstorming »
animés. Merci également pour son aide précieuse en biologie moléculaire qui a été plus
qu’utile pour ce projet. Merci aux membres de l’équipe de la PARERA : Emilie Manny,
Attara Hell, Gabrielle Vatin, Eliane Picard-Deland, Sébastien La Vieille, Silvia Dominguez,
Gabor Molnar et Joseph Touma. Merci pour vos conseils et votre bonne humeur durant ces
deux années.
Je tiens à remercier du fond du cœur toute l’équipe du CER Groupe de m’avoir si bien
accueilli et de m’avoir permis d’utiliser leur laboratoire pendant ces deux mois ainsi que de
m’avoir partagé un bout de leur culture si riche qu’est la Belgique. Merci à Philippe Delahaut
et Natalie Gillard pour la latitude d’action inconditionnelle qu’ils m’ont offerte
gracieusement dans leurs installations. Un merci spécial à Riccardo Marega pour son aide
technique précieuse, son temps et ses bons conseils dans la mise en œuvre de la stratégie
d’immunisation. Merci à Anne-Catherine Huet pour son aide technique concernant les
immunoessais. Merci également à Melody Paulus, Amandine Lamote, Catherine Jacqmin,
Jean Henrottin et Maxime Gavage pour leur bon accueil et leur support lors du séjour.
Je tiens à remercier les nombreux membres du personnel de l’Université Laval qui m’ont
apporté de l’aide dans ce projet. Un merci spécial à Diane Gagnon pour son travail
exemplaire et sa patience à toute épreuve. Merci à Sophie Fortin pour son aide technique
pour l’utilisation du FPLC. Merci également à Mélanie Martineau et Pascal Lavoie pour leur
temps et l’accès au laboratoire de transformation alimentaire.
xiiMerci au partenaire industriel R-Biopharm inc. USA/Canada et à l’Université Laval pour le
financement de ce projet via une bourse MITACS Accelerate. Merci à Semican International
inc. et Avena Foods qui ont gentiment fourni les grains utilisés dans ce projet.
Je voudrais glisser un mot pour remercier ma famille qui m’a toujours encouragé et soutenu
dans mes décisions scolaires, professionnelles et personnelles et qui reste toujours tout près
lorsque j’en ai besoin. Merci d’avoir toujours cru en moi.
Finalement, je voudrais remercier ma meilleure amie et ma bien-aimée conjointe Florence
Martin. Merci pour ton écoute et ton support dans les moments de joie comme dans les
moments difficiles. Merci pour tout ce temps, cette confiance et cette bienveillance que tu
me portes. Je t’en serai pour toujours reconnaissant. Sans toi, je ne serais certainement pas
en train d’écrire ces mots aujourd’hui. Sans toi, ce grade de maîtrise serait encore loin.
xiiiAvant-propos
Les travaux présentés dans ce mémoire s’intéressent à l’évaluation du pouvoir discriminatif
de nouveaux anticorps développés à partir de séquences peptidiques synthétiques issues de
gluten de blé, d’orge, de seigle et d’avoine afin d’en discriminer la source. Ils ont été financés
conjointement par l’entremise d’une bourse MITACS Acccelerate par l’Université Laval et
R-Biopharm inc. USA/Canada ainsi que par le CER Groupe.
Ce mémoire comprend quatre chapitres, le premier étant une revue de littérature présentant
l’état des connaissances concernant le gluten et les méthodes de déterminations associées. Ce
chapitre présente aussi l’intérêt et les motivations derrière la mise en œuvre du présent projet
de maîtrise. Le deuxième chapitre présente le but, l’hypothèse de recherche ainsi que les
objectifs du projet menant à la confirmation ou l’infirmation de l’hypothèse.
Le troisième chapitre de ce mémoire est présenté par insertion d’un article scientifique rédigé
en anglais et intitulé « Evaluation of the discriminatory potential of antibodies created from
synthetic peptides derived from wheat, barley, rye and oat gluten » qui sera soumis dans la
revue PLOS ONE. Il présente la méthodologie de sélection des peptides synthétiques qui ont
été utilisés et la création des immunogènes associés ainsi que les résultats de l’évaluation de
la spécificité des anticorps développés envers leur protéine de gluten respective. Les auteurs
de cet article sont David Poirier, Jérémie Théolier, Riccardo Marega, Philippe Delahaut et
Samuel Godefroy. David Poirier a réalisé la sélection et la bioconjugaison des antigènes, les
expérimentations en laboratoire, l’analyse des résultats, le traitement statistique des données
et la rédaction de l’article à titre de premier auteur. Jérémie Théolier a révisé et corrigé
l’article. Riccardo Marega a déterminé la stratégie de confection de l’immunogène utilisé et
supervisé l’immunisation des lapins et la récolte des sérums ainsi que révisé l’article. Philippe
Delahaut a révisé l’article. Samuel Godefroy a révisé et corrigé l’article ainsi que supervisé
les travaux de recherche du projet.
Finalement, le dernier chapitre présente les conclusions et les perspectives de recherche de
ce projet.
xivIntroduction
Il est estimé qu’un pour cent de la population mondiale souffre de la maladie cœliaque. Les
symptômes de cette dernière se manifestent par l’ingestion de céréales contenant du gluten
et provoquent des symptômes indésirables et de graves conséquences sur les microvillosités
de l’intestin de cette population [1]. Le gluten est un ensemble de protéines contenu dans le
blé, l’orge, le seigle et dans de rares cas l’avoine [2]. Pour éviter les effets néfastes provoqués
par la maladie, il n’existe à ce jour qu’un seul moyen : se soumettre à un régime sans gluten.
Le Codex Alimentarius indique dans la Norme pour les aliments diététiques ou de régime
destiné aux personnes souffrant d’une intolérance au gluten qu’un aliment ne dépassant pas
une teneur en gluten de 20 mg/kg peut être déclaré sans gluten [3]. Il est donc primordial
qu’une méthode de quantification du gluten fiable et sensible soit disponible afin de garantir
la sécurité des patients cœliaques. Depuis 2008, cette même norme stipule que la méthode à
privilégier pour la détermination du gluten est l’essai d’immunoabsorption enzymatique
(ELISA) R5 Méndez. Cette méthode est endossée par l’AOAC comme méthode officielle et
comme méthode de type I par le Codex Committee of Methods of Analysis and Sampling
(CCMAS) depuis 2006 [4]. Une méthode de type I détermine une valeur qui peut seulement
être obtenue par ladite méthode et sert, par définition, de seule méthode pouvant établir la
valeur acceptée pour l’élément qui est mesuré [5]. Plusieurs autres méthodes immuno-
enzymatiques de quantification du gluten existent sur le marché. Cependant, plusieurs
auteurs ont mis en lumière les problèmes d’harmonisation et de performance de ces tests
ELISA, incluant le R5. Plus spécifiquement pour le R5, des variabilités intrinsèques ont été
démontrées en fonction de l’origine du gluten analysé (orge, seigle, blé), ce qui mène à des
sur- et sous-estimations de la concentration exacte de gluten [6–9].
D’autres auteurs ont démontré que ces variabilités pouvaient être réduites si la calibration du
R5 était faite avec le même type de gluten que l’analyte [10–12]. Toutefois, cette calibration
nécessite de connaître l’origine du gluten, qui est fréquemment inconnue en situation
d’analyse courante comme dans le cas des contaminations croisées [7,8]. Dans un autre ordre
d’idée, la réglementation canadienne exige que la source de gluten soit déclarée selon
l’origine du grain sur les étiquettes des aliments préemballés [13]. Ainsi, pour répondre aux
1problèmes de variabilité du R5 et pour combler l’exigence réglementaire canadienne, il est
nécessaire de créer un nouvel outil de distinction de la source de gluten.
Par cette étude, il est donc visé de discriminer les différentes sources de gluten par la création
de nouveaux anticorps. Ces derniers devront être dirigés de façon à reconnaître des segments
uniques de blé, d’orge, de seigle et d’avoine. Pour ce faire, des peptides synthétiques issus
de gluten des quatre grains à l’étude seront utilisés afin de développer ces nouveaux
anticorps. L’avantage de cette stratégie, qui a déjà fait ses preuves par le passé, réside dans
la sélection en amont d’haptènes spécifiques [14–16]. Le premier objectif est donc
d’identifier les segments uniques à synthétiser en tant qu’haptène. Ensuite, le second objectif
est de créer l’immunigène par bioconjugaison des peptides synthétiques à une protéine
porteuse afin d’induire une réponse immunitaire humorale pour la production d’anticorps.
Finalement, le troisième objectif réside en l’évaluation du pouvoir discriminatif des anticorps
produits envers les différents types de gluten de blé, d’orge, de seigle et d’avoine.
21. Revue de littérature
31.1-Les céréales nuisibles pour la population cœliaque
La maladie cœliaque est une condition touchant environ 1,4 % de la population mondiale [1].
La première description claire de la maladie cœliaque a été donnée par Samuel Gee en 1888,
mais il a fallu attendre en 1950 pour commencer à élucider les facteurs causant les symptômes
de cette maladie [17]. C’est cette même année que la toxicité du blé pour les patients
cœliaques a été établie [18]. Quelque temps après, une série d’investigations a mené à la
conclusion que le seigle et l’orge étaient aussi toxiques, tandis que le riz, le sarrasin et le maïs
ne l’étaient pas [19–21]. La toxicité de l’avoine reste un sujet débattu dans la communauté
scientifique. Les plus récentes études montrent que les différences entre les cultivars d’avoine
sont la raison pour laquelle il n’a pas été clairement établi si l’avoine est sans danger pour
tous les patients cœliaques [22,23]. Les contradictions entre les études seraient dues à
l’utilisation de variétés d’avoine qui diffèrent sur l’immunoréactivité de leurs prolamines et
de leurs gènes prolaminiques. Des études plus récentes montrent que l’introduction d’avoine
pure dans l’alimentation d’enfants souffrant de la maladie cœliaque en conditions contrôlées
permettrait de traiter la maladie [24,25]. Cela dit, de plus amples recherches sont nécessaires
pour déterminer les effets de l’avoine chez les patients adultes ainsi que les variétés d’intérêt.
D’autres chercheurs tentent d’identifier et de sélectionner les cultivars d’avoine non toxiques
et ayant un bon potentiel agronomique [26]. En 2016, Santé Canada s’est positionné sur le
sujet après une revue de la littérature. Tout en reconnaissant que quelques personnes souffrant
de la maladie cœliaque semblent intolérantes à l'avoine, leur revue conclut que l'avoine non
contaminée par les céréales contenant du gluten (blé, seigle et orge) peut être ingérée sans
danger par la plupart des patients cœliaques. Elle conclut aussi qu’il n’y a aucune preuve que
la consommation d'avoine pure ne contenant pas plus de 20 ppm de gluten par les patients
atteints de la maladie cœliaque devrait être limitée à une quantité quotidienne spécifique.
Cependant, les personnes atteintes de la maladie cœliaque doivent observer une phase de
stabilisation avant d'introduire de l'avoine non contaminée ou pure dans leur régime sans
gluten [2]. Ainsi, le Canada adopte les recommandations du Codex Alimentarius selon
lesquelles un aliment ne dépassant pas une teneur en gluten de 20 ppm peut être déclaré sans
gluten [3]. La décision d’incorporer l’avoine non contaminée par du gluten de blé, d’orge ou
de seigle aux produits considérés sans gluten est une décision nationale adoptée au Canada
et aux États-Unis [2,27,28]. La législation européenne et australienne inclut l’avoine dans
4leur définition de grains contenant du gluten pouvant être nocifs pour la population cœliaque
[29,30]. Considérant les applications internationales qu'auront potentiellement les finalités
du projet, la détection de l'avoine est pertinente.
1.2-Composition protéique des grains céréaliers contenant du gluten
Les différents grains céréaliers contiennent plusieurs types de protéines qui sont classifiées
historiquement en fonction de leur solubilité et nommées les fractions d’Osborne [31]. Les
albumines sont solubles dans l’eau et les solutions salines diluées. Les globulines sont
insolubles dans l’eau, mais solubles dans les solutions salines diluées. Les prolamines sont
identifiées comme étant insolubles dans l’eau et dans les solutions salines, mais solubles dans
l’alcool aqueux 60-70 % (v/v). Les glutélines sont insolubles dans l’eau et les acides faibles.
Leur solubilisation est plus laborieuse, mais possible à plus haute température (50°C) dans
un solvant d’alcools aqueux (ex : 50 % propan-1-ol ou 60% éthanol) contenant un agent
réducteur (ex : 2-mercaptoéthanol ou DTT) et un agent désagrégeant (ex : urée ou
guanidium) [32]. Ce traitement permet de briser les ponts disulfures et d’obtenir des sous-
unités de glutélines qui sont alors solubles dans l’alcool comme les prolamines [31,33]. Les
albumines et les globulines (ALGL, ≈ 20-25 % du contenu en protéines du grain) sont
composées essentiellement d’enzymes et d’inhibiteurs enzymatiques, tandis que les
prolamines et les glutélines (≈ 75-80 % du contenu en protéines du grain) servent de protéines
de stockage. Ce sont ces protéines de stockage qui forment le gluten identifié chez le blé
comme étant les gliadines (prolamines) et les gluténines (glutélines), les sécalines chez le
seigle, les hordéines chez l’orge et les avénines chez l’avoine. En se basant sur les différences
de poids moléculaire, le gluten peut être divisé en sous-unités de haut poids moléculaire
(HMW-GS), de poids moléculaire moyen (MMW-GS) et de bas poids moléculaire (LMW-
GS) présentées au Tableau 1 [21,33].
5Tableau 1: Classification et proportion des protéines de stockage des Pooideaea
Groupes Blé Seigle Orge Avoine
HMW-GS (p) HMW-Sécalines (p) D-Hordéines (p)
HMW-GS -
11% 9% 5%
ω1,2-Gliadines (m) ω-Sécalines (m) C-Hordéines (m)
-
4% 18% 36%
MMW-GS
ω5-Gliadines (m)
- - -
3%
LMW-GS (p) γ-75k-Sécalines (p) B-Hordéines (p)
-
22% 48% 27%
γ-Gliadines (m) γ-40k-Sécalines (m) γ-Hordéines (m) Avénines
LMW-GS
27% 25% 32% 100%
α-Gliadines (m)
- - -
33%
a
Blé : cv. Monopol, Seigle : cv. Halo, Orge : cv. Golden Promise;
m = monomérique; p = polymérique.
Tableau adapté de [33]
Le Codex Alimentarius spécifie dans la Norme pour les aliments diététiques ou de régime
destiné aux personnes souffrant d’une intolérance au gluten (CODEX STAN 118-1979) que
l’anticorps utilisé dans une méthode d’analyse immunochimique «doit réagir avec les
fractions protéiques toxiques pour les personnes intolérantes au gluten et ne doit pas interagir
avec d’autres protéines de céréales» [3]. Cette contrainte implique donc que la détection du
gluten ne peut se faire de façon indirecte, comme par la détection des albumines et des
globulines des différents grains. Il est donc nécessaire de connaitre la toxicité des protéines
de stockage pour les patients cœliaques afin d’obtenir des anticorps pouvant être utilisés dans
des méthodes d’analyses immunochimiques. La littérature montre que les protéines de
stockage (prolamines et glutélines) du blé, de l’orge, du seigle et possiblement de l’avoine
pour les raisons indiquées précédemment peuvent induire des réactions chez les patients
cœliaques [34–39]. Les protéines de stockage sont donc toutes susceptibles de pouvoir être
utilisées pour la création d’anticorps à des fins d’analyses immunochimiques.
61.3-Méthodes d’analyse du gluten
1.3.1-Chromatographie liquide à haute performance (HPLC)
La chromatographie liquide à haute performance en phase inverse ou à perméation de gel
(RP- ou GP-HPLC) avec détection UV et l’électrophorèse sur gel sont les techniques les plus
utilisées pour la caractérisation des protéines des céréales [40,41]. Cependant, ces techniques
ont un pouvoir de séparation dans une seule dimension. Elles sont efficaces pour établir le
profil protéique de différentes farines, mais inefficaces pour la quantification de traces de
gluten dans les matrices alimentaires considérant leur basse sélectivité et sensibilité. Les
méthodes plus modernes combinent les méthodes de séparation à haute définition (le plus
souvent en deux dimensions) avec la spectroscopie de masse (MS) utilisant une source
d’ionisation laser assistée par une matrice (MALDI, Matrix-assisted laser
desorption/ionisation) et un analyseur à temps de vol (TOF, Time of flight) [42].
1.3.2-Spectrométrie de masse
Plusieurs études ont été menées pour employer la spectrométrie de masse comme méthode
de détection et de quantification des traces de gluten dans les aliments sans gluten. Les
méthodes par spectrométrie de masse ont d’abord été utilisées pour identifier les prolamines
toxiques impliquées dans la maladie cœliaque [43,44]. Une méthode comme le MALDI-
TOF-MS, même avec une extraction en deux étapes, s’est révélée inefficace pour de très
faibles concentrations de gluten avec une limite de détection de 100 ppm de gliadine. À cause
de l’insuffisance de sensibilité, la basse exactitude avec des protéines de haut poids
moléculaire et la baisse de résolution et d’intensité observables après le traitement thermique
des prolamines, la MALDI-TOF-MS est applicable seulement pour des mesures semi-
quantitatives [41,45].
Une autre approche utilisant la chromatographie liquide couplée avec une spectrométrie de
masse en tandem (LC-MS/MS) permet d’obtenir une meilleure résolution et donc une
meilleure précision [46]. Malgré cette amélioration, les études les plus récentes indiquent que
la quantification du gluten par LC-MS/MS reste une tâche très complexe considérant qu’il
existe actuellement un nombre limité de marqueurs peptidiques permettant d’identifier et de
7quantifier certaines séquences toxiques spécifiques de gluten [41]. De plus, les protéines du
gluten subissent fréquemment des modifications mineures (ex. délétion, substitution,
insertion) de certains acides aminés sur leur séquence qui n’affectent pas nécessairement le
potentiel immunogène des protéines, mais qui a un impact majeur sur les résultats d’analyse
obtenus par LC-MS/MS [47]. Il en est de même lors de l’analyse de produits transformés
dans lesquels le gluten subit certaines modifications [47]. Cette méthode montre certaines
lacunes au niveau de la sensibilité pour quantifier des traces de gluten, elle nécessite un
personnel hautement qualifié et l’utilisation d’équipements onéreux qui en font, pour le
moment, une méthode inappropriée pour les analyses de routine des industries alimentaires.
1.3.3-Méthodes basées sur la génomique
Les méthodes basées sur la génomique ne quantifient pas les protéines contrairement aux
autres méthodes, mais plutôt l’ADN ou l’ARN. Ces méthodes sont plus sensibles que les
méthodes basées sur les protéines. La réaction en chaîne par polymérase (PCR) permet
l’amplification de milliers de fragments de l’ADN cible. Ces méthodes présentent aussi
l’avantage de pouvoir distinguer aisément la source de gluten, soit le blé, l’orge, le seigle ou
l’avoine, ce qui en fait de très bonnes méthodes de détection du gluten [41,42]. Cependant,
Mujico et al. (2011) ont démontré que les procédés de transformation alimentaire utilisant
des traitements de chaleur ont pour conséquence de dénaturer les acides nucléiques, rendant
impossible l’établissement d’une corrélation entre la teneur en acides nucléiques et la teneur
en protéines de gluten [48]. Ainsi, l’utilisation des méthodes génomiques doit uniquement
être réservée à la détection de gluten tout en relativisant que leur résultat n’est pas forcément
indicateur de la présence de protéines de gluten, mais plutôt à la présence d’acide nucléique.
1.3.4-Méthodes immunochimiques
À ce jour, les méthodes immunochimiques sont les plus utilisées pour la détection et la
quantification du gluten [42]. L’essai d’immunoabsorption enzymatique (ELISA) est basé
sur la liaison spécifique d’anticorps et d’antigènes. Indépendamment du type de test ELISA,
l’un des anticorps a une liaison covalente avec une enzyme (ex. peroxydase de raifort ou
alcaline phosphatase) permettant de générer, à partir d’un substrat, des composés
8chimioluminescents, fluorescents ou colorimétriques pouvant être mesurés par
spectrophotométrie [42].
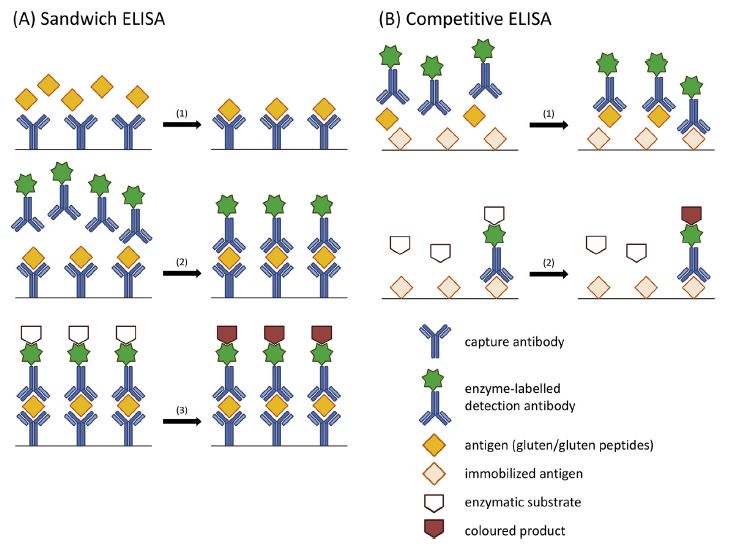
Le premier type d’ELISA pour la détermination du gluten est le sandwich dont le principe
schématique est présenté à la Figure 1. Le puits d’une microplaque est couvert avec une
quantité prédéterminée d’anticorps ayant leurs sites de liaison libres. L’échantillon contenant
potentiellement des antigènes de gluten en une concentration inconnue est ensuite introduit.
L’antigène, si présent, se lie aux anticorps libres fixés au fond de la microplaque. Le puits est
ensuite lavé avec une solution de lavage pour éliminer tous les autres constituants résiduels
non liés aux anticorps. Le conjugué, une solution d’anticorps liés à une enzyme, est ajouté
au puits et va se fixer à un épitope libre de l’antigène déjà en place. Le puits est ensuite lavé
pour retirer l’excès de conjugué. Un substrat réagissant avec l’enzyme du conjugué est
ensuite ajouté en une concentration connue. Ce substrat est alors transformé en composé
chimioluminescent, fluorescent ou colorimétrique proportionnellement à la quantité de
conjugué, qui elle est proportionnelle à la quantité d’antigènes fixés au fond du puits.
L’absorbance est ensuite lue à l’aide d’un spectrophotomètre et est comparée à celle de la
courbe des standards pour déterminer la concentration en antigènes [41]. Comme les ELISA
sandwich ont besoin de deux épitopes disponibles sur l’antigène, ils sont efficaces avec des
antigènes de plus grande taille. Cette condition rend ce test inefficace pour les produits
transformés contenant du gluten hydrolysé comme la bière ou les extraits de malt [33].
L’ELISA compétitif permet de contrer le problème de la nécessité de deux épitopes libres
sur le même antigène puisqu’il n’a besoin que d’un seul épitope [49]. Cette caractéristique le
rend adéquat pour détecter les protéines de gluten natives, mais aussi les plus petits peptides.
Comparativement à l’ELISA sandwich, les puits du test compétitif sont tapissés d’un
antigène (ex. peptides de gluten) au lieu d’immunoglobulines tel qu’illustré à la Figure 1.
Une quantité connue d’anticorps marqués par une enzyme et l’échantillon contenant
potentiellement du gluten sont ensuite ajoutés simultanément dans le puits. De cette façon,
les antigènes libres de l’échantillon et les antigènes fixés au fond du puits entrent en
compétition pour se lier à la quantité limitée d’anticorps libres. Le puits est ensuite lavé et
tous les anticorps s’étant liés aux antigènes libres de l’échantillon sont donc éjectés. Les
9anticorps restants sont liés aux antigènes fixés au puits et leur enzyme est donc disponible
pour réagir avec le substrat chromogène. Le composé chimioluminescent ou fluorescent ainsi
créé est ensuite dosé par spectrophotométrie en comparant l’absorbance avec une courbe
standard. La quantité de gluten présente dans l’échantillon est donc inversement
proportionnelle à l’absorbance mesurée [41].
Figure 1: Principe de l'ELISA sandwich (A) et de l'ELISA compétitif (B). Tiré de [41]
1.3.5-Législation entourant l’analyse du gluten
Le Codex Alimentarius fait état d’exigences générales entourant l’analyse du gluten dans sa
norme CODEX STAN 118-1979 [3]. La détermination quantitative du gluten devrait être
basée sur une méthode immunologique ou une méthode pouvant égaler la sensibilité et
spécificité de cette dernière. Comme indiqué précédemment, les anticorps utilisés doivent
réagir contre les fractions protéiques toxiques du gluten et ne doivent pas faire de réactions
croisées avec d’autres constituants. La limite de détection doit être plus petite ou égale à 10
mg de gluten/kg. De plus, depuis 2008, le Codex Alimentarius a endossé comme méthode de
type I l’immunoessai R5 Méndez pour la détermination du gluten.
101.4-Facteurs influençant la réponse des tests ELISA de gluten
1.4.1-Extraction du gluten de la matrice alimentaire
L’extraction du gluten de la matrice alimentaire est la première étape constituant la
détermination du gluten par test ELISA. Cette étape est critique pour obtenir une réponse
précise et exacte de la concentration en gluten. Historiquement, l’utilisation des prolamines
pour la quantification du gluten était privilégiée en raison de leur facilité d’extraction. En
effet, l’extraction des prolamines à l’aide d’alcools aqueux (ex. éthanol 60-70%) est plus
simple que l’extraction combinée des prolamines et des glutélines qui demande l’utilisation
d’agents dénaturants et désagrégeants [32,50,51]. Cependant, il a été démontré que les
glutélines ont un rôle à jouer sur le déclenchement des symptômes de la maladie cœliaque
[35,38,52,53]. Ainsi, il est primordial d’extraire les prolamines et les glutélines de l’analyte
afin d’obtenir une quantification du gluten total et non seulement des prolamines. Le cocktail
d’extraction utilisé pour l’ELISA R5, qui est l’ELISA le plus répandu, est une combinaison
de 2-mercaptoéthanol (250 mM) pour réduire les ponts disulfures liant les glutélines entre
elles et de chlorure de guanidium (2 M) servant d’agent désagrégeant, le tout dans un tampon
phosphate salin [32]. La suite de l’extraction se fait ensuite dans de l’éthanol 60% pour bien
solubiliser le gluten. Il a été confirmé que cette méthode présente un taux de récupération de
98 à 100% pour les gliadines [32]. Cette méthode d’extraction présente l’avantage d’extraire
les gliadines ayant subi un traitement thermique, ce qui est souvent le cas dans les produits
transformés contenant de la farine. Considérant la toxicité du 2-mercatoéthanol, une
alternative moins toxique présentant un taux d’efficacité similaire a été développée en
substituant le 2-mercatoéthanol par du TCEP, mais ce dernier étant plus onéreux que le 2-
mercaptoéthanol, celui-ci reste moins utilisé dans les trousses ELISA commerciales [54].
L’extraction du gluten est une variable encore instable. Bien qu’optimisée pour les gliadines,
il manque actuellement d’études sur l’efficacité des différents protocoles d’extraction sur les
glutélines et les prolamines de seigle, d’orge et d’avoine, et ce, en considérant aussi le niveau
de transformation de la matrice alimentaire comme les traitements thermiques ou
d’hydrolyse. [55].
111.4.2-Standard de référence utilisé pour l’étalonnage
Le standard de référence sert d’étalon pour l’élaboration de la courbe de calibration d’une
trousse ELISA. Le Working Group on Prolamin Analysis and Toxicity (WGPAT) ont
développé la «PWG-gliadin», un matériel de référence basé sur 28 cultivars de blé européens
représentant un ratio standard d’épitopes toxiques de blé pouvant servir de standard de
calibration dans les trousses ELISA [56]. En 2005, la PWG-gliadin a été soumise à l’Institute
for Reference Materials and Measurements (IRMM, Geel, Belgique) pour être approuvée
comme matériel de référence certifié, mais sans succès [57]. Ainsi, le manque de matériel de
référence clairement défini rend l’analyse du gluten une tâche complexe. La PWG-gliadin
reste largement utilisée par la communauté scientifique et sert de standard d’étalonnage dans
certaines trousses ELISA commerciales [58,59]. L’utilisation de gliadine comme standard
d’étalonnage cause des problèmes de sur- et sous-estimation de la quantité totale de gluten
contenue dans l’analyte. Ces difficultés ont été démontrées en fonction de l’origine du gluten
utilisé, soit de blé, d’orge ou de seigle [8]. Plusieurs études récentes cherchent à combler les
lacunes concernant les matériaux de référence pouvant servir dans les trousses ELISA [10–
12,40,60]. La plupart de ces auteurs ont démontré que la variabilité des réponses en fonction
du type de gluten analysé peut être réduite si la calibration de l’ELISA est faite avec le même
type de gluten que l’analyte [10–12,61]. Toutefois, cette calibration nécessite de connaître
l’origine du gluten, qui est fréquemment inconnue en situation d’analyse courante [7,8].
1.4.3-Niveau de transformation de la matrice alimentaire
Le niveau de transformation a un effet direct sur la réponse obtenue par ELISA. En effet, les
traitements de chaleur ont pour effet de créer de forts agrégats entre les prolamines et les
glutélines. Ces changements induisent des différences de solubilité, ce qui rend l’extraction
de ces agrégats difficile et mène à des sous-estimations de la quantité de gluten [55,62].
L’utilisation d’agents dénaturants et désagrégeants est alors nécessaire pour une extraction
efficace du gluten tel que discuté précédemment. Les traitements d’hydrolyse du gluten (ex.
bière) peuvent aussi avoir un impact considérable sur la détection du gluten par les anticorps
[63]. Un ELISA compétitif basé sur le R5 s’est montré efficace pour la détection de
prolamines partiellement hydrolysées, mais cette méthode est seulement compatible avec une
extraction à l’éthanol non dénaturante du gluten, ce qui implique un risque de récupération
12Vous pouvez aussi lire

















































